Inulin and Chinese Gallotannin Affect Meat Quality and Lipid Metabolism on Hu Sheep

 , , , , and
, , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Sample Collection
2.2. Basal Diet
2.3. Measurement of Muscle Fatty Acids
2.4. Extraction and LC-MS Analysis of Urine Metabolites
2.5. Statistical Analysis
3. Results
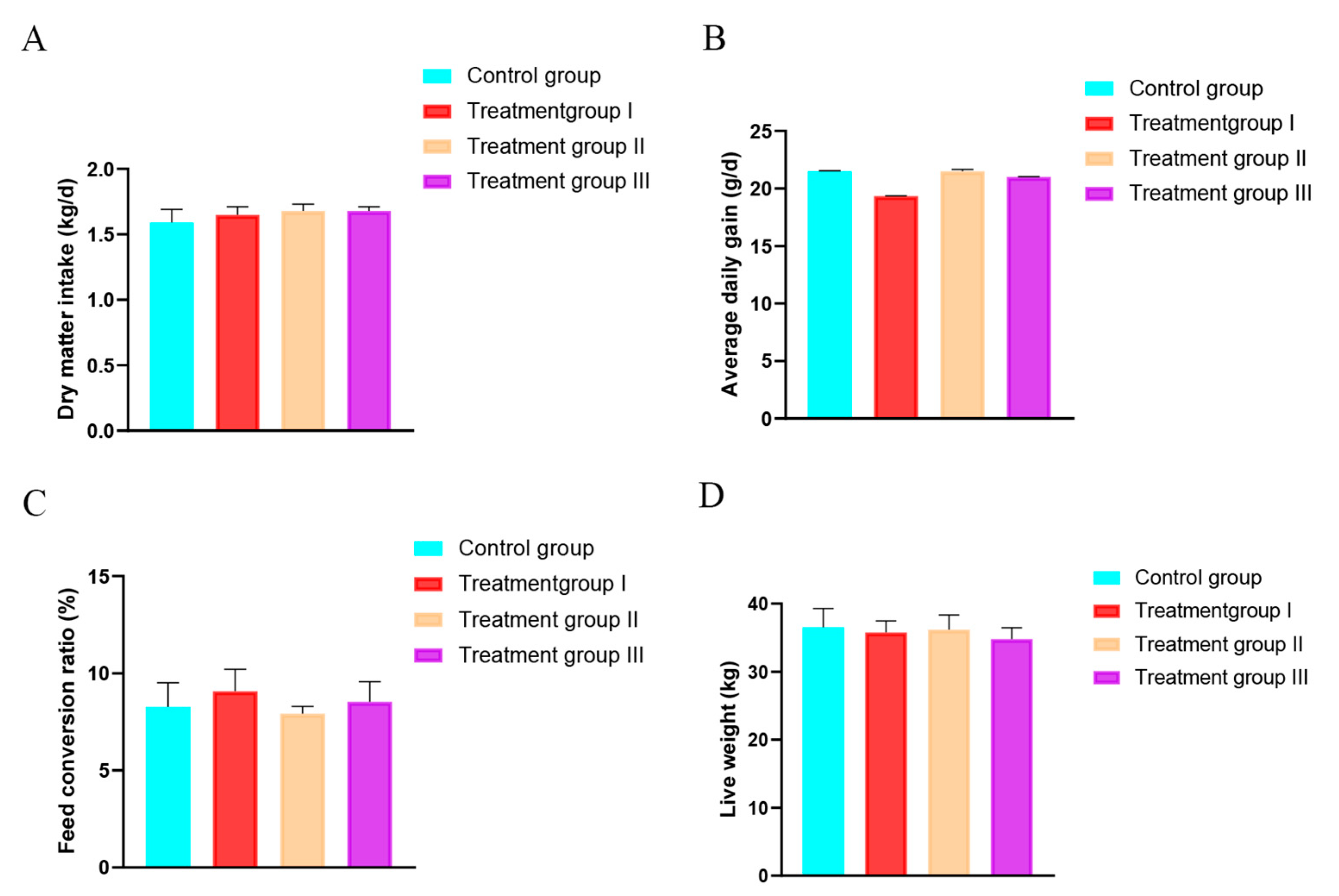
3.1. Effect of Inulin and Chinese Gallotannin on Lamb Performance
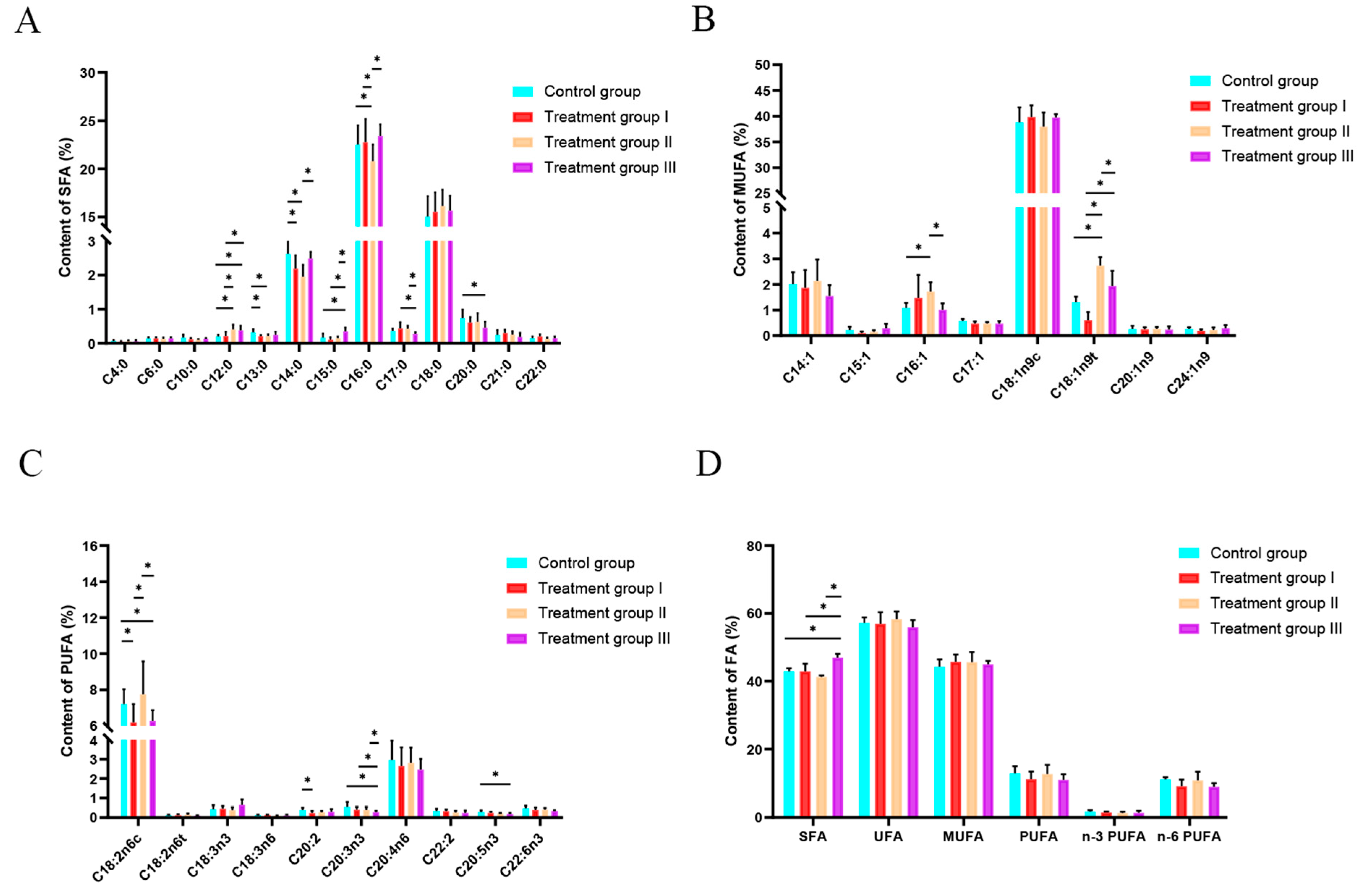
3.2. Effect of Inulin and Chinese Gallotannin on Meat Saturated Fatty Acids
3.3. Effect of Inulin and Chinese Gallotannin on Meat Unsaturated Fatty Acid
3.4. Effect of Inulin and Chinese Gallotannin on Meat Total Fatty Acid
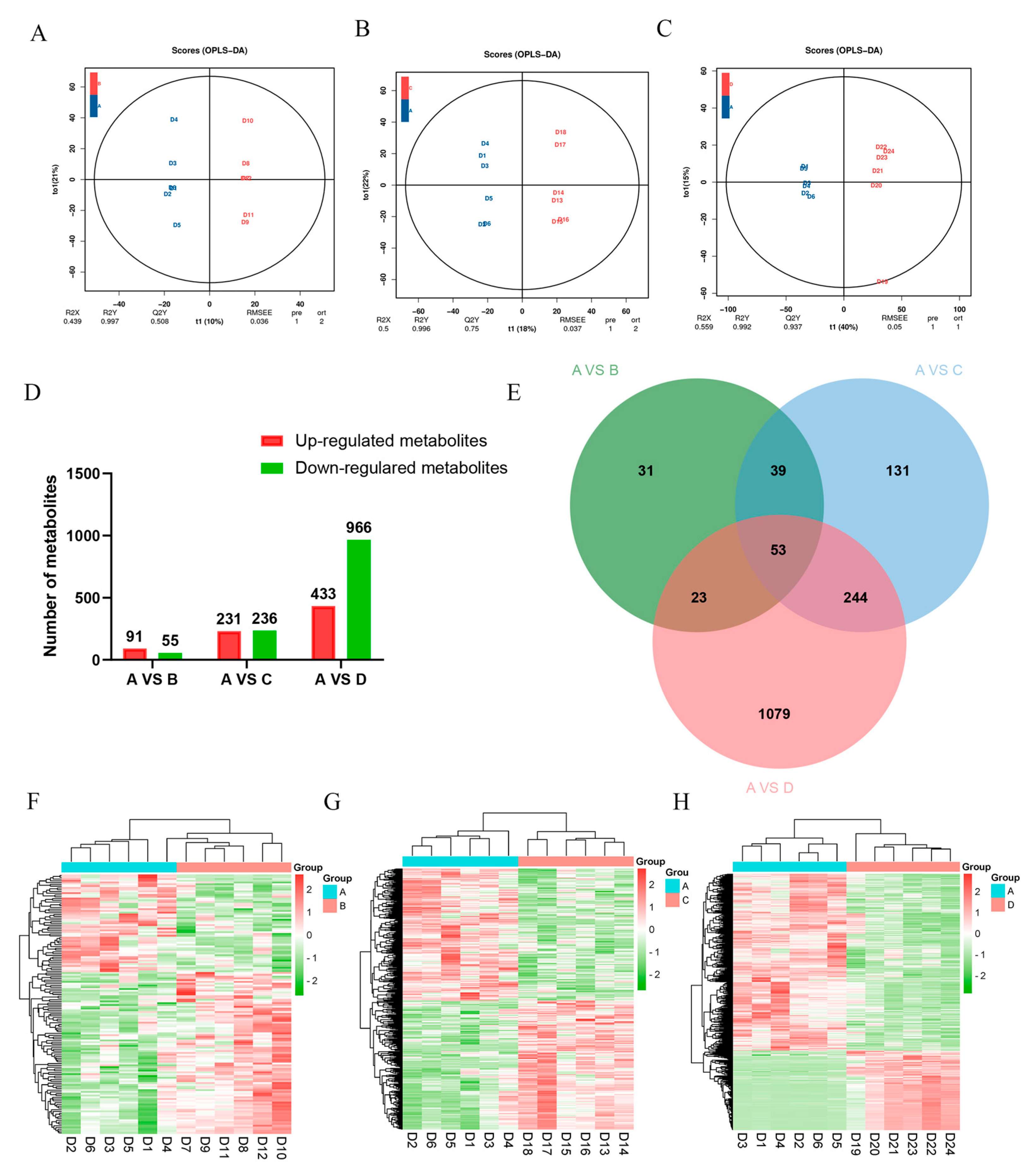
3.5. Screening of Differential Metabolites
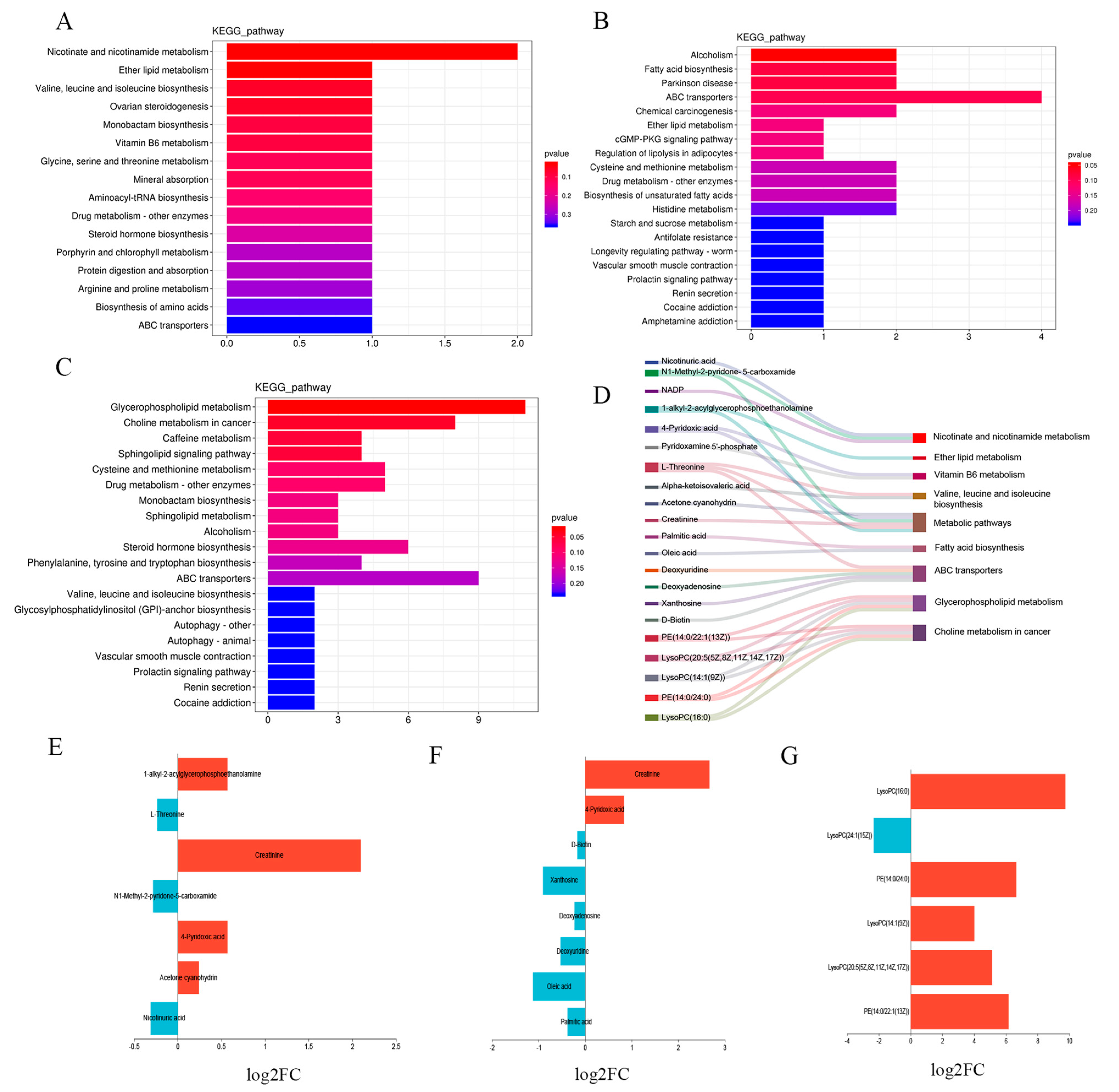
3.6. Functional Enrichment Analysis of Differential Metabolites
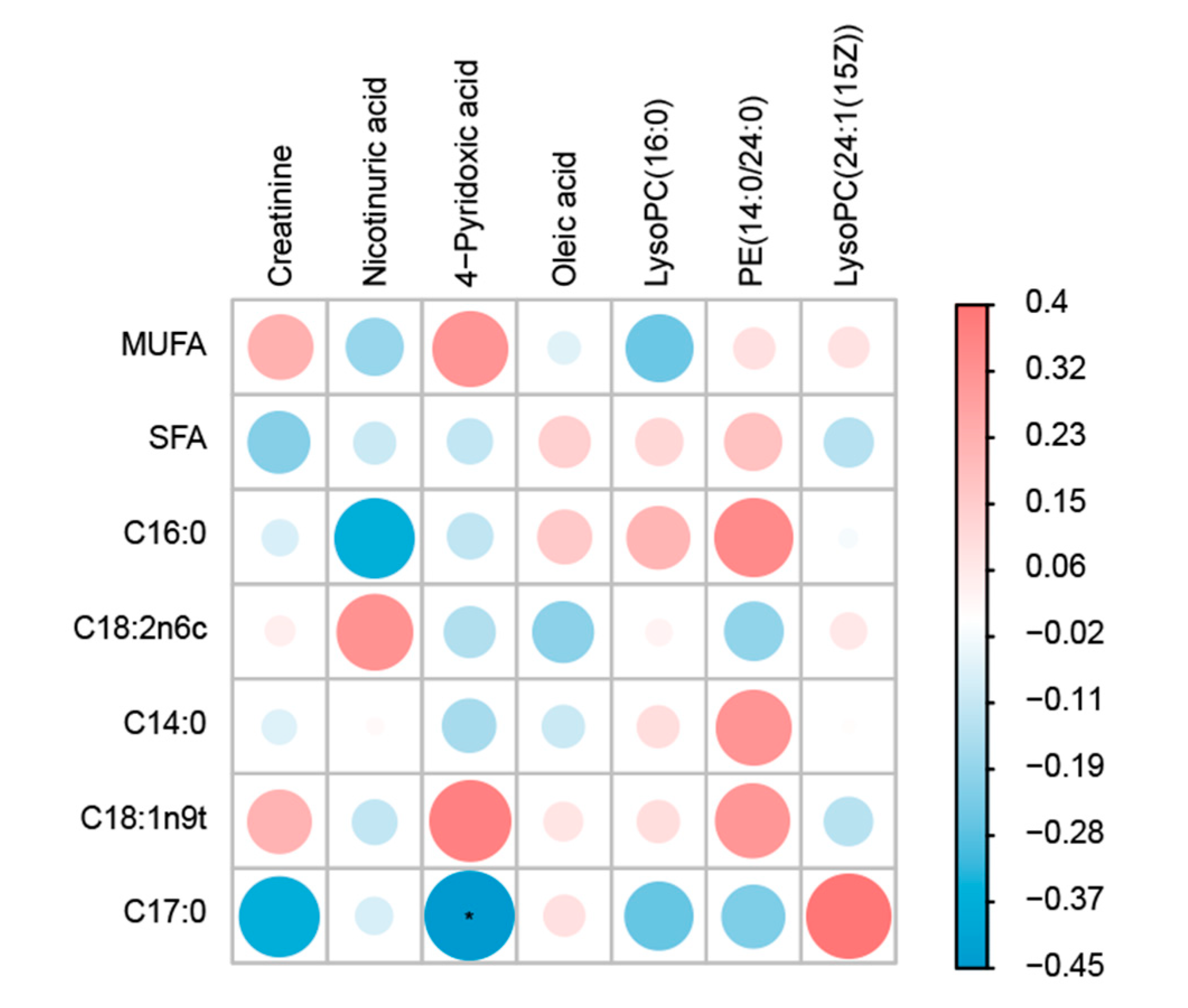
3.7. Correlation Analysis
4. Discussion
4.1. Analysis of Meat Fatty Acid
4.2. Effect of Inulin Addition to Diet on Urine Metabolism in Hu Sheep
4.3. Effect of Inulin and Chinese Gallotannin Addition to Diet on Urine Metabolism in Hu Sheep
4.4. Effect of Inulin, Chinese Gallotannin, and PEG Addition to Diet on Urine Metabolism in Hu Sheep
4.5. Correlation Analysis
4.6. The Clinical Significance of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peng, Q.H.; Cheng, L.; Kun, K.; Gang, T.; Mohammad, A.M.; Bai, X.; Wang, L.Z.; Zou, H.W.; Gicheha, M.G.; Wang, Z.S. Effects of yeast and yeast cell wall polysaccharides supplementation on beef cattle growth performance, rumen microbial populations and lipopolysaccharides production. J. Integr. Agric. 2020, 19, 810–819. [Google Scholar] [CrossRef]
- Van Loo, J.; Coussement, P.; De Leenheer, L.; Hoebregs, H.; Smits, G. On the presence of Inulin and Oligofructose as natural ingredients in the western diet. Crit. Rev. Food Sci. Nutr. 1995, 35, 525–552. [Google Scholar] [CrossRef]
- Shoaib, M.; Shehzad, A.; Omar, M.; Rakha, A.; Raza, H.; Sharif, H.R.; Shakeel, A.; Ansari, A.; Niazi, S. Inulin: Properties, health benefits and food applications. Carbohydr. Polym. 2016, 147, 444–454. [Google Scholar] [CrossRef]
- Verdonk, J.M.; Shim, S.B.; van Leeuwen, P.; Verstegen, M.W. Application of inulin-type fructans in animal feed and pet food. Br. J. Nutr. 2005, 93 (Suppl. S1), S125–S138. [Google Scholar] [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef]
- Uerlings, J.; Arevalo Sureda, E.; Schroyen, M.; Kroeske, K.; Tanghe, S.; De Vos, M.; Bruggeman, G.; Wavreille, J.; Bindelle, J.; Purcaro, G.; et al. Impact of citrus pulp or inulin on intestinal microbiota and metabolites, barrier, and immune function of weaned piglets. Front. Nutr. 2021, 8, 650211. [Google Scholar] [CrossRef]
