
The Effects of Protease Supplementation and Faba Bean Extrusion on Growth, Gastrointestinal Tract Physiology and Selected Blood Indices of Weaned Pigs

 ,
,  ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Faba Bean Seeds
2.2. Animals, Diets, and Protease
2.3. Blood Sampling
2.4. Metabolic Blood Serum Profile
2.5. Enzyme Activity
2.6. Chemical Analysis
2.7. Morphometric Analysis
2.8. Statistical Analysis
3. Results
3.1. The Chemical Composition of Faba Bean Seeds
3.2. Animal Experiment
4. Discussion
4.1. Extrusion Effect
4.2. Protease Effect
4.3. Extrusion and Protease Effect
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuśnio, A.; Taciak, M.; Barszcz, M.; Święch, E.; Bachanek, I.; Skomiał, J. Effect of replacing soybean meal by raw or extruded pea seeds on growth performance and selected physiological parameters of the ileum and distal colon of pigs. PLoS ONE 2017, 12, e0169467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaworska, A.; Kasprowicz-Potocka, M.; Rutkowski, A.; Jamroz, D. The influence of dietary raw and extruded field peas (Pisum sativum L.) on nutrients digestibility and performance of weaned and fattening pigs. J. Anim. Feed Sci. 2018, 27, 123–130. [Google Scholar] [CrossRef]
- Zaworska-Zakrzewska, A.; Kasprowicz-Potocka, M.; Twarużek, M.; Kosicki, R.; Grajewski, J.; Wiśniewska, Z.; Rutkowski, A. A comparison of the composition and contamination of soybean cultivated in Europe and limitation of raw soy seed content in weaned pigs’ diets. Animals 2020, 10, 1972. [Google Scholar] [CrossRef] [PubMed]
- Gulewicz, P.; Martinez-Villaluenga, C.; Kasprowicz-Potocka, M.; Frias, J. Non-nutritive compounds in fabaceae family seeds and the improvement of their nutritional quality by traditional processing—A Review. Pol. J. Food Nutr. Sci. 2014, 64, 75. [Google Scholar] [CrossRef] [Green Version]
- Kasprowicz-Potocka, M.; Zaworska, A.; Kaczmarek, S.A.; Rutkowski, A. The nutritional value of narrow-leafed lupine (Lupinus angustifolius) for fattening pigs. Arch. Anim. Nutr. 2016, 70, 209–223. [Google Scholar] [CrossRef]
- Kasprowicz-Potocka, M.; Zaworska, A.; Kaczmarek, S.; Hejdysz, M.; Mikuła, R.; Rutkowski, A. The effect of Lupinus albus seeds on digestibility, performance and gastrointestinal tract indices in pigs. J. Anim. Physiol. Anim. Nutr. 2017, 10, e216–e224. [Google Scholar] [CrossRef]
- Rahate, K.A.; Madhumita, M.; Prabhakar, P.K. Nutritional composition, anti-nutritional factors, pretreatments-cum-processing impact and food formulation potential of faba bean (Vicia faba L.): A comprehensive review. LWT 2021, 138, 110796. [Google Scholar] [CrossRef]
- Hejdysz, M.; Kaczmarek, S.A.; Rutkowski, A. Extrusion cooking improves the metabo11able energy of faba beans and the amino acid digestibility in broilers. Anim. Feed Sci. Technol. 2016, 212, 100–111. [Google Scholar] [CrossRef]
- Tuśnio, A.; Barszcz, M.; Taciak, M.; Święch, E.; Wójtowicz, A.; Skomiał, J. The effect of a diet containing extruded faba bean seeds on growth performance and selected microbial activity indices in the large intestine of piglets. Animals 2021, 11, 1703. [Google Scholar] [CrossRef]
