
Prevalence of Fox Tapeworm in Invasive Muskrats in Flanders (North Belgium)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
- What is the prevalence of EM in muskrats captured in Flanders?
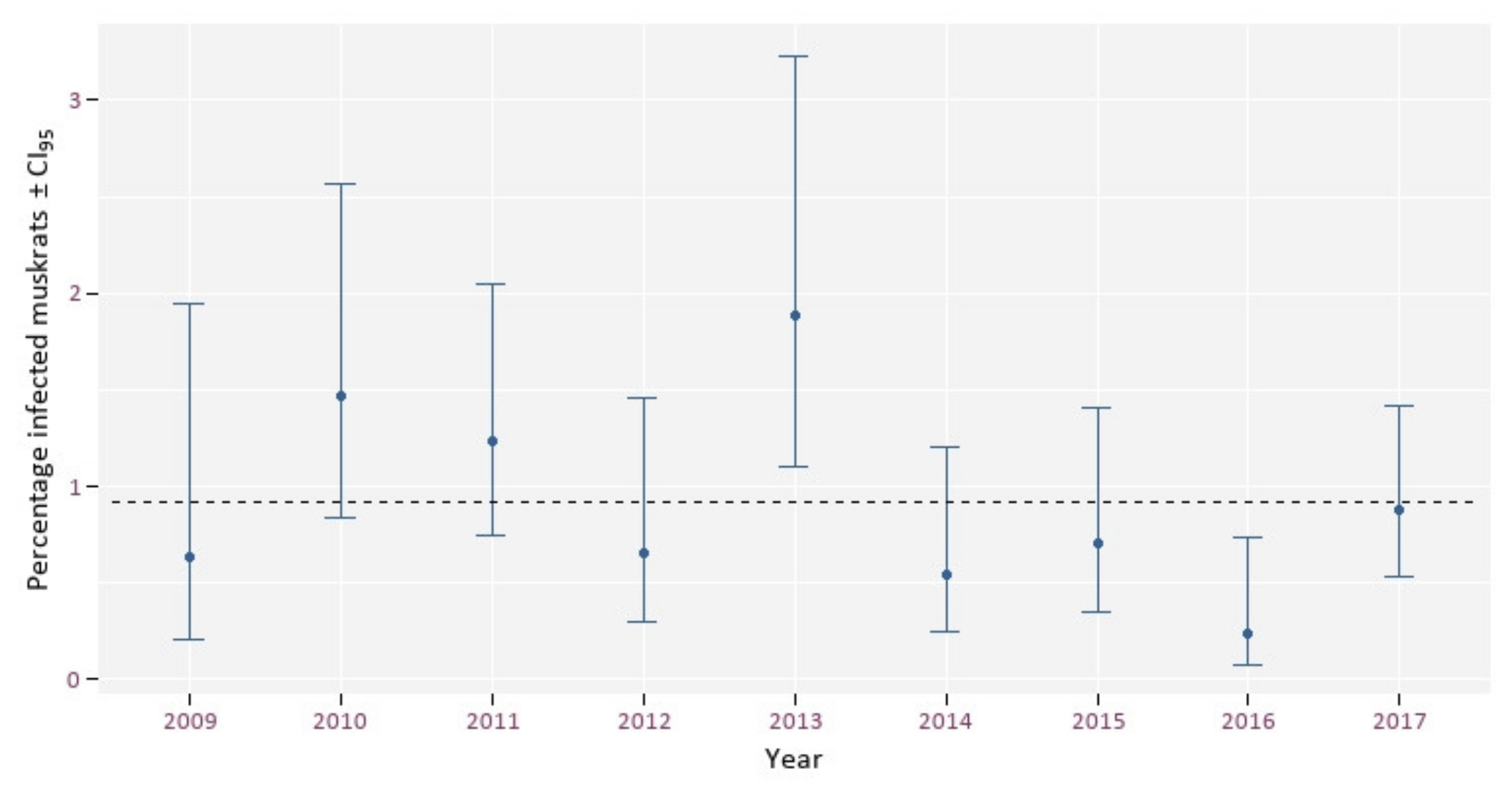
- Does this prevalence show an increase over time?