- Tian, K.; Liu, J.H.; Sun, Y.W.; Wu, Y.J.; Chen, J.C.; Zhang, R.M.; He, T.L.; Dong, G.Z. Effects of dietary supplementation of inulin on rumen fermentation and bacterial microbiota, inflammatory response and growth performance in finishing beef steers fed high or low-concentrate diet. Anim. Feed Sci. Technol. 2019, 258, 114299. [Google Scholar] [CrossRef]
- Cerezuela, R.; Cuesta, A.; Meseguer, J.; Esteban, M.A. Effects of dietary inulin and heat-inactivated Bacillus subtilis on gilthead seabream (Sparus aurata L.) innate immune parameters. Benef. Microbes. 2012, 3, 77–81. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Zhang, W.; Sun, M.; Zhang, Z. Hypoglycemic effect of inulin combined with ganoderma lucidum polysaccharides in T2DM rats. J. Funct. Foods 2019, 55, 381–390. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, J.; Henning, S.M.; Lee, R.; Hsu, M.; Grojean, E.; Pisegna, R.; Ly, A.; Heber, D.; Li, Z. Dietary pomegranate extract and inulin affect gut microbiome differentially in mice fed an obesogenic diet. Anaerobe 2017, 48, 184–193. [Google Scholar] [CrossRef]
- Mueller-Harvey, I. Analysis of hydrolysable tannins. Anim. Feed Sci. Technol. 2001, 91, 3–20. [Google Scholar] [CrossRef]
- Biagia, G.; Cipollini, I.; Paulicks, B.R.; Roth, F.X. Effect of tannins on growth performance and intestinal ecosystem in weaned piglets. Arch. Anim. Nutr. 2010, 64, 121–135. [Google Scholar] [CrossRef]
- Gülçin, İ.; Huyut, Z.; Elmastaş, M.; Aboul-Enein, H.Y. Radical scavenging and antioxidant activity of tannic acid. Arab. J. Chem. 2010, 3, 43–53. [Google Scholar] [CrossRef]
- Ren, Y.Y.; Zhang, X.R.; Li, T.N.; Zeng, Y.J.; Wang, J.; Huang, Q.W. Galla chinensis, a Traditional Chinese Medicine: Comprehensive review of botany, traditional uses, chemical composition, pharmacology and toxicology. J. Ethnopharmacol. 2021, 278, 114247. [Google Scholar] [CrossRef]
- Aboagye, I.A.; Beauchemin, K.A. Potential of molecular weight and structure of tannins to reduce methane emissions from ruminants: A review. Animals 2019, 9, 856. [Google Scholar] [CrossRef]
- Waghorn, G. Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production—Progress and challenges. Anim. Feed Sci. Technol. 2008, 147, 116–139. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D. Preference for polyethylene glycol by sheep fed a quebracho tannin diet. J. Anim. Sci. 2001, 79, 2066–2074. [Google Scholar] [CrossRef][Green Version]
- Silanikove, N.; Perevolotsky, A.; Provenza, F.D. Use of tannin-binding chemicals to assay for tannins and their negative postingestive effects in ruminants. Anim. Feed Sci. Technol. 2001, 91, 69–81. [Google Scholar] [CrossRef]
