
Distribution and Difference of Gastrointestinal Flora in Sheep with Different Body Mass Index


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Groups
2.2. Sample Collection and Character Determination
2.3. DNA Extraction and Amplification
2.4. Library Construction and Data Processing
3. Results
3.1. Baseline Characteristics of Test Population
3.2. Sequencing Data Overview
3.3. Diversity Analysis
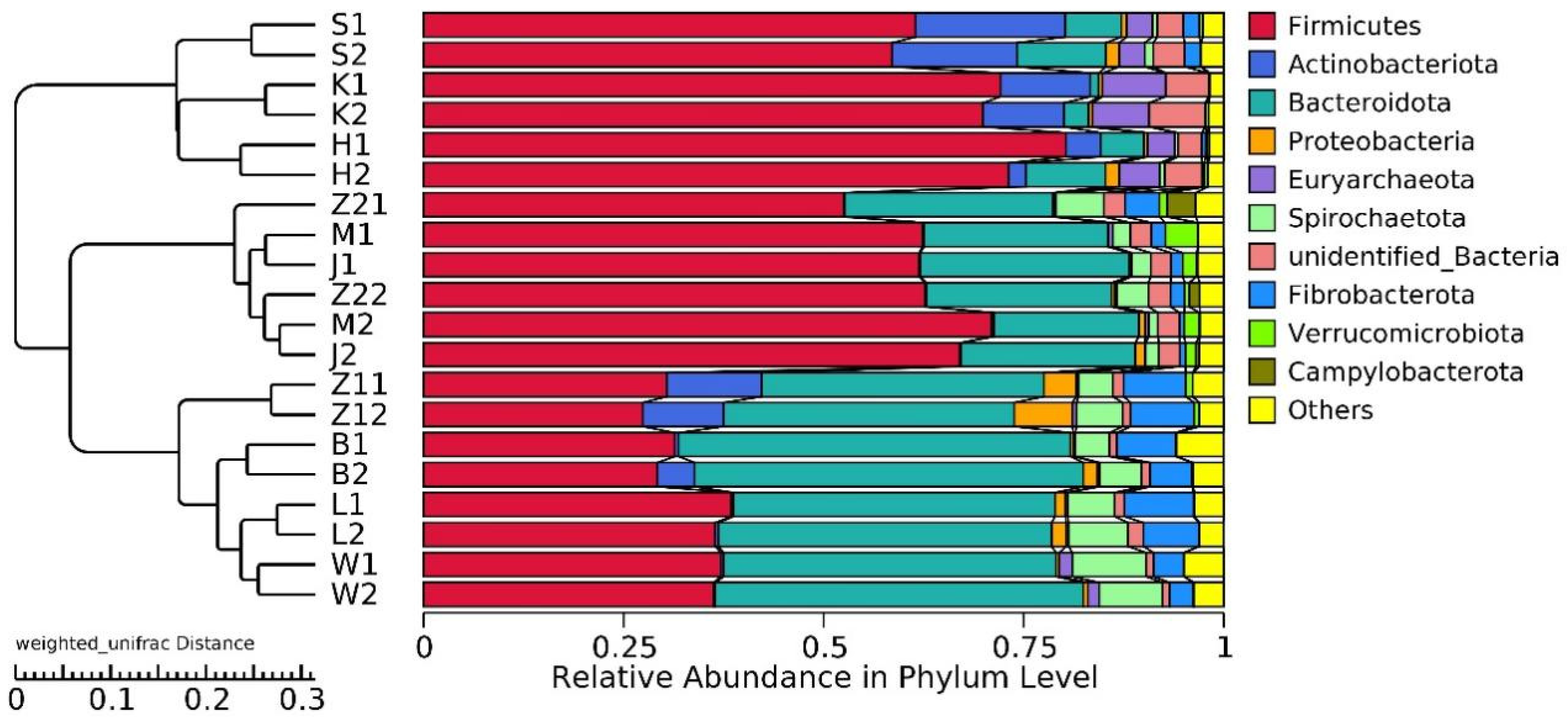
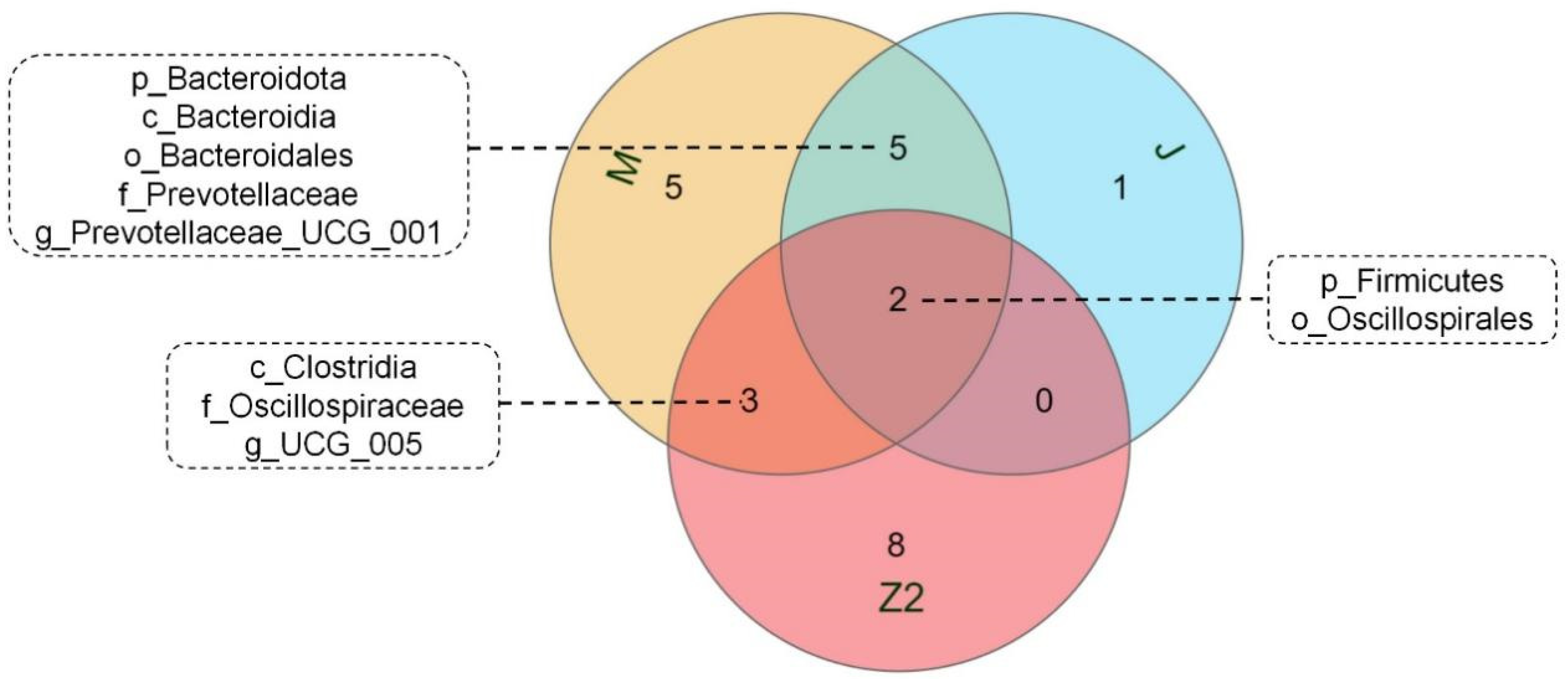
3.4. Analysis of the Gastrointestinal Tract Microbiota Composition
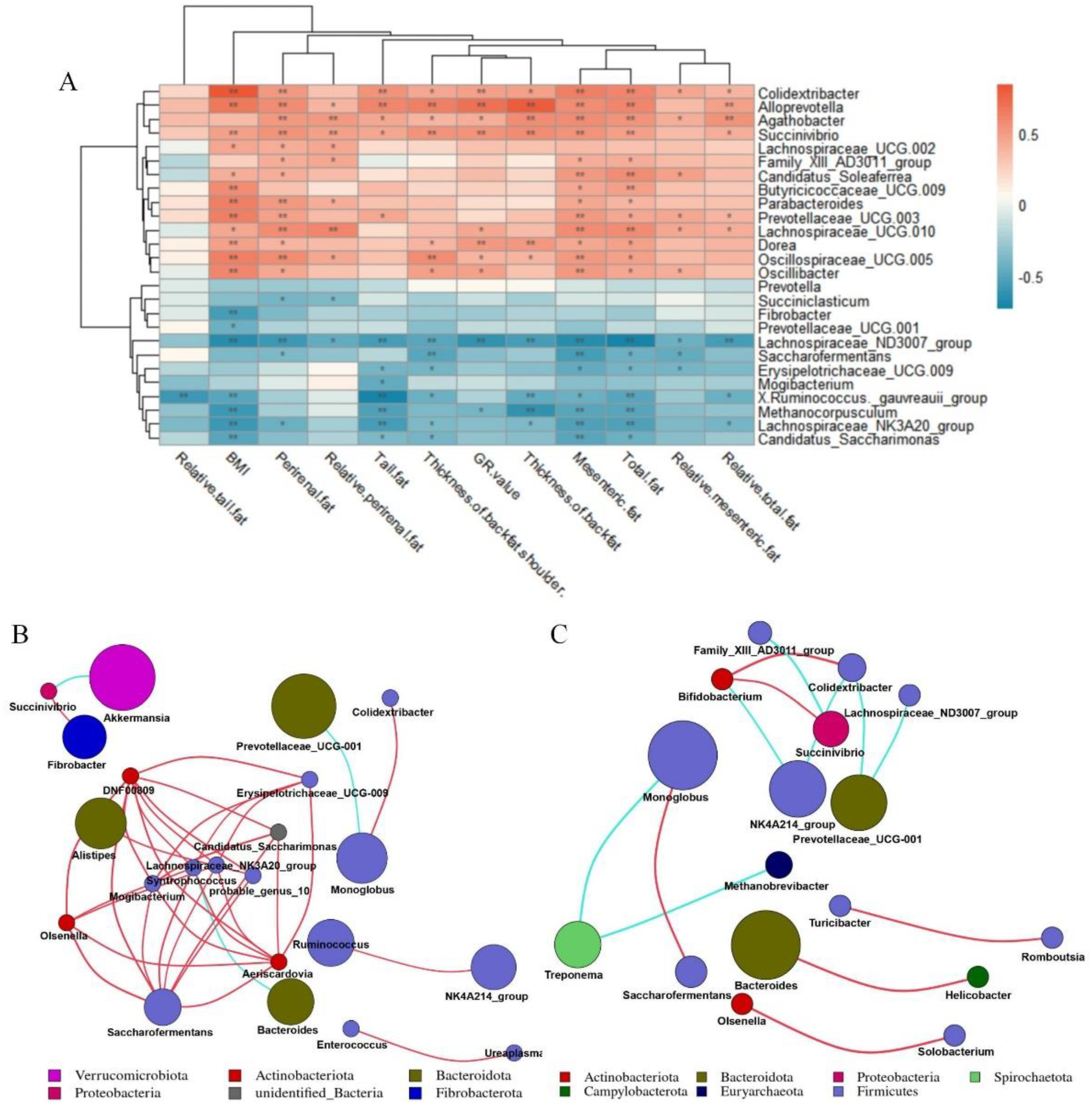
3.5. Cecal Microorganisms Affect Fat Deposition
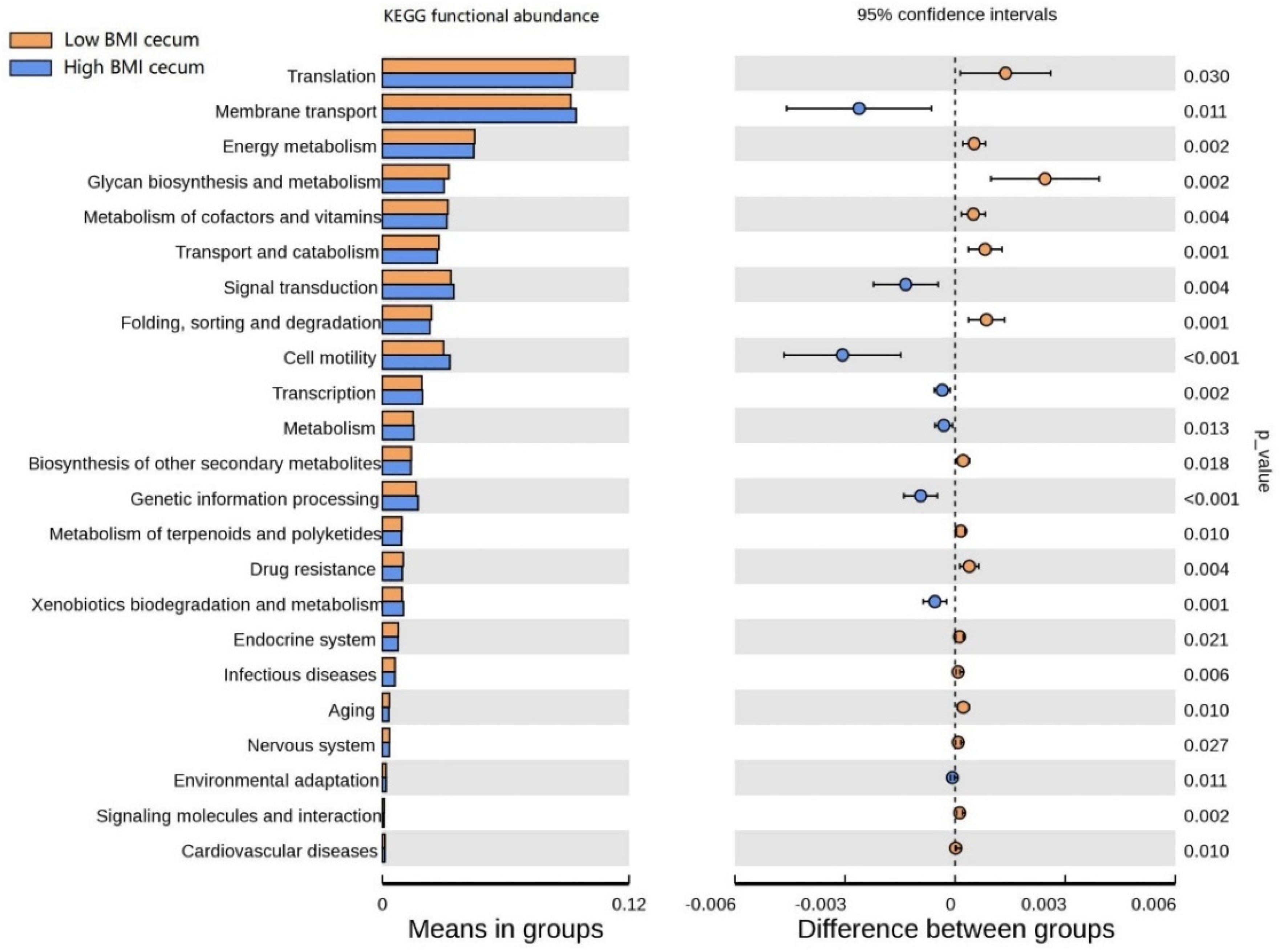
3.6. Functional Prediction Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberto, F.J.; Boyer, F.; Orozco-terWengel, P.; Streeter, I.; Servin, B.; de Villemereuil, P.; Benjelloun, B.; Librado, P.; Biscarini, F.; Colli, L.; et al. Convergent genomic signatures of domestication in sheep and goats. Nat. Commun. 2018, 9, 813. [Google Scholar] [CrossRef] [PubMed]
- Montossi, F.; Font-i-Furnols, M.; del Campo, M.; San Julián, R.; Brito, G.; Sañudo, C. Sustainable sheep production and consumer preference trends: Compatibilities, contradictions, and unresolved dilemmas. Meat Sci. 2013, 95, 772–789. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Jin, Y.; Cui, H.; Yu, L.; Luo, Y.; Wang, S.; Wang, H. Effects of diet supplementation with rumen-protected betaine on carcass characteristics and fat deposition in growing lambs. Meat Sci. 2020, 166, 108154. [Google Scholar] [CrossRef] [PubMed]
- Chavarría-Aguilar, L.; García-Herrera, R.; Salazar-Cuytun, R.; Chay-Canul, A.J.; Casanova-Lugo, F.; PiñEiro-Vázquez, A.; Aguilar-Caballero, A.J. Relationship between body fat depots and body mass index in Pelibuey ewes. Small Rumin. Res. 2016, 141, 124–126. [Google Scholar] [CrossRef]