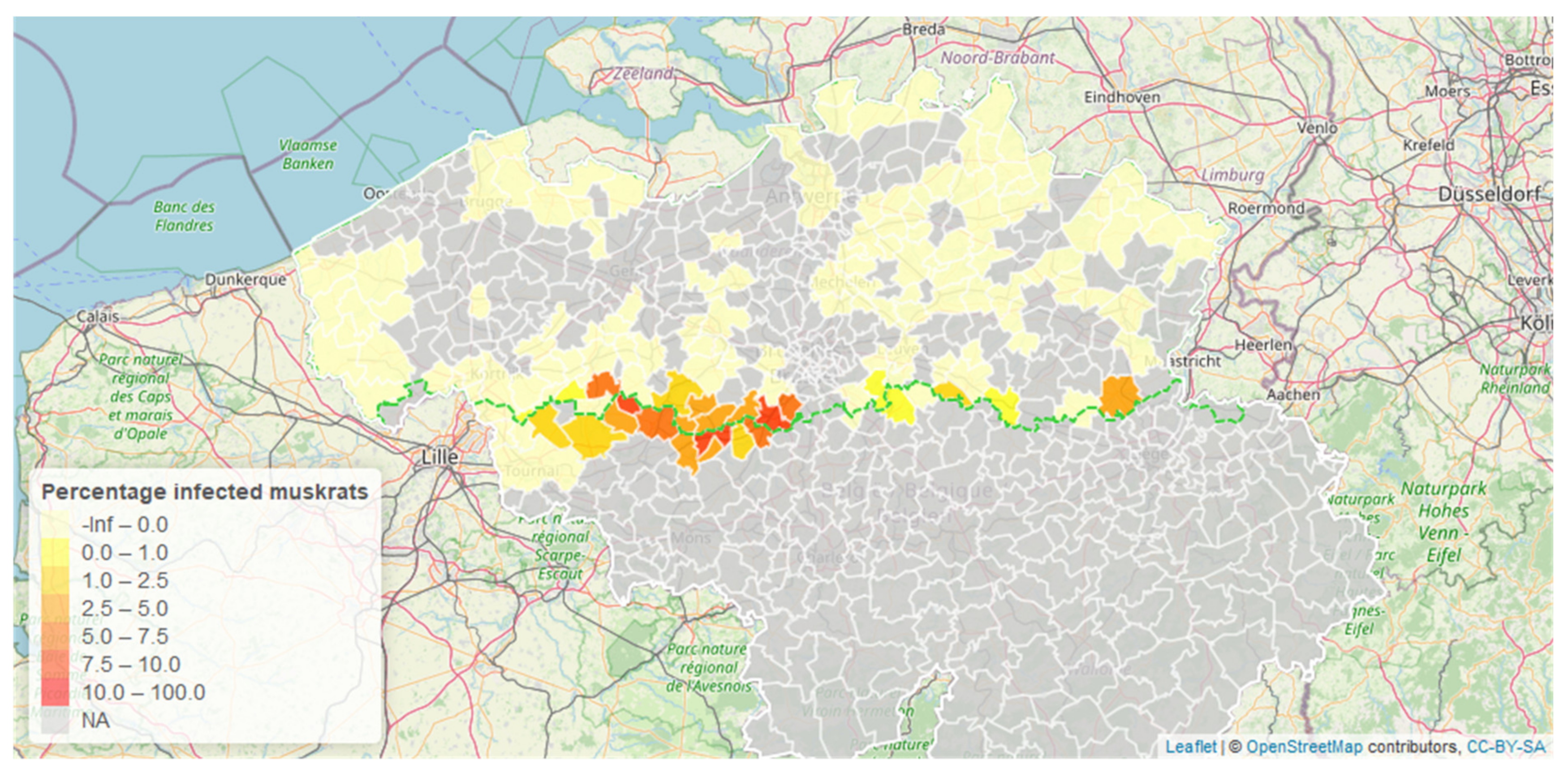
- Does the geographical range of EM infected muskrats expand?
2. Materials and Methods
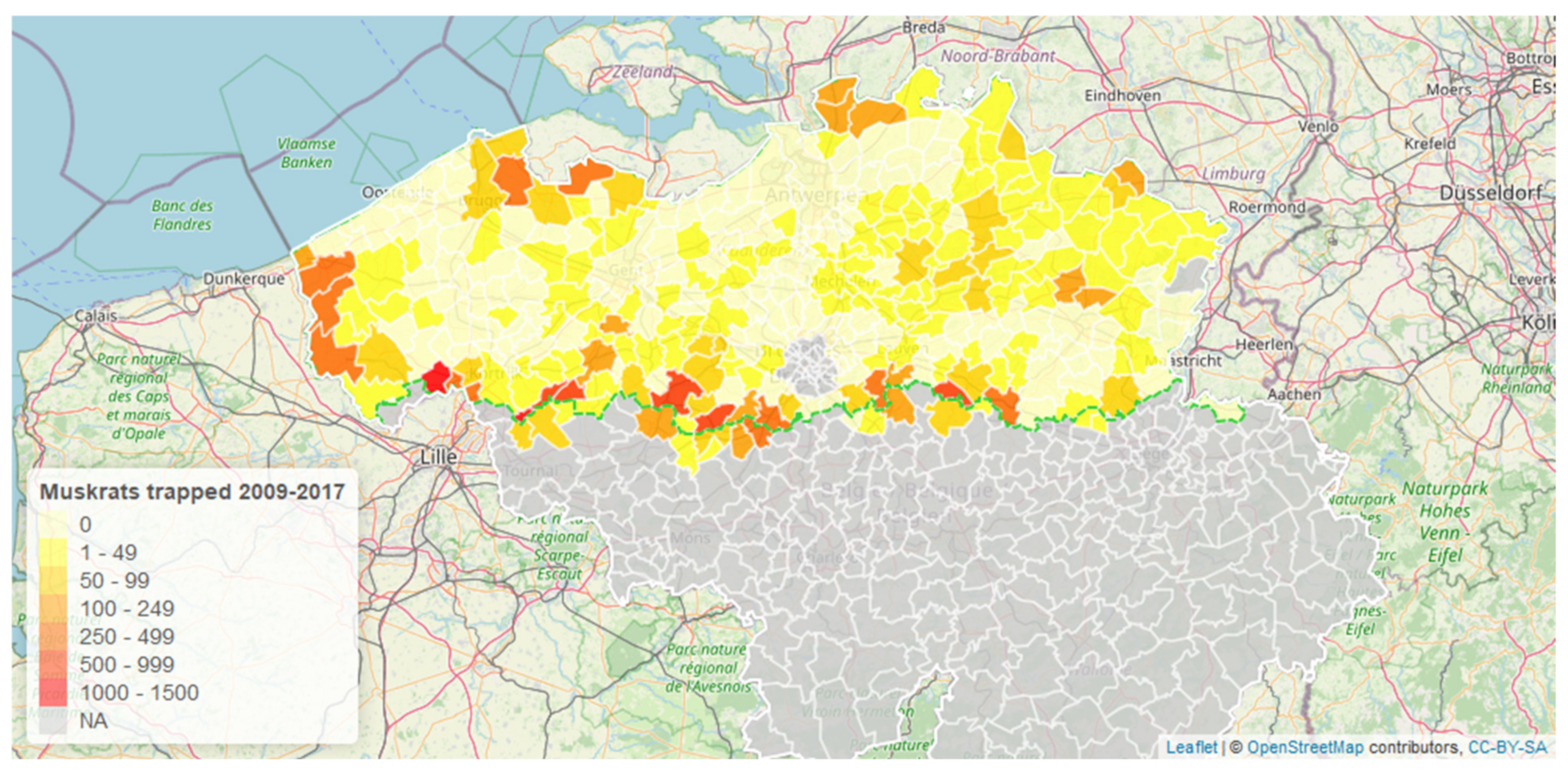
3. Results
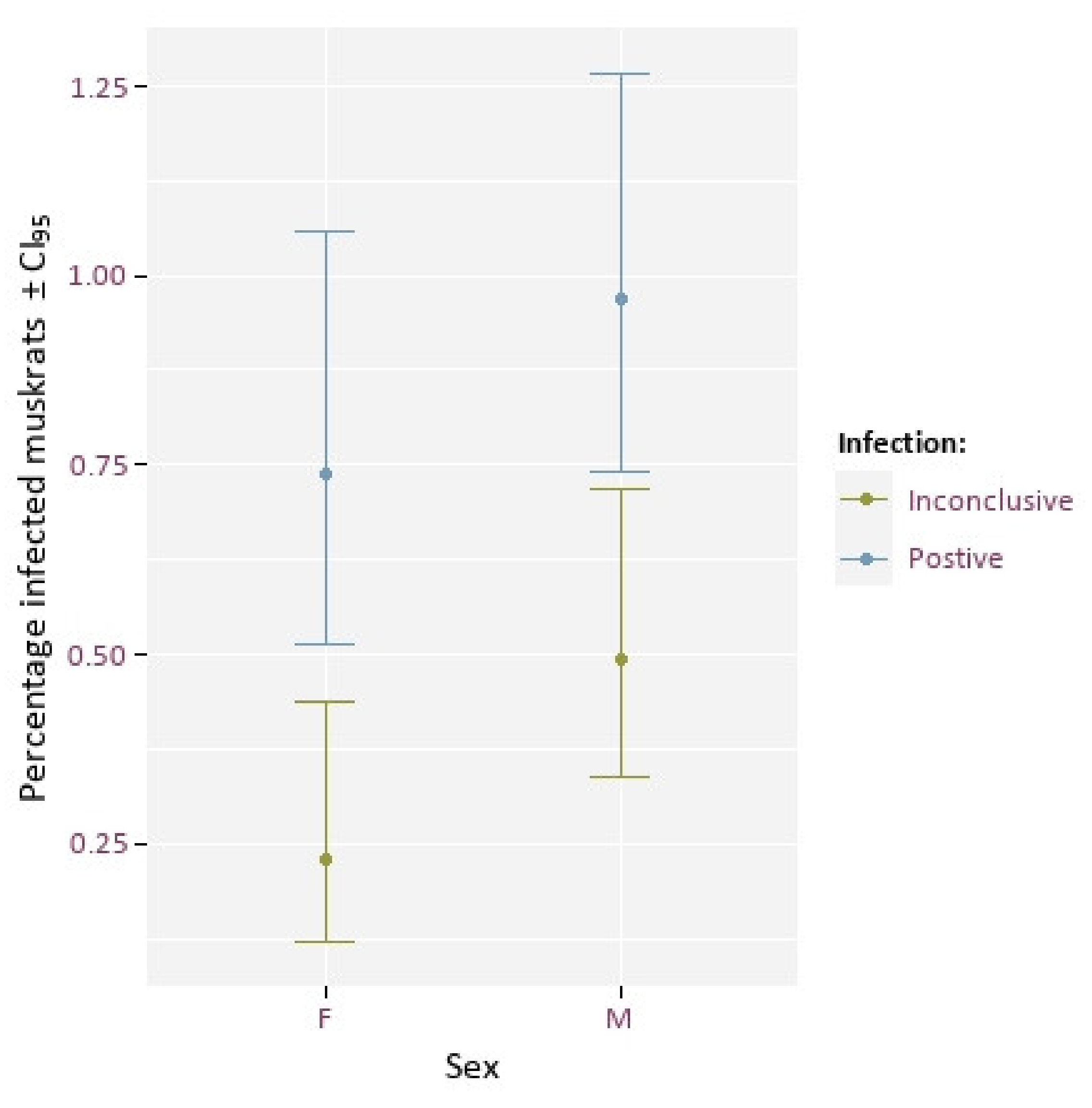
3.1. Sex Ratio and Infection Rate
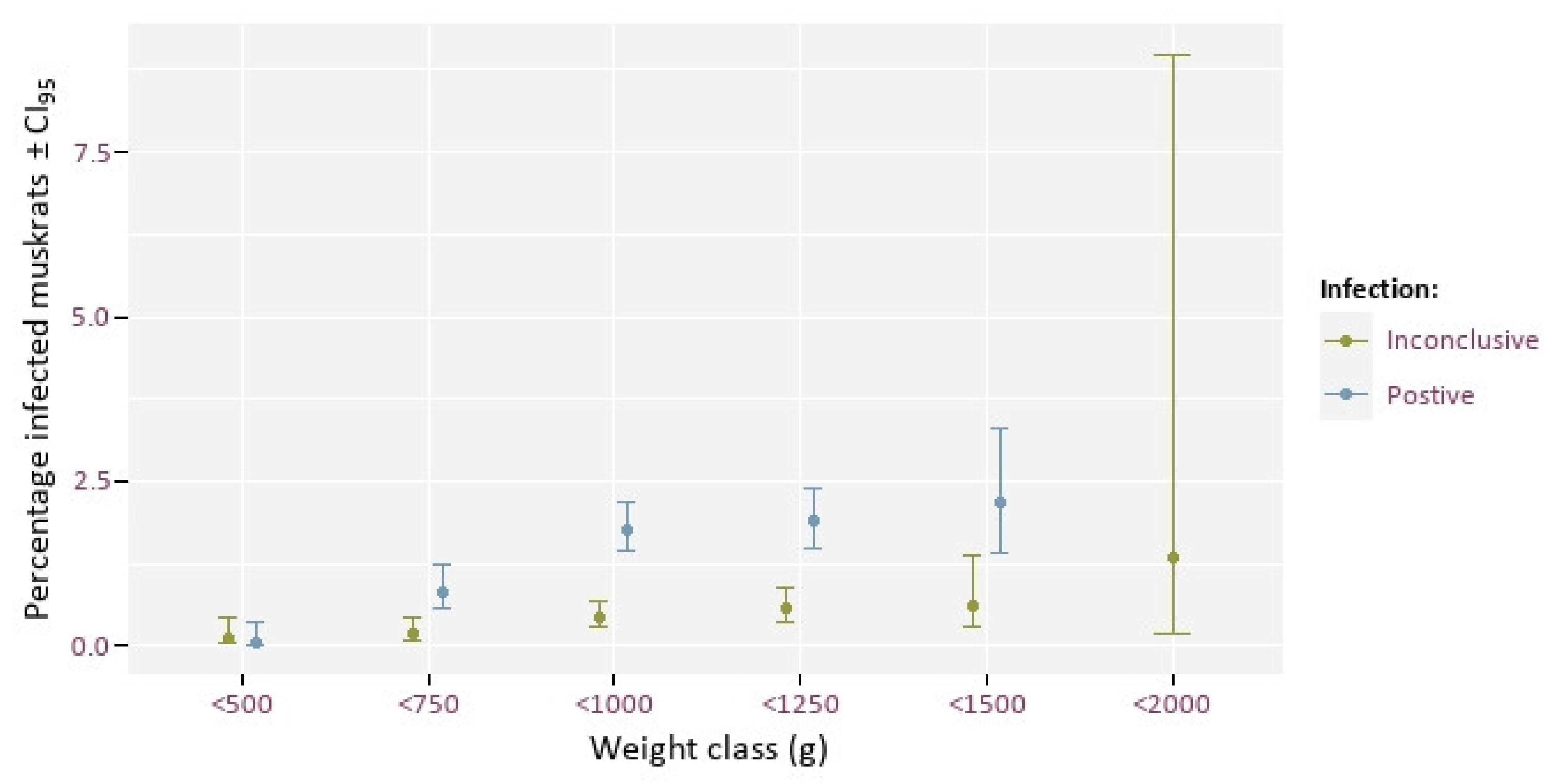
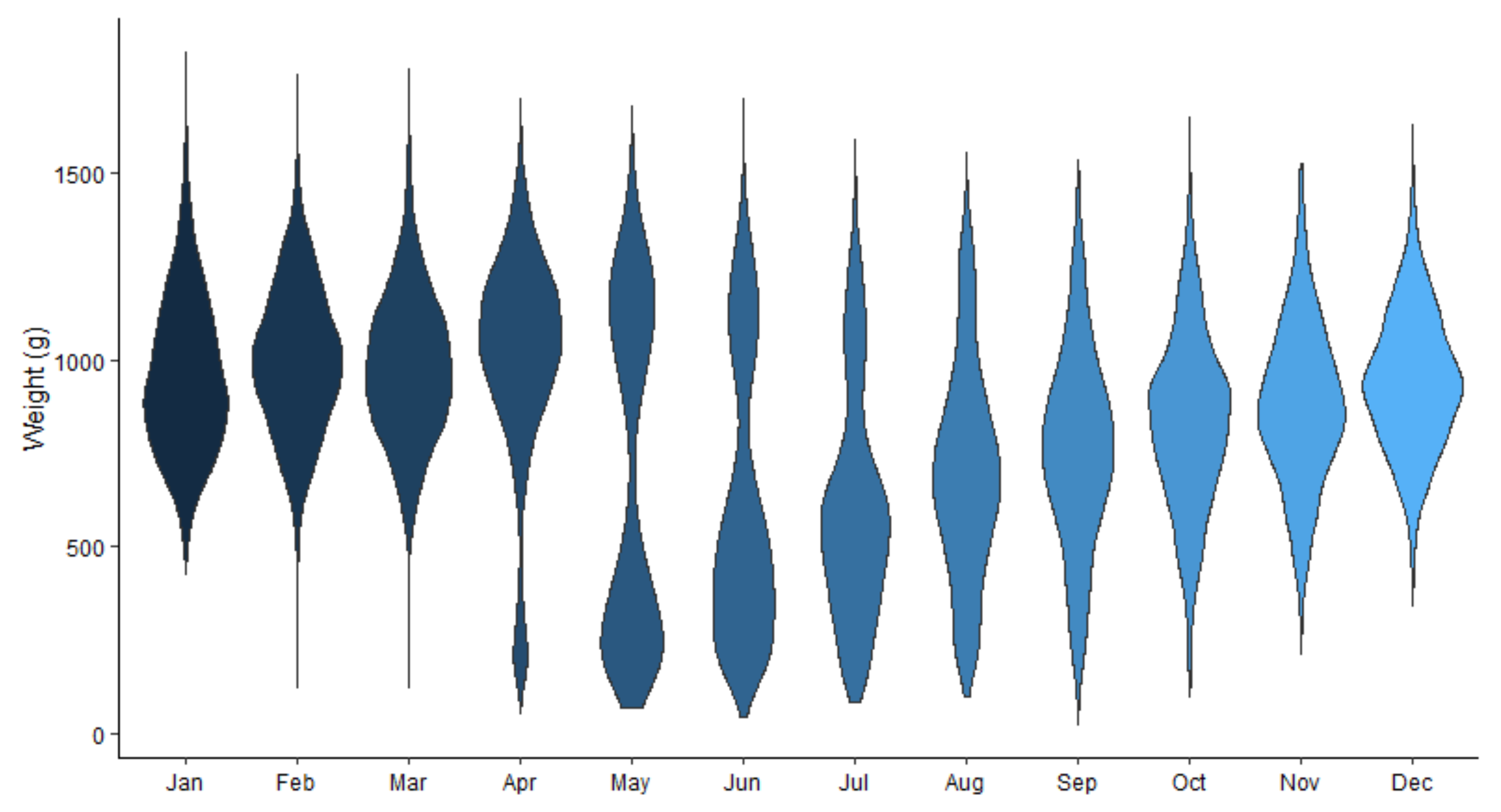
3.2. Weight/Age Distribution and Infection Rate
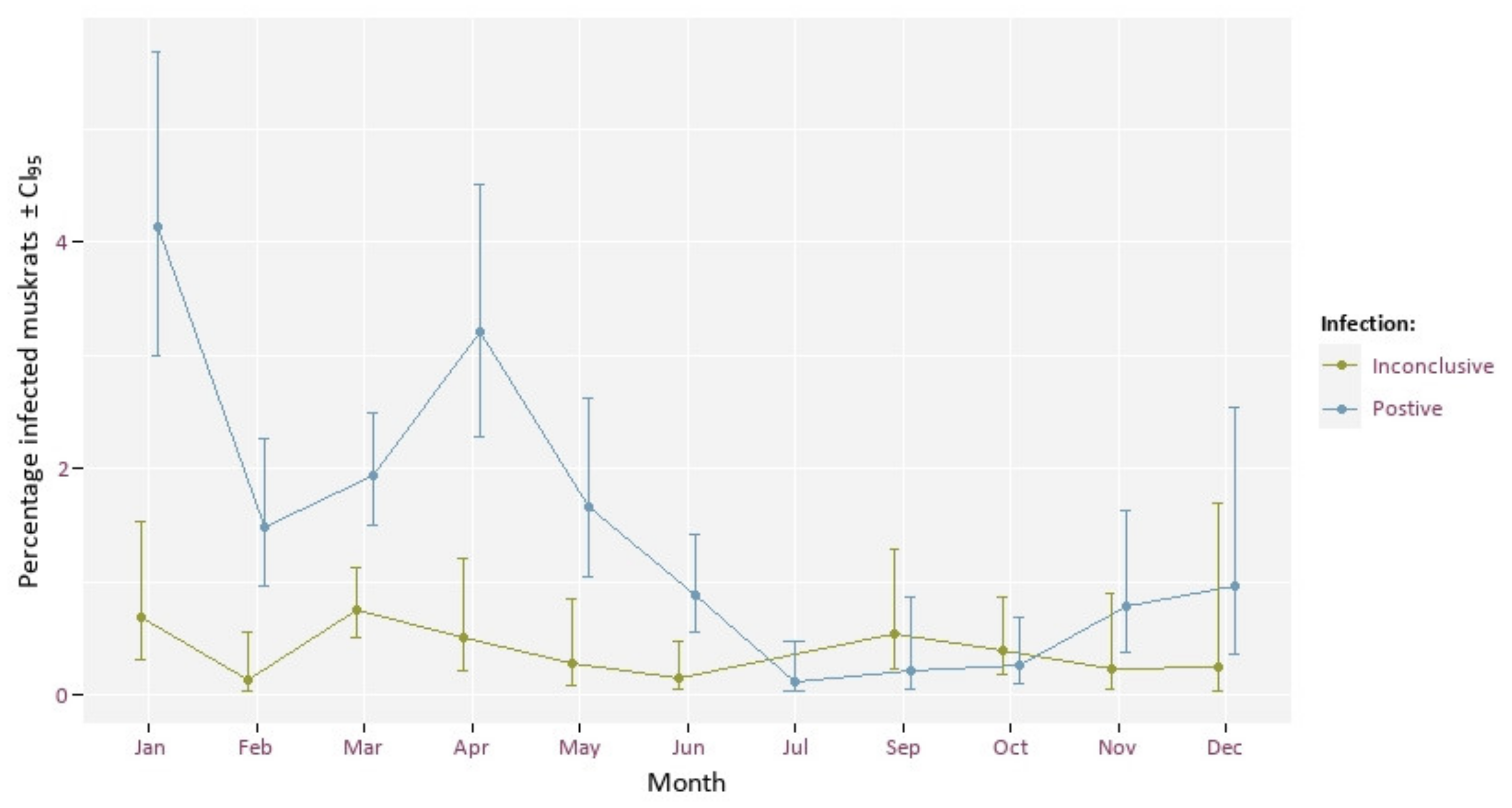
3.3. Infection Rate and Month
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roy, H.E.; Hesketh, H.; Purse, B.V.; Eilenberg, J.; Santini, A.; Scalera, R.; Stentiford, G.D.; Adriaens, T.; Bacela-Spychalska, K.; Bass, D. Alien pathogens on the horizon: Opportunities for predicting their threat to wildlife. Conserv. Lett. 2017, 10, 477–484. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Trop. 2001, 78, 103–116. [Google Scholar] [CrossRef]
- Kelly, D.W.; Paterson, R.A.; Townsend, C.R.; Poulin, R.; Tompkins, D.M. Parasite spillback: A neglected concept in invasion ecology? Ecology 2009, 90, 2047–2056. [Google Scholar] [CrossRef] [PubMed]
- Civitello, D.J.; Cohen, J.; Fatima, H.; Halstead, N.T.; Liriano, J.; McMahon, T.A.; Ortega, C.N.; Sauer, E.L.; Sehgal, T.; Young, S. Biodiversity inhibits parasites: Broad evidence for the dilution effect. Proc. Natl. Acad. Sci. USA 2015, 112, 8667–8671. [Google Scholar] [CrossRef]
- Sainsbury, A.W.; Deaville, R.; Lawson, B.; Cooley, W.A.; Farelly, S.S.; Stack, M.J.; Duff, P.; McInnes, C.J.; Gurnell, J.; Russell, P.H. Poxviral disease in red squirrels Sciurus vulgaris in the UK: Spatial and temporal trends of an emerging threat. EcoHealth 2008, 5, 305–316. [Google Scholar] [CrossRef]