- Vlaanderen, J.J.; Janssen, N.A.; Hoek, G.; Keski-Rahkonen, P.; Barupal, D.K.; Cassee, F.R.; Gosens, I.; Strak, M.; Steenhof, M.; Lan, Q.; et al. The impact of ambient air pollution on the human blood metabolome. Environ. Res. 2017, 156, 341–348. [Google Scholar] [CrossRef]
- Sawano, S.; Oza, K.; Murakami, T.; Nakamura, M.; Tatsumi, R.; Mizunoya, W. Effect of gender, rearing, and cooking on the metabolomic profile of porcine muscles. Metabolites 2020, 10, 10. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, L.; Johnson, M.; Mandal, R.; Wishart, D.S. Comprehensive targeted metabolomic assay for urine analysis. Anal. Chem. 2020, 92, 10627–10634. [Google Scholar] [CrossRef]
- Karu, N.; Deng, L.; Slae, M.; Guo, A.C.; Sajed, T.; Huynh, H.; Wine, E.; Wishart, D.S. A review on human fecal metabolomics: Methods, applications and the human fecal metabolome database. Anal. Chim. Acta 2018, 1030, 1–24. [Google Scholar] [CrossRef]
- Wang, B.; Ma, M.P.; Diao, Q.Y.; Tu, Y. Saponin-induced shifts in the rumen microbiome and metabolome of young cattle. Front. Microbiol. 2019, 10, 356. [Google Scholar] [CrossRef]
- Wood, J.D.; Richardson, R.I.; Nute, G.R.; Fisher, A.V.; Campo, M.M.; Kasapidou, E.; Sheard, P.R.; Enser, M. Effects of fatty acids on meat quality: A review. Meat Sci. 2004, 66, 21–32. [Google Scholar] [CrossRef]
- Kowalczyk, J.; Skiba, B.; Buczkowski, Z.; Kowalik, B. A device for quantitative urine collection from male sheep in balance trials. J. Anim. Feed Sci. 1996, 5, 297–301. [Google Scholar] [CrossRef]
- Liu, T.; Lei, Z.M.; Wu, J.P.; Brown, M.A. Fatty acid composition differences between adipose depot sites in dairy and beef steer breeds. J. Food Sci. Technol. 2015, 52, 1656–1662. [Google Scholar] [CrossRef][Green Version]
- Garcia, A.; Barbas, C. Gas chromatography-mass spectrometry (GC-MS)-based metabolomics. Metab. Profiling 2011, 708, 191–204. [Google Scholar]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vazquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. LIPID MAPS online tools for lipid research. Nucleic Acids Res. 2007, 35, W606–W612. [Google Scholar] [CrossRef]
- Thevenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the human adult urinary metabolome variations with age, body mass index, and gender by implementing a comprehensive workflow for univariate and OPLS statistical analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Arshad, M.S.; Sohaib, M.; Ahmad, R.S.; Nadeem, M.T.; Imran, A.; Arshad, M.U.; Kwon, J.H.; Amjad, Z. Ruminant meat flavor influenced by different factors with special reference to fatty acids. Lipids Health Dis. 2018, 17, 223. [Google Scholar] [CrossRef]