- Zierle-Ghosh, A.; Jan, A. Physiology, Body Mass Index. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Yang, Y.; Miao, Q.; Zhu, X.; Qin, L.; Gong, W.; Zhang, S.; Zhang, Q.; Lu, B.; Ye, H.; Li, Y. Sleeping Time, BMI, and Body Fat in Chinese Freshmen and Their Interrelation. Obes. Facts 2020, 13, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, M.; Kerr, D.; Uchida, H.; Binns, C.W. Differences in the relationship between BMI and percentage body fat between Japanese and Australian-Caucasian young men. Br. J. Nutr. 2006, 95, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, M.O. Genetics of obesity: What genetic association studies have taught us about the biology of obesity and its complications. Lancet Diabetes Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef]
- Cornejo-Pareja, I.; Muñoz-Garach, A.; Clemente-Postigo, M.; Tinahones, F.J. Importance of gut microbiota in obesity. Eur. J. Clin. Nutr. 2019, 72, 26–37. [Google Scholar] [CrossRef]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial dysbiosis-induced obesity: Role of gut microbiota in homoeostasis of energy metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef]
- Jian, S.; Obin, M.S.; Zhao, L. The Gut Microbiota, Obesity and Insulin Resistance. Mol. Asp. Med. 2012, 34, 39–58. [Google Scholar]
- Yang, G.; Zhang, S.; Li, Z.; Huang, J.; Liu, Y.; Liu, Y.; Wang, Q.; Li, X.; Yan, Y.; Li, M. Comparison Between the Gut Microbiota in Different Gastrointestinal Segments of Large-Tailed Han and Small-Tailed Han Sheep Breeds with High-Throughput Sequencing. Indian J. Microbiol. 2020, 60, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.J.; Wang, M.C. Microbial metabolites regulate host lipid metabolism through NR5A-Hedgehog signalling. Nat. Cell Biol. 2017, 19, 550–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi-Abyane, M.; Alipour, D.; Moghimi, H.R. Effects of different sources of nitrogen on performance, relative population of rumen microorganisms, ruminal fermentation and blood parameters in male feedlotting lambs. Anim. Int. J. Anim. Biosci. 2020, 14, 1438–1446. [Google Scholar] [CrossRef]
- Wang, J.; Fan, H.; Han, Y.; Zhao, J.; Zhou, Z. Characterization of the microbial communities along the gastrointestinal tract of sheep by 454 pyrosequencing analysis. Asian-Australas. J. Anim. Sci. 2017, 30, 100–110. [Google Scholar] [CrossRef]