- Martel, A.; Blooi, M.; Adriaensen, C.; Van Rooij, P.; Beukema, W.; Fisher, M.C.; Farrer, R.A.; Schmidt, B.R.; Tobler, U.; Goka, K. Recent introduction of a chytrid fungus endangers Western Palearctic salamanders. Science 2014, 346, 630–631. [Google Scholar] [CrossRef]
- Svoboda, J.; Mrugała, A.; Kozubíková-Balcarová, E.; Petrusek, A. Hosts and transmission of the crayfish plague pathogen Aphanomyces astaci: A review. J. Fish Dis. 2017, 40, 127–140. [Google Scholar] [CrossRef]
- Nentwig, W.; Kühnel, E.; Bacher, S. A generic impact-scoring system applied to alien mammals in Europe. Conserv. Biol. 2010, 24, 302–311. [Google Scholar] [CrossRef]
- Borgsteede, F.H.; Tibben, J.H.; van der Giessen, J.W. The musk rat (Ondatra zibethicus) as intermediate host of cestodes in the Netherlands. Vet. Parasitol. 2003, 117, 29–36. [Google Scholar] [CrossRef]
- Cartuyvels, E.; Adriaens, T.; Baert, K.; Huysentruyt, F.; Stuyck, J. Best Practice for Trapping Muskrat in Flanders.: Implementation of the Agreement on International Humane Trapping Standards; Research Institute for Nature and Forest: Brussels, Belgium, 2020. [Google Scholar]
- Cartuyvels, E.; Brosens, D.; Adriaens, T.; Baert, K.; Desmet, P.; Devisscher, S.; Neukermans, A.; Stuyck, J.; Huysentruyt, F. Muskrat Captures in Flanders, Belgium; Research Institute for Nature and Forest (INBO): Brussels, Belgium, 2021. [Google Scholar] [CrossRef]
- Oksanen, A.; Siles-Lucas, M.; Karamon, J.; Possenti, A.; Conraths, F.J.; Romig, T.; Wysocki, P.; Mannocci, A.; Mipatrini, D.; La Torre, G. The geographical distribution and prevalence of Echinococcus multilocularis in animals in the European Union and adjacent countries: A systematic review and meta-analysis. Parasites Vectors 2016, 9, 1–23. [Google Scholar] [CrossRef]