- Panda, C.; Varadharaj, S.; Voruganti, V.S. PUFA, genotypes and risk for cardiovascular disease. Prostaglandins Leukot. Essent. Fat. Acids 2022, 176, 102377. [Google Scholar] [CrossRef]
- Mukherjee, A.; Kenny, H.A.; Lengyel, E. Unsaturated Fatty Acids Maintain Cancer Cell Stemness. Cell Stem Cell 2017, 20, 291–292. [Google Scholar] [CrossRef]
- Froyen, E. The effects of fat consumption on low-density lipoprotein particle size in healthy individuals: A narrative review. Lipids Health Dis. 2021, 20, 86. [Google Scholar] [CrossRef]
- Flock, M.R.; Kris-Etherton, P.M. Diverse physiological effects of long-chain saturated fatty acids: Implications for cardiovascular disease. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 133–140. [Google Scholar] [CrossRef]
- Sokoła-Wysoczańska, E.; Wysoczański, T.; Wagner, J.; Czyż, K.; Bodkowski, R.; Lochyński, S.; Patkowska-Sokoła, B. Polyunsaturated fatty acids and their potential therapeutic role in cardiovascular system disorders—A review. Nutrients 2018, 10, 1561. [Google Scholar] [CrossRef]
- Hosomi, R.; Fukunaga, K.; Arai, H.; Kanda, S.; Nishiyama, T.; Kanada, T.; Yoshida, M. Effect of phospholipid n-3 polyunsaturated fatty acids on rat lipid metabolism. Eur. J. Lipid Sci. Technol. 2010, 112, 537–544. [Google Scholar] [CrossRef]
- Baublits, R.T.; Pohlman, F.W.; Brown, A.H.; Johnson, Z.B.; Proctor, A.; Sawyer, J.; Dias-Morse, P.; Galloway, D.L. Injection of conjugated linoleic acid into beef strip loins. Meat Sci. 2007, 75, 84–93. [Google Scholar] [CrossRef]
- Hernández-Sánchez, D.; Gómez-Hernández, J.L.; Crosby-Galván, M.M.; Hernández-Anguiano, A.M.; Ramirez-Bribiesca, J.E.; Aranda-Ibañez, E.; Gonzalez-Muñoz, S.S.; Pinto-Ruiz, R. 1632 Inulin as prebiotic for Lactobacillus salivarius and Enterococcus faecium with probiotic potential in ruminants. J. Anim. Sci. 2016, 94, 794–795. [Google Scholar] [CrossRef]
- Tripodo, G.; Mandracchia, D. Inulin as a multifaceted (active) substance and its chemical functionalization: From plant extraction to applications in pharmacy, cosmetics and food. Eur. J. Pharm. Biopharm. 2019, 141, 21–36. [Google Scholar] [CrossRef]
- Zou, Y.F.; Li, C.Y.; Fu, Y.P.; Feng, X.; Peng, X.; Feng, B.; Li, L.X.; Jia, R.Y.; Huang, C.; Song, X.; et al. Restorative Effects of Inulin from Codonopsis pilosula on Intestinal Mucosal Immunity, Anti-Inflammatory Activity and Gut Microbiota of Immunosuppressed Mice. Front. Pharmacol. 2022, 13, 786141. [Google Scholar] [CrossRef]
- Huang, C.F.; Cheng, M.L.; Fan, C.M.; Hong, C.Y.; Shiao, M.S. Nicotinuric Acid: A potential marker of metabolic syndrome through a metabolomics-based approach. Diabetes Care 2013, 36, 1729–1731. [Google Scholar] [CrossRef]