- Xie, F.; Jin, W.; Si, H.; Yuan, Y.; Tao, Y.; Liu, J.; Wang, X.; Yang, C.; Li, Q.; Yan, X.; et al. An integrated gene catalog and over 10,000 metagenome-assembled genomes from the gastrointestinal microbiome of ruminants. Microbiome 2021, 9, 137. [Google Scholar] [CrossRef]
- Ekiz, B.; Kecici, P.D.; Ograk, Y.Z.; Yalcintan, H.; Yilmaz, A. Evaluation of the functionality of EUROP carcass classification system in thin-tailed and fat-tailed lambs. Meat Sci. 2021, 181, 108603. [Google Scholar] [CrossRef]
- Gardner, G.E.; Anderson, F.; Smith, C.; Williams, A. Using dual energy X-ray absorptiometry to estimate commercial cut weights at abattoir chain-speed. Meat Sci. 2021, 173, 108400. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef] [PubMed]
- Freedman, D.S.; Wang, J.; Maynard, L.M.; Thornton, J.C.; Mei, Z.; Pierson, R.N.; Dietz, W.H.; Horlick, M. Relation of BMI to fat and fat-free mass among children and adolescents. Int. J. Obes. 2005, 29, 1–8. [Google Scholar] [CrossRef]
- Wang, J.; Gallagher, D.; Thornton, J.C.; Yu, W.; Weil, R.; Kovac, B.; Pi-Sunyer, F.X. Regional body volumes, BMI, waist circumference, and percentage fat in severely obese adults. Obesity 2007, 15, 2688–2698. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Sha, Y.; Dingkao, R.; Zhang, W.; Lv, W.; Wei, H.; Shi, H.; Hu, J.; Wang, J.; Li, S.; et al. Interactions Between Rumen Microbes, VFAs, and Host Genes Regulate Nutrient Absorption and Epithelial Barrier Function During Cold Season Nutritional Stress in Tibetan Sheep. Front. Microbiol. 2020, 11, 593062. [Google Scholar] [CrossRef]
- Ping, Z.; Pei, X.; Xia, P.; Chen, Y.; Guo, R.; Hu, C.; Imam, M.U.; Chen, Y.; Sun, P.; Liu, L. Anthropometric indices as surrogates for estimating abdominal visceral and subcutaneous adipose tissue: A meta-analysis with 16,129 participants. Diabetes Res. Clin. Pract. 2018, 143, 310–319. [Google Scholar] [CrossRef]