- Eckert, J.; Deplazes, P. Biological, epidemiological, and clinical aspects of echinococcosis, a zoonosis of increasing concern. Clin. Microbiol. Rev. 2004, 17, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Kapel, C.; Torgerson, P.; Thompson, R.; Deplazes, P. Reproductive potential of Echinococcus multilocularis in experimentally infected foxes, dogs, raccoon dogs and cats. Int. J. Parasitol. 2006, 36, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Eckert, J.; Gemmell, M.; Meslin, F.-X.; Pawlowski, Z.S.; World Health Organization. WHO/OIE Manual on Echinococcosis in Humans and Animals: A Public Health Problem of Global Concern; World Organisation for Animal Health: Paris, France, 2001. [Google Scholar]
- Romig, T.; Deplazes, P.; Jenkins, D.; Giraudoux, P.; Massolo, A.; Craig, P.S.; Wassermann, M.; Takahashi, K.; De La Rue, M. Ecology and life cycle patterns of Echinococcus species. Adv. Parasitol. 2017, 95, 213–314. [Google Scholar] [PubMed]
- Miller, A.L.; Olsson, G.E.; Walburg, M.R.; Sollenberg, S.; Skarin, M.; Ley, C.; Wahlström, H.; Höglund, J. First identification of Echinococcus multilocularis in rodent intermediate hosts in Sweden. Int. J. Parasitol. Parasites Wildl. 2016, 5, 56–63. [Google Scholar] [CrossRef]
- Reperant, L.; Hegglin, D.; Tanner, I.; Fischer, C.; Deplazes, P. Rodents as shared indicators for zoonotic parasites of carnivores in urban environments. Parasitology 2009, 136, 329–337. [Google Scholar] [CrossRef]
- Umhang, G.; Richomme, C.; Boucher, J.-M.; Guedon, G.; Boué, F. Nutrias and muskrats as bioindicators for the presence of Echinococcus multilocularis in new endemic areas. Vet. Parasitol. 2013, 197, 283–287. [Google Scholar] [CrossRef]