- Zuo, J.J.; Lan, Y.; Hu, H.L.; Hou, X.Q.; Li, J.S.; Wang, T.; Zhang, H.; Zhang, N.N.; Guo, C.N.; Peng, F.; et al. Metabolomics-based multidimensional network biomarkers for diabetic retinopathy identification in patients with type 2 diabetes mellitus. BMJ Open Diabetes Res. Care 2021, 9, e001443. [Google Scholar] [CrossRef]
- Jones, K.M. The mechanism of nicotinuric acid synthesis. Biochem. J. 1959, 73, 714–719. [Google Scholar] [CrossRef]
- Rinaldo, P.; Schmidt-Sommerfeld, E.; Posca, A.P.; Heales, S.J.R.; Woolf, D.A.; Leonard, J.V. Effect of treatment with glycine and L-carnitine in medium-chain acyl-coenzyme A dehydrogenase deficiency. J. Pediatr. 1993, 122, 580–584. [Google Scholar] [CrossRef]
- Ellis, B.A.; Poynten, A.N.N.; Lowy, A.J.; Furler, S.M.; Cooney, G.J. Long-chain acyl-CoA esters as indicators of lipid metabolism and insulin sensitivity in rat and human muscle. J. Appl. Physiol. Endocrinol. Metab. 2000, 279, E554–E560. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Rochell, S.J.; Applegate, T.J. Threonine, arginine, and glutamine: Influences on intestinal physiology, immunology, and microbiology in broilers. Poult. Sci. 2018, 97, 937–945. [Google Scholar] [CrossRef]
- Feng, J.; Li, X.; Pei, F.; Chen, X.; Li, S.; Nie, Y. 1H NMR analysis for metabolites in serum and urine from rats administrated chronically with La(NO3)3. Anal. Biochem. 2002, 301, 1–7. [Google Scholar] [CrossRef]
- Runesson, B.; Qureshi, A.R.; Xu, H.; Gasparini, A.; Lindholm, B.; Barany, P.; Elinder, C.G.; Carrero, J.J. Causes of death across categories of estimated glomerular filtration rate: The Stockholm CREAtinine Measurements (SCREAM) project. PLoS ONE 2019, 14, e0209440. [Google Scholar] [CrossRef]
- Toney, M.D. Reaction specificity in pyridoxal phosphate enzymes. Arch. Biochem. Biophys. 2005, 433, 279–287. [Google Scholar] [CrossRef]
- Tanaka, T.; Tateno, Y.; Gojobori, T. Evolution of vitamin B6 (pyridoxine) metabolism by gain and loss of genes. Mol. Biol. Evol. 2005, 22, 243–250. [Google Scholar] [CrossRef]
- Piccinin, E.; Cariello, M.; De Santis, S.; Ducheix, S.; Sabbà, C.; Ntambi, J.M.; Moschetta, A. Role of oleic acid in the gut-liver axis: From diet to the regulation of its synthesis via stearoyl-CoA desaturase 1 (SCD1). Nutrients 2019, 11, 2283. [Google Scholar] [CrossRef]
- Tsai, Y.W.; Lu, C.H.; Chang, R.C.; Hsu, Y.P.; Ho, L.T.; Shih, K.C. Palmitoleic acid ameliorates palmitic acid-induced proinflammation in J774A.1 macrophages via TLR4-dependent and TNF-alpha-independent signallings. Prostaglandins Leukot. Essent. Fat. Acids 2021, 169, 102270. [Google Scholar] [CrossRef]