- Flegal, K.M.; Shepherd, J.A.; Looker, A.C.; Graubard, B.I.; Borrud, L.G.; Ogden, C.L.; Harris, T.B.; Everhart, J.E.; Schenker, N. Comparisons of percentage body fat, body mass index, waist circumference, and waist-stature ratio in adults. Am. J. Clin. Nutr. 2009, 89, 500–508. [Google Scholar] [CrossRef]
- Freedman, D.S.; Goodman, A.B.; King, R.J.; Daymont, C. The Longitudinal Relation of Childhood Height to Subsequent Obesity in a Large Electronic Health Record Database. Obesity 2020, 28, 1742–1749. [Google Scholar] [CrossRef]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of the gut microbiota composition between obese and non-obese individuals in a Japanese population, as analyzed by terminal restriction fragment length polymorphism and next-generation sequencing. BMC Gastroenterol. 2015, 15, 100. [Google Scholar] [CrossRef] [Green Version]
- Logan, I.E.; Bobe, G.; Miranda, C.L.; Vasquez-Perez, S.; Choi, J.; Lowry, M.B.; Sharpton, T.J.; Morgun, A.; Maier, C.S.; Stevens, J.F.; et al. Germ-Free Swiss Webster Mice on a High-Fat Diet Develop Obesity, Hyperglycemia, and Dyslipidemia. Microorganisms 2020, 8, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.K.; Amoranto, M.B.C.; Oh, N.S.; Kim, S.; Lee, J.Y.; Oh, Y.N.; Shin, Y.K.; Yoon, Y.; Kang, D.K. Synergistic effect of Lactobacillus gasseri and Cudrania tricuspidata on the modulation of body weight and gut microbiota structure in diet-induced obese mice. Appl. Microbiol. Biotechnol. 2020, 104, 6273–6285. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.F.; Lindberg, M.; Jakobsson, H.; Bäckhed, F.; Nyrén, P.; Engstrand, L. Comparative analysis of human gut microbiota by barcoded pyrosequencing. PLoS ONE 2008, 3, e2836. [Google Scholar] [CrossRef] [PubMed]
- Rawls, J.F.; Mahowald, M.A.; Ley, R.E.; Gordon, J.I. Reciprocal gut microbiota transplants from zebrafish and mice to germ-free recipients reveal host habitat selection. Cell 2006, 127, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Bou, G.; Su, S.; Xing, J.; Qu, H.; Zhang, X.; Wang, X.; Zhao, Y.; Dugarjaviin, M. Microbial diversity within the digestive tract contents of Dezhou donkeys. PLoS ONE 2019, 14, e0226186. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Du, J.; Lu, Z.; Han, J.; Wang, Z.; Li, D.; Guan, X.; Wang, Z. Distribution of microbiota across different intestinal tract segments of a stranded dwarf minke whale, Balaenoptera acutorostrata. MicrobiologyOpen 2020, 9, e1108. [Google Scholar] [CrossRef]
- Wurm, P.; Dörner, E.; Kremer, C.; Spranger, J.; Maddox, C.; Halwachs, B.; Harrison, U.; Blanchard, T.; Haas, R.; Högenauer, C.; et al. Qualitative and Quantitative DNA- and RNA-Based Analysis of the Bacterial Stomach Microbiota in Humans, Mice, and Gerbils. mSystems 2018, 3, e00262-18. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Xiang, Y.; Zhou, W.; Chen, J.; Li, K.; Yang, H. Microbial community mapping in intestinal tract of broiler chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Araújo-Pérez, F.; McCoy, A.N.; Okechukwu, C.; Carroll, I.M.; Smith, K.M.; Jeremiah, K.; Sandler, R.S.; Asher, G.N.; Keku, T.O. Differences in microbial signatures between rectal mucosal biopsies and rectal swabs. Gut Microbes 2012, 3, 530–535. [Google Scholar] [CrossRef] [Green Version]
- Riva, A.; Borgo, F.; Lassandro, C.; Verduci, E.; Morace, G.; Borghi, E.; Berry, D. Pediatric obesity is associated with an altered gut microbiota and discordant shifts in Firmicutes populations. Environ. Microbiol. 2017, 19, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, J.; Hong, F.; Wang, S.; Jin, X.; Xue, T.; Jia, L.; Zhai, Y. Melatonin prevents obesity through modulation of gut microbiota in mice. J. Pineal Res. 2017, 62, e12399. [Google Scholar] [CrossRef] [PubMed]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Companys, J.; Gosalbes, M.J.; Pla-Pagà, L.; Calderón-Pérez, L.; Llauradó, E.; Pedret, A.; Valls, R.M.; Jiménez-Hernández, N.; Sandoval-Ramirez, B.A.; Del Bas, J.M.; et al. Gut Microbiota Profile and Its Association with Clinical Variables and Dietary Intake in Overweight/Obese and Lean Subjects: A Cross-Sectional Study. Nutrients 2021, 13, 2032. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Jeong, M.Y.; Jang, H.M.; Kim, D.H. High-fat diet causes psychiatric disorders in mice by increasing Proteobacteria population. Neurosci. Lett. 2019, 698, 51–57. [Google Scholar] [CrossRef]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [Green Version]