- Ivarsson, E.; Neil, M. Variations in nutritional and antinutritional contents among faba bean cultivars and effects on growth performance of weaner pigs. Livest. Sci. 2018, 212, 14–21. [Google Scholar] [CrossRef]
- Riaz, M.N. Extrusion processing of oilseed meals for food and feed production. In Bailey’s Industrial Oil and Fat Products; Shahidi, F., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2005; pp. 533–572. [Google Scholar] [CrossRef]
- Huss, A.; Cochrane, R.; Jones, C.; Atungulu, G.G. Physical and chemical methods for the reduction of biological hazards in animal feeds. In Food and Feed Safety Systems and Analysis; Academic Press: Cambridge, MA, USA, 2018; pp. 83–95. [Google Scholar]
- Li, Y.; Fang, Z.; Dai, J.; Partridge, G.; Ru, Y.; Peng, J. Corn extrusion and enzyme addition improves digestibility of corn/soyabased diets by pigs: In vitro and in vivo studies. Anim. Feed Sci. Technol. 2010, 158, 146–154. [Google Scholar]
- Zuo, J.; Ling, B.; Long, L.; Li, T.; Lahaye, L.; Yang, C.; Feng, D. Effect of dietary supplementation with protease on growth performance, nutrient digestibility, intestinal morphology, digestive enzymes and gene expression of weaned piglets. Anim. Nutr. 2015, 1, 276–282. [Google Scholar] [CrossRef]
- Hegazy, H.S.; El-Bedawey, A.E.A.; Rahma, E.H.; Gaafar, A.M. Effect of extrusion process on nutritional, functional properties and antioxidant activity of germinated chickpea incorporated corn extrudates. Am. J. Food Sci. Nutr. Res. 2017, 4, 59–66. [Google Scholar]
- O’Doherty, J.V.; Keady, U. The effect of expander processing and extrusion on the nutritive value of peas for pigs. Anim. Sci. 2001, 72, 43–53. [Google Scholar] [CrossRef]
- Gilani, G.S.; Xiao, C.W.; Cockell, K.A. Impact of antinutritional factors in food proteins on the digestibility of protein and the bioavailability of amino acids and on protein quality. Br. J. Nutr. 2012, 108, 315–332. [Google Scholar] [CrossRef]
- Pasqualone, A.; Costantini, M.; Coldea, T.E.; Summo, C. Use of legumes in extrusion cooking: A review. Foods 2020, 9, 958. [Google Scholar] [CrossRef]
- Kamel, N.F.; Ragaa, M.; El-Banna, R.A.; Mohamed, F.F. Effects of a monocomponent protease on performance parameters and protein digestibility in broiler chickens. Agric. Agric. Sci. Proc. 2015, 6, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Cowieson, A.J.; Roos, F.F. Toward optimal value creation through the application of exogenous mono-component protease in the diets of non-ruminants. Anim. Feed Sci. Technol. 2016, 221, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Hanczakowska, E.; Swiatkiewicz, M. Legume seeds and rapeseed press cake as replacers of soybean meal in feed for fattening pigs. Ann. Anim. Sci. 2014, 14, 921. [Google Scholar]
- EU. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union L 2010, 276, 33–79. [Google Scholar]
- GfE. Recommendations for the Energy and Nutrient Supply for Pigs; DLG-Verlag: Frankfurt am Main, Germany, 2006; ISBN 978-3769006834. (In German) [Google Scholar]