- Hierons, S.J.; Abbas, K.; Sobczak, A.I.S.; Cerone, M.; Smith, T.K.; Ajjan, R.A.; Stewart, A.J. Changes in plasma free fatty acids in obese patients before and after bariatric surgery highlight alterations in lipid metabolism. Sci. Rep. 2022, 12, 15337. [Google Scholar] [CrossRef]
- Bogdanov, M.; Umeda, M.; Dowhan, W. Phospholipid-assisted refolding of an integral membrane protein. Minimum structural features for phosphatidylethanolamine to act as a molecular chaperone. J. Biol. Chem. 1999, 274, 12339–12345. [Google Scholar] [CrossRef]
- Zhou, X.S.; Wang, R.P.; Zhang, T.; Liu, F.; Zhang, W.; Wang, G.L.; Gu, G.R.; Han, Q.Q.; Xu, D.M.; Yao, C.L.; et al. Identification of lysophosphatidylcholines and sphingolipids as potential biomarkers for acute aortic dissection via serum metabolomics. Eur. J. Vasc. Endovasc. Surg. 2018, 57, 434–441. [Google Scholar] [CrossRef]
- Chang, M.C.; Lee, J.J.; Chen, Y.J.; Lin, S.I.; Lin, L.D.; Liou, E.J.W.; Huang, W.L.; Chan, C.P.; Huang, C.C.; Jeng, J.H. Lysophosphatidylcholine induces cytotoxicity/apoptosis and IL-8 production of human endothelial cells: Related mechanisms. Oncotarget 2017, 8, 106177–106189. [Google Scholar] [CrossRef]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef]
- Hung, N.D.; Sok, D.E.; Kim, M.R. Prevention of 1-palmitoyl lysophosphatidylcholine-induced inflammation by polyunsaturated acyl lysophosphatidylcholine. Inflamm. Res. 2012, 61, 473–483. [Google Scholar] [CrossRef]
- Parra, M.; Stahl, S.; Hellmann, H. Vitamin B(6) and Its Role in Cell Metabolism and Physiology. Cells 2018, 7, 84. [Google Scholar] [CrossRef]
- Niness, K.R. Inulin and Oligofructose: What Are They? J. Nutr. 1999, 129, 1402S–1406S. [Google Scholar] [CrossRef]
- Quinville, B.M.; Deschenes, N.M.; Ryckman, A.E.; Walia, J.S. A comprehensive review: Sphingolipid metabolism and implications of disruption in sphingolipid homeostasis. Int. J. Mol. Sci. 2021, 22, 5793. [Google Scholar] [CrossRef]
- Kalinichenko, L.S.; Gulbins, E.; Kornhuber, J.; Muller, C.P. Sphingolipid control of cognitive functions in health and disease. Prog. Lipid. Res. 2022, 86, 101162. [Google Scholar] [CrossRef]
- Reiner, A.P.; Gross, M.D.; Carlson, C.S.; Bielinski, S.J.; Lange, L.A.; Fornage, M.; Jenny, N.S.; Walston, J.; Tracy, R.P.; Williams, O.D.; et al. Common coding variants of the HNF1A gene are associated with multiple cardiovascular risk phenotypes in community-based samples of younger and older European-American adults: The Coronary Artery Risk Development in Young Adults Study and The Cardiovascular Health Study. Circ. Cardiovasc. Genet. 2009, 2, 244–254. [Google Scholar]
- Wang, Y.; Ai, Z.; Xing, X.; Fan, Y.; Zhang, Y.; Nan, B.; Li, X.; Wang, Y.; Liu, J. The ameliorative effect of probiotics on diet-induced lipid metabolism disorders: A review. Crit. Rev. Food Sci. Nutr. 2022, 1–17. [Google Scholar] [CrossRef]