- Hanosset, R.; Saegerman, C.; Adant, S.; Massart, L.; Losson, B. Echinococcus multilocularis in Belgium: Prevalence in red foxes (Vulpes vulpes) and in different species of potential intermediate hosts. Vet. Parasitol. 2008, 151, 212–217. [Google Scholar] [CrossRef]
- Craig, P.; Hegglin, D.; Lightowlers, M.; Torgerson, P.R.; Wang, Q. Echinococcosis: Control and prevention. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 96, pp. 55–158. [Google Scholar]
- Vuitton, D.; Demonmerot, F.; Knapp, J.; Richou, C.; Grenouillet, F.; Chauchet, A.; Vuitton, L.; Bresson-Hadni, S.; Millon, L. Clinical epidemiology of human AE in Europe. Vet. Parasitol. 2015, 213, 110–120. [Google Scholar] [CrossRef]
- Chautan, M.; Pontier, D.; Artois, M. Role of rabies in recent demographic changes in red fox (Vulpes vulpes) populations in Europe. Mammalia 2000, 64, 391–410. [Google Scholar] [CrossRef]
- Van Den Berge, K.; De Pauw, W. Vos Vulpes vulpes (Linnaeus, 1758). In Zoogdieren in Vlaanderen. Ecologie En Verspreiding van 1987 Tot 2002; Verkem, S., De Maeseneer, J., Vandendriessche, B., Verbeylen, G., Yskout, S., Eds.; Natuurpunt Studie en JNM-Zoogdierenwerkgroep: Mechelen, Belgium, 2003; pp. 363–369. [Google Scholar]
- Van Den Berge, K.; Gouwy, J.; Berlengee, F.; Vansevenant, D. Oriënterende Verkenning naar de Stadsvos in Vlaanderen; INBO.R.2013.1336286; Instituut voor Natuur- en Bosonderzoek: Brussels, Belgium, 2013. [Google Scholar]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar]
- Eckert, J.; Thompson, R. Historical aspects of echinococcosis. Adv. Parasitol. 2017, 95, 1–64. [Google Scholar] [PubMed]