- Adeola, O. Digestion and balance techniques in pigs. In Swine Nutrition, 2nd ed.; Lewis, A.J., Southern, L.L., Eds.; CRC Press: Washington, DC, USA, 2001; pp. 903–916. [Google Scholar]
- Pruszynska-Oszmalek, E.; Kolodziejski, P.; Stadnicka, K.; Sassek, M.; Chalupka, D.; Kuston, B.; Nogowski, L.; Mackowiak, P.; Maiorano, G.; Jankowski, J. In ovo injection of prebiotics and synbiotics affects the digestive potency of the pancreas in growing chickens. Poult. Sci. 2015, 94, 1909–1916. [Google Scholar] [CrossRef] [PubMed]
- AOAC-Association of Official Analytical Chemists. Official Methods of Analysis, Agricultural Chemicals, 19th ed; Gaithersburg: Maryland, VA, USA, 2007; pp. 46–48. [Google Scholar]
- Englyst, H.N.; Kingman, S.M.; Hudson, G.J.; Cummings, J.H. Measurement of resistant starch in vitro and in vivo. Br. J. Nutr. 1996, 75, 749–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weurding, R.E.; Veldman, A.; Veen, W.A.G.; van der Aar, P.J.; Verstegen, M.W.A. In vitro starch digestion correlates well with rate and extent of starch digestion in broiler chickens. J. Nutr. 2001, 131, 2336–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhla, S.; Ebmeier, C. Untersuchungen zum Tanningehalt in Ackerbohnen. Arch. Tierernaehr. 1981, 31, 573. [Google Scholar] [CrossRef]
- PN-EN ISO 14902:2005. Animal Feeding Stuffs—Determination of Trypsin Inhibitor Activity of Soya Products; Polish Committee for Standardization: Warsaw, Poland, 2005; pp. 1–14. (In Polish) [Google Scholar]
- Zalewski, K.; Lahuta, L.; Horbowicz, M. The effect of soil drought on the composition of carbohydrates in yellow lupin seeds and triticale kernels. Acta Physiol. Plant. 2001, 23, 73–78. [Google Scholar] [CrossRef]
- Haug, W.; Lantzsch, H.J. Sensitive method for the rapid determination of phytate in cereals and cereal products. J. Sci. Food Agric. 1983, 34, 1423–1426. [Google Scholar] [CrossRef]
- Englyst, H.N.; Cummings, J.H. Simplified method for the measurement of total non-starch polysaccharides by gas-liquid chromatography of constitiuent sugars as alditol acetates. Analyst 1984, 109, 937–942. [Google Scholar] [CrossRef]
- Englyst, H.N.; Cummings, J.H. Improved method for measurement of dietary fiber as non-starch polysaccharides in plant foods. AOAC 1988, 71, 808–814. [Google Scholar] [CrossRef]
- Slominski, B.A.; Campbell, L. Non-starch polysaccharides of canola meal. Quantification, digestibility in poultry and potential benefit of dietary enzyme supplementation. J. Sci. Food Agric. 1990, 53, 175–184. [Google Scholar] [CrossRef]
- Scott, R.W. Colorimetric determination of hexuronic acids in plant materials. Anal. Chem. 1979, 51, 936–941. [Google Scholar] [CrossRef]
- Short, F.J.; Gorton, P.; Wiseman, J.; Boorman, K.N. Determination of titanium dioxide added as an inert marker in chicken digestibility studies. Anim. Feed Sci. Technol. 1996, 59, 215–221. [Google Scholar] [CrossRef]