- Innis, S.M. Fatty acids and early human development. Early Hum. Dev. 2007, 83, 761–766. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Material | Proportion | Nutrient Levels | Content |
|---|---|---|---|
| Ensilage corn stalks (%) | 8 | DM (kg/d) | 1 |
| Alfalfa hay (%) | 22 | ME (MJ/kg) | 12.01 |
| Corn (%) | 34 | CP (%) | 18.17 |
| Soybean meal (%) | 14 | Ca (%) | 0.31 |
| Rice bran (%) | 3 | P (%) | 0.30 |
| Flax cake (%) | 3 | NDF (%) | 23.22 |
| Corn DDGS (%) | 12 | ||
| NaCl (%) | 0.7 | ||
| Premix 1 (%) | 0.5 | ||
| Puffing urea (%) | 1.3 | ||
| NaHCO3 (%) | 1.5 | ||
| Total (%) | 100 |
| Item | Treatment | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| A 1 | B 2 | C 3 | D 4 | |||
| C4:0 (%) | 0.08 ± 0.02 | 0.05 ± 0.02 | 0.06 ± 0.03 | 0.06 ± 0.04 | 0.01 | 0.271 |
| C6:0 (%) | 0.15 ± 0.03 | 0.15 ± 0.03 | 0.13 ± 0.04 | 0.15 ± 0.03 | 0.01 | 0.630 |
| C10:0 (%) | 0.17 ± 0.09 | 0.12 ± 0.04 | 0.11 ± 0.02 | 0.14 ± 0.02 | 0.01 | 0.277 |
| C12:0 (%) | 0.20 b ± 0.05 | 0.22 b ± 0.12 | 0.41 a ± 0.14 | 0.39 a ± 0.13 | 0.03 | 0.006 |
| C13:0 (%) | 0.33 a ± 0.09 | 0.21 b ± 0.03 | 0.22 b ± 0.05 | 0.25 ab ± 0.09 | 0.02 | 0.036 |
| C14:0 (%) | 2.62 a ± 0.88 | 2.19 bc ± 0.39 | 1.95 c ± 0.35 | 2.49 ab ± 0.19 | 0.11 | 0.001 |
| C15:0 (%) | 0.17 b ± 0.12 | 0.11 b ± 0.07 | 0.16 b ± 0.05 | 0.35 a ± 0.11 | 0.03 | 0.001 |
| C16:0 (%) | 22.53 a ± 1.98 | 22.77 a ± 2.40 | 20.79 b ±1.73 | 23.41 a ± 1.20 | 0.41 | 0.002 |
| C17:0 (%) | 0.37 ab ± 0.07 | 0.44 a ± 0.18 | 0.43 a ± 0.09 | 0.28 b ± 0.05 | 0.03 | 0.042 |
| C18:0 (%) | 15.03 ± 2.13 | 15.52 ± 2.02 | 16.13 ± 1.70 | 15.65 ± 1.55 | 0.36 | 0.788 |
| C20:0 (%) | 0.74 a ± 0.25 | 0.62 ab ± 0.15 | 0.63 ab ± 0.26 | 0.46 b ± 0.17 | 0.05 | 0.019 |
| C21:0 (%) | 0.25 ± 0.14 | 0.31 ± 0.09 | 0.26 ± 0.10 | 0.19 ± 0.12 | 0.03 | 0.418 |
| C22:0 (%) | 0.16 ± 0.05 | 0.20 ± 0.07 | 0.13 ± 0.04 | 0.16 ± 0.05 | 0.01 | 0.272 |
| Item | Treatment | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| A 1 | B 2 | C 3 | D 4 | |||
| MUFA | ||||||
| C14:1 (%) | 2.01 ± 0.46 | 1.87 ± 0.69 | 2.14 ± 0.83 | 1.55 ± 0.42 | 0.12 | 0.409 |
| C15:1 (%) | 0.23 ± 0.12 | 0.11 ± 0.06 | 0.15 ± 0.06 | 0.29 ± 0.18 | 0.03 | 0.073 |
| C16:1 (%) | 1.08 b ± 0.20 | 1.47 ab ± 0.90 | 1.72 a ± 0.37 | 1.01 b ± 0.25 | 0.12 | 0.045 |
| C17:1 (%) | 0.57 ± 0.09 | 0.47 ± 0.09 | 0.47 ± 0.06 | 0.47 ± 0.10 | 0.02 | 0.151 |
| C18:1n9c (%) | 38.83 ± 2.90 | 39.86 ± 2.26 | 37.96 ± 2.76 | 39.75 ± 0.64 | 0.47 | 0.465 |
| C18:1n9t (%) | 1.31 bc ± 0.21 | 0.61 c ± 0.31 | 2.73 a ± 0.33 | 1.94 b ± 0.59 | 0.22 | 0.001 |
| C20:1n9 (%) | 0.26 ± 0.13 | 0.25 ± 0.08 | 0.26 ± 0.08 | 0.24 ± 0.13 | 0.02 | 0.987 |