- Karamon, J.; Kochanowski, M.; Sroka, J.; Cencek, T.; Różycki, M.; Chmurzyńska, E.; Bilska-Zając, E. The prevalence of Echinococcus multilocularis in red foxes in Poland--current results (2009–2013). Parasitol. Res. 2014, 113, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Combes, B.; Comte, S.; Raton, V.; Raoul, F.; Boué, F.; Umhang, G.; Favier, S.; Dunoyer, C.; Woronoff, N.; Giraudoux, P. Westward spread of Echinococcus multilocularis in foxes, France, 2005–2010. Emerg. Infect. Dis. 2012, 18, 2059. [Google Scholar] [CrossRef]
- Takumi, K.; de Vries, A.; Chu, M.L.; Mulder, J.; Teunis, P.; van der Giessen, J. Evidence for an increasing presence of Echinococcus multilocularis in foxes in The Netherlands. Int. J. Parasitol. 2008, 38, 571–578. [Google Scholar] [CrossRef]
- Vervaeke, M.; van der Giessen, J.; Brochier, B.; Losson, B.; Jordaens, K.; Verhagen, R.; de Lezenne Coulander, C.; Teunis, P. Spatial spreading of Echinococcus multilocularis in Red foxes (Vulpes vulpes) across nation borders in Western Europe. Prev. Vet. Med. 2006, 76, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Deplazes, P.; Rinaldi, L.; Rojas, C.A.; Torgerson, P.; Harandi, M.; Romig, T.; Antolova, D.; Schurer, J.; Lahmar, S.; Cringoli, G. Global distribution of alveolar and cystic echinococcosis. Adv. Parasitol. 2017, 95, 315–493. [Google Scholar] [PubMed]
- Jansen, F.; Claes, M.; Bakkers, E.; Aryal, A.; Madimba, K.; Gabriël, S.; Dermauw, V.; Van Hul, A.; Vervaeke, M.; Dorny, P. Echinococcus multilocularis in red foxes in North Belgium: Prevalence and trends in distribution. Vet. Parasitol. Reg. Stud. Rep. 2020, 22, 100470. [Google Scholar] [CrossRef]