- Myers, W.D.; Ludden, P.A.; Nayigihugu, V.; Hess, B.W. Technical note. A procedure for the preparation and quantitative analysis of samples for titanium dioxide. J. Anim. Sci. 2004, 82, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Stuper-Szablewska, K.; Buśko, M.; Góral, T.; Perkowski, J. The fatty acid profile in different wheat cultivars depending on the level of contamination with microscopic fungi. Food Chem. 2014, 153, 216–223. [Google Scholar] [CrossRef]
- Punia, S.; Dhull, S.B.; Sandhu, K.S.; Kaur, M. Faba bean (Vicia faba) starch: Structure, properties, and in vitro digestibility—A review. Legume Sci. 2019, 1, e18. [Google Scholar] [CrossRef] [Green Version]
- Gulati, P.; Brahma, S.; Rose, D.J. Impacts of extrusion processing on nutritional components in cereals and legumes: Carbohydrates, proteins, lipids, vitamins, and minerals. In Extrusion Cooking; Woodhead Publishing: Sawston, UK, 2020; pp. 415–443. [Google Scholar]
- Dhull, S.B.; Kidwai, M.K.; Noor, R.; Chawla, P.; Rose, P.K. A review of nutritional profile and processing of faba bean (Vicia faba L.). Legume Sci. 2021. [Google Scholar] [CrossRef]
- Meng, Z.; Liu, Q.; Zhang, Y.; Chen, J.; Sun, Z.; Ren, C.; Huang, Y. Nutritive value of faba bean (Vicia faba L.) as a feedstuff resource in livestock nutrition: A review. Food Sci. Nutr. 2021, 9, 5244–5262. [Google Scholar] [CrossRef]
- Biagia, G.; Cipollini, I.; Paulicks, B.R.; Roth, F.X. Effect of tannins on growth performance and intestinal ecosystem in weaned piglets. Arch. Anim. Nutr. 2010, 64, 121–135. [Google Scholar] [CrossRef]
- Kasiga, T.; Karki, B.; Croat, J.; Kaur, J.; Gibbons, W.R.; Muthukumarappan, K.; Brown, M.L. Process effects on carinata Brassica carinata and camelina Camelina sativa seed meal compositions and diet palatability in Rainbow Trout Oncorhynchus mykiss. Anim. Feed Sci. Technol. 2020, 267, 114578. [Google Scholar] [CrossRef]
- Rodrigues, E.A.; Badiola, I.; Francesch, M.; Torrallardona, D. Effect of cereal extrusion on performance, nutrient digestibility, and cecal fermentation in weanling pigs. J. Anim. Sci. 2016, 94, 298–302. [Google Scholar] [CrossRef]
- Lancheros, J.P.; Espinosa, C.D.; Stein, H.H. Effects of particle size reduction, pelleting, and extrusion on the nutritional value of ingredients and diets fed to pigs: A review. Anim. Feed Sci. Technol. 2020, 268, 114603. [Google Scholar] [CrossRef]
- Świątkiewicz, M.; Księżak, J.; Hanczakowska, E. The effect of feeding native faba bean seeds (Vicia faba L.) to sows and supplemented with enzymes to piglets and growing pigs. Ann. Anim. Sci. 2018, 18, 1007–1027. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Lv, Y.; Guo, L.; Wang, Z.; Zhao, F. Effects of three kinds of protease on growth performance, apparent digestibility of nutrients and caecal microbial counts in weanling pigs. Czech. J. Anim. Sci. 2020, 65, 373–379. [Google Scholar] [CrossRef]
- Tactacan, G.B.; Cho, S.Y.; Cho, J.H.; Kim, I.H. Performance Responses, Nutrient Digestibility, Blood Characteristics, and Measures of Gastrointestinal Health in Weanling Pigs Fed Protease Enzyme. Asian-Australas. J. Anim Sci. 2016, 29, 998–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
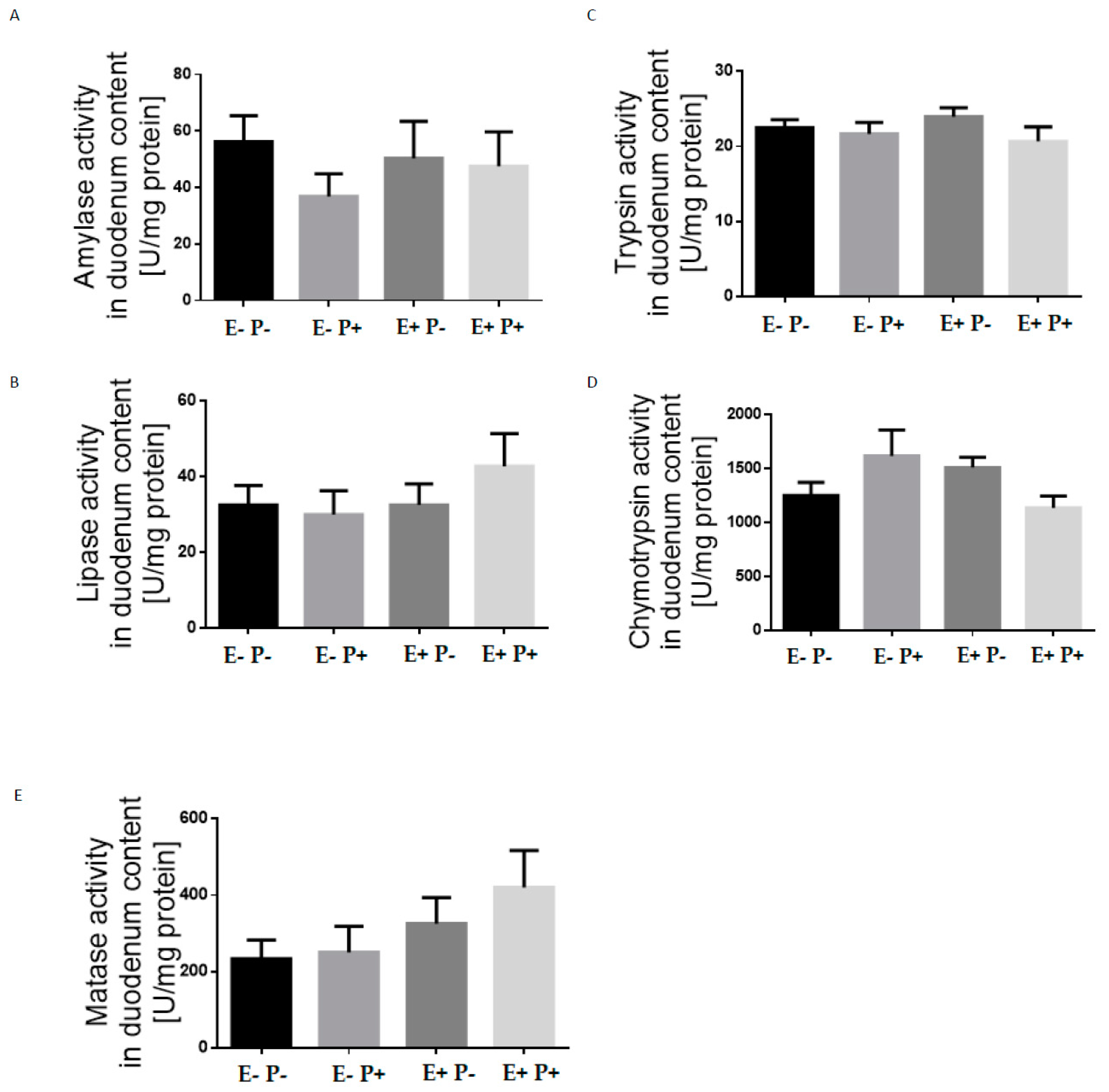