| C24:1n9 (%) | 0.26 ± 0.07 | 0.19 ± 0.06 | 0.23 ± 0.09 | 0.29 ± 0.12 | 0.02 | 0.341 |
| PUFA | ||||||
| C18:2n6c (%) | 7.21 a ± 0.82 | 6.20 b ± 1.00 | 7.75 a ± 1.82 | 6.27 b ± 0.60 | 0.26 | 0.034 |
| C18:2n6t (%) | 0.11 ± 0.04 | 0.12 ± 0.05 | 0.14 ± 0.07 | 0.09 ± 0.03 | 0.01 | 0.431 |
| C18:3n3 (%) | 0.43 ± 0.20 | 0.45 ± 0.14 | 0.38 ± 0.15 | 0.65 ± 0.27 | 0.05 | 0.258 |
| C18:3n6 (%) | 0.12 ± 0.03 | 0.13 ± 0.02 | 0.09 ± 0.02 | 0.10 ± 0.05 | 0.01 | 0.401 |
| C20:2 (%) | 0.39 a ± 0.11 | 0.23 b ± 0.11 | 0.27 ab ± 0.07 | 0.30 ab ± 0.14 | 0.02 | 0.028 |
| C20:3n3 (%) | 0.55 a ± 0.24 | 0.40 a ± 0.14 | 0.39 a ± 0.15 | 0.28 b ± 0.07 | 0.04 | 0.026 |
| C20:4n6 (%) | 2.97 ± 1.00 | 2.65 ± 0.97 | 2.81 ± 0.80 | 2.47 ± 0.55 | 0.17 | 0.771 |
| C22:2 (%) | 0.34 ± 0.11 | 0.33 ± 0.08 | 0.26 ± 0.09 | 0.24 ± 0.12 | 0.02 | 0.279 |
| C20:5n3 (%) | 0.29 a ± 0.08 | 0.24 ab ± 0.06 | 0.21 ab ± 0.03 | 0.19 b ± 0.05 | 0.02 | 0.038 |
| C22:6n3 (%) | 0.48 ± 0.13 | 0.39 ± 0.13 | 0.40 ± 0.09 | 0.34 ± 0.04 | 0.02 | 0.280 |
| Item | Treatment | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| A 1 | B 2 | C 3 | D 4 | |||
| SFA (%) | 42.92 b ± 0.90 | 42.91 b ± 2.27 | 41.27 b ± 0.44 | 46.91 a ± 1.15 | 0.62 | 0.001 |
| UFA (%) | 57.16 ± 1.62 | 56.88 ± 3.45 | 58.29 ± 2.20 | 55.93 ± 2.07 | 0.49 | 0.431 |
| MUFA (%) | 44.27 ± 2.14 | 45.74 ± 2.10 | 45.64 ± 2.98 | 44.98 ± 1.04 | 0.43 | 0.268 |
| PUFA (%) | 12.89 ± 2.11 | 11.14 ± 2.28 | 12.66 ± 2.71 | 10.96 ± 1.67 | 0.46 | 0.327 |
| n-3PUFA (%) | 1.76 ± 0.38 | 1.48 ± 0.26 | 1.32 ± 0.36 | 1.36 ± 0.59 | 0.09 | 0.239 |
| n-6PUFA (%) | 11.10 ± 0.67 | 9.10 ± 1.95 | 10.80 ± 2.55 | 8.92 ± 1.08 | 0.40 | 0.104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Cheng, L.; Li, S.; Liu, Q.; Liang, X.; Hu, J.; Wang, J.; Liu, X.; Zhao, F. Inulin and Chinese Gallotannin Affect Meat Quality and Lipid Metabolism on Hu Sheep. Animals 2023, 13, 160. https://doi.org/10.3390/ani13010160
He Z, Cheng L, Li S, Liu Q, Liang X, Hu J, Wang J, Liu X, Zhao F. Inulin and Chinese Gallotannin Affect Meat Quality and Lipid Metabolism on Hu Sheep. Animals. 2023; 13(1):160. https://doi.org/10.3390/ani13010160
Chicago/Turabian StyleHe, Zhaohua, Long Cheng, Shaobin Li, Qiaoling Liu, Xue Liang, Jiang Hu, Jiqing Wang, Xiu Liu, and Fangfang Zhao. 2023. "Inulin and Chinese Gallotannin Affect Meat Quality and Lipid Metabolism on Hu Sheep" Animals 13, no. 1: 160. https://doi.org/10.3390/ani13010160
APA StyleHe, Z., Cheng, L., Li, S., Liu, Q., Liang, X., Hu, J., Wang, J., Liu, X., & Zhao, F. (2023). Inulin and Chinese Gallotannin Affect Meat Quality and Lipid Metabolism on Hu Sheep. Animals, 13(1), 160. https://doi.org/10.3390/ani13010160