- Van Gucht, S.; Van Den Berge, K.; Quataert, P.; Verschelde, P.; Le Roux, I. No emergence of Echinococcus multilocularis in foxes in flanders and Brussels anno 2007–2008. Zoonoses Public Health 2010, 57, e65–e70. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 3.5.1; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Zhenghuan, W.; Xiaoming, W.; Xiaoqing, L. Echinococcosis in China, a review of the epidemiology of Echinococcus spp. Ecohealth 2008, 5, 115–126. [Google Scholar] [CrossRef]
- Adriaens, T.; Branquart, E.; Gosse, D.; Reniers, J.; Vanderhoeven, S. Feasibility of Eradication and Spread Limitation for Species of Union Concern Sensu the EU IAS Regulation (EU 1143/2014) in Belgium; Institute for Nature and Forest Research, Service Public de Wallonie, National Scientific Secretariat on Invasive Alien Species, Belgian Biodiversity Platform: Brussels, Belgium, 2019. [Google Scholar]
- Plug, K. Handboek Muskusrattenbestrijding; Landbouw en Visserij: Directie Natuur, Milieu en Faunabeheer: The Hague, The Netherlands, 1988. [Google Scholar]
- Mathy, A.; Hanosset, R.; Adant, S.; Losson, B. The carriage of larval Echinococcus multilocularis and other cestodes by the musk rat (Ondatra zibethicus) along the Ourthe River and its tributaries (Belgium). J. Wildl. Dis. 2009, 45, 279–287. [Google Scholar] [CrossRef]
- Knapp, J.; Bart, J.-M.; Giraudoux, P.; Glowatzki, M.-L.; Breyer, I.; Raoul, F.; Deplazes, P.; Duscher, G.; Martinek, K.; Dubinsky, P. Genetic diversity of the cestode Echinococcus multilocularis in red foxes at a continental scale in Europe. PLoS Negl. Trop. Dis. 2009, 3, e452. [Google Scholar] [CrossRef] [PubMed]