- Duan, R.; Guan, X.; Huang, K.; Zhang, Y.; Li, S.; Xia, J.; Shen, M. Flavonoids from Whole-Grain Oat Alleviated High-Fat Diet-Induced Hyperlipidemia via Regulating Bile Acid Metabolism and Gut Microbiota in Mice. J. Agric. Food Chem. 2021, 69, 7629–7640. [Google Scholar] [CrossRef]
- Tang, S.; Xin, Y.; Ma, Y.; Xu, X.; Zhao, S.; Cao, J. Screening of Microbes Associated With Swine Growth and Fat Deposition Traits Across the Intestinal Tract. Front. Microbiol. 2020, 11, 586776. [Google Scholar] [CrossRef]
- Panasevich, M.R.; Meers, G.M.; Linden, M.A.; Booth, F.W.; Perfield, J.W., 2nd; Fritsche, K.L.; Wankhade, U.D.; Chintapalli, S.V.; Shankar, K.; Ibdah, J.A.; et al. High-fat, high-fructose, high-cholesterol feeding causes severe NASH and cecal microbiota dysbiosis in juvenile Ossabaw swine. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E78–E92. [Google Scholar] [CrossRef]
- Sharma, S.; Tripathi, P. Gut microbiome and type 2 diabetes: Where we are and where to go? J. Nutr. Biochem. 2019, 63, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | BMI |
|---|---|
| Correlation | |
| GR value, cm | 0.37 ** |
| Thickness of backfat, cm | 0.23 ** |
| Perirenal fat weight, kg | 0.41 ** |
| Relative weight of perirenal fat, % | 0.29 ** |
| Mesenteric fat weight, kg | 0.47 ** |
| Relative weight of mesenteric fat, % | 0.33 ** |
| Tail fat weight, kg | 0.41 ** |
| Relative weight of tail fat, % | 0.14 ** |
| Total fat weight, kg | 0.54 ** |
| Relative weight of total fat, % | 0.33 ** |
| Characteristics | All Hu Sheep (n = 357) | Low BMI Hu Sheep (n = 18) | High BMI Hu Sheep (n = 18) | p-Value |
|---|---|---|---|---|
| Body mass index, kg/m2 | 89.99 ± 8.36 | 74.26 ± 3.32 | 106.34 ± 2.93 | <0.01 |
| Body weight, kg | 48.66 ± 7.23 | 37.08 ± 4.28 | 55.33 ± 4.91 | <0.01 |