- Maes, D.; Baert, K.; Boers, K.; Casaer, J.; Criel, D.; Crevecoeur, L.; Dekeukeleire, D.; Gouwy, J.; Gyselings, R.; Haelters, J.; et al. De Iucn Rode Lijst van de Zoogdieren in Vlaanderen.; INBO.R.2014.1828211; Het Instituut Voor Natuur-En Bosonderzoek: Brussels, Belgium, 2014. [Google Scholar]
- Libois, R. The non flying mammals of the walloon region: Populations trends; AGRISl: Charente, France, 2006. [Google Scholar]
- Lloyd, H.G. The Red Fox; BT Batsford: London, UK, 1980. [Google Scholar]
- Keune, H.; Kretsch, C.; De Blust, G.; Gilbert, M.; Flandroy, L.; Van den Berge, K.; Versteirt, V.; Hartig, T.; De Keersmaecker, L.; Eggermont, H. Science–policy challenges for biodiversity, public health and urbanization: Examples from Belgium. Environ. Res. Lett. 2013, 8, 025015. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cartuyvels, E.; Adriaens, T.; Baert, K.; Huysentruyt, F.; Van Den Berge, K. Prevalence of Fox Tapeworm in Invasive Muskrats in Flanders (North Belgium). Animals 2022, 12, 879. https://doi.org/10.3390/ani12070879
Cartuyvels E, Adriaens T, Baert K, Huysentruyt F, Van Den Berge K. Prevalence of Fox Tapeworm in Invasive Muskrats in Flanders (North Belgium). Animals. 2022; 12(7):879. https://doi.org/10.3390/ani12070879
Chicago/Turabian StyleCartuyvels, Emma, Tim Adriaens, Kristof Baert, Frank Huysentruyt, and Koen Van Den Berge. 2022. "Prevalence of Fox Tapeworm in Invasive Muskrats in Flanders (North Belgium)" Animals 12, no. 7: 879. https://doi.org/10.3390/ani12070879
APA StyleCartuyvels, E., Adriaens, T., Baert, K., Huysentruyt, F., & Van Den Berge, K. (2022). Prevalence of Fox Tapeworm in Invasive Muskrats in Flanders (North Belgium). Animals, 12(7), 879. https://doi.org/10.3390/ani12070879