| Body length, cm | 73.38 ± 3.87 | 70.56 ± 3.42 | 72.06 ± 2.48 | 0.14 |
| GR value, cm | 2.61 ± 0.52 | 1.95 ± 0.40 | 2.86 ± 0.63 | <0.01 |
| Thickness of backfat, cm | 0.63 ± 0.24 | 0.36 ± 0.12 | 0.75 ± 0.43 | <0.01 |
| Perirenal fat weight, kg | 0.69 ± 0.36 | 0.37 ± 0.17 | 0.87 ± 0.45 | <0.01 |
| Relative weight of perirenal fat, % | 1.39 ± 0.66 | 0.94 ± 0.35 | 1.57 ± 0.78 | <0.01 |
| Mesenteric fat weight, kg | 1.22 ± 0.49 | 0.79 ± 0.39 | 1.51 ± 0.49 | <0.01 |
| Relative weight of mesenteric fat, % | 2.47 ± 0.86 | 2.07 ± 0.93 | 2.73 ± 0.72 | <0.01 |
| Tail fat weight, kg | 1.56 ± 0.49 | 1.11 ± 0.54 | 1.87 ± 0.62 | <0.01 |
| Relative weight of tail fat, % | 3.17 ± 0.85 | 2.91 ± 1.41 | 3.38 ± 1.00 | 0.13 |
| Total fat weight, kg | 3.48 ± 1.06 | 2.27 ± 0.91 | 4.25 ± 1.10 | <0.01 |
| Relative weight of Total fat, % | 7.03 ± 1.69 | 5.91 ± 2.11 | 7.69 ± 1.59 | <0.01 |
| Group | A | Observed Delta | Expected Delta | Significance |
|---|---|---|---|---|
| L1–L2 | −0.0005 | 0.5305 | 0.5303 | 0.5090 |
| W1–W2 | 0.0160 | 0.6165 | 0.6266 | 0.0030 |
| B1–B2 | 0.0104 | 0.6158 | 0.6223 | 0.0170 |
| Z11–Z12 | 0.0024 | 0.5789 | 0.5803 | 0.2320 |
| S1–S2 | −0.0032 | 0.6752 | 0.6730 | 0.7090 |
| K1–K2 | 0.0056 | 0.5623 | 0.5654 | 0.1230 |
| H1–H2 | 0.0101 | 0.6228 | 0.6292 | 0.1350 |
| M1–M2 | 0.0482 | 0.4297 | 0.4514 | 0.0010 |
| J1–J2 | 0.0251 | 0.4257 | 0.4367 | 0.0010 |
| Z21–Z22 | 0.0091 | 0.5706 | 0.5758 | 0.0340 |
| Taxonomic Name | Relative Abundance | p Value | Trend | |
|---|---|---|---|---|
| Low BMI | High BMI | |||
| Oscillospiraceae_UCG-005 | 11.248% | 14.814% | 0.008 | + |
| Prevotella | 4.791% | 2.114% | 0.040 | − |
| Prevotellaceae_UCG-001 | 3.833% | 1.706% | 0.005 | − |
| Succinivibrio | 0.059% | 0.695% | 0.005 | + |
| Fibrobacter | 1.774% | 0.552% | 0.008 | − |
| Saccharofermentans | 1.283% | 0.543% | 0.005 | − |
| Succiniclasticum | 0.177% | 0.022% | 0.017 | − |
| Lachnospiraceae_UCG-010 | 0.442% | 0.724% | 0.005 | + |
| Alloprevotella | 0.002% | 0.153% | 0.005 | + |
| Butyricicoccaceae_UCG-009 | 0.458% | 0.590% | 0.022 | + |
| Methanocorpusculum | 0.124% | 0.003% | 0.005 | − |
| Agathobacter | 0.111% | 0.297% | 0.005 | + |
| Candidatus_Soleaferrea | 0.352% | 0.497% | 0.011 | + |
| Lachnospiraceae_NK3A20_group | 0.198% | 0.046% | 0.005 | − |
| Colidextribacter | 0.179% | 0.420% | 0.005 | + |
| Oscillibacter | 0.414% | 0.542% | 0.005 | + |
| Mogibacterium | 0.154% | 0.053% | 0.029 | − |
| Lachnospiraceae_UCG-002 | 0.072% | 0.146% | 0.017 | + |
| Candidatus_Saccharimonas | 0.144% | 0.049% | 0.044 | − |
| Parabacteroides | 0.076% | 0.178% | 0.019 | + |
| Prevotellaceae_UCG-003 | 0.048% | 0.142% | 0.005 | + |
| Family_XIII_AD3011_group | 0.218% | 0.303% | 0.014 | + |
| Lachnospiraceae_ND3007_group | 0.149% | 0.006% | 0.005 | − |
| Erysipelotrichaceae_UCG-009 | 0.110% | 0.050% | 0.011 | − |
| [Ruminococcus]_gauvreauii_group | 0.113% | 0.034% | 0.005 | − |
| Dorea | 0.077% | 0.114% | 0.040 | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, J.; Wang, W.; Zhang, D.; Zhang, Y.; Song, Q.; Li, X.; Zhao, Y.; Xu, D.; Zhao, L.; Li, W.; et al. Distribution and Difference of Gastrointestinal Flora in Sheep with Different Body Mass Index. Animals 2022, 12, 880. https://doi.org/10.3390/ani12070880
Cheng J, Wang W, Zhang D, Zhang Y, Song Q, Li X, Zhao Y, Xu D, Zhao L, Li W, et al. Distribution and Difference of Gastrointestinal Flora in Sheep with Different Body Mass Index. Animals. 2022; 12(7):880. https://doi.org/10.3390/ani12070880
Chicago/Turabian StyleCheng, Jiangbo, Weimin Wang, Deyin Zhang, Yukun Zhang, Qizhi Song, Xiaolong Li, Yuan Zhao, Dan Xu, Liming Zhao, Wenxin Li, and et al. 2022. "Distribution and Difference of Gastrointestinal Flora in Sheep with Different Body Mass Index" Animals 12, no. 7: 880. https://doi.org/10.3390/ani12070880
APA StyleCheng, J., Wang, W., Zhang, D., Zhang, Y., Song, Q., Li, X., Zhao, Y., Xu, D., Zhao, L., Li, W., Wang, J., Zhou, B., Lin, C., & Zhang, X. (2022). Distribution and Difference of Gastrointestinal Flora in Sheep with Different Body Mass Index. Animals, 12(7), 880. https://doi.org/10.3390/ani12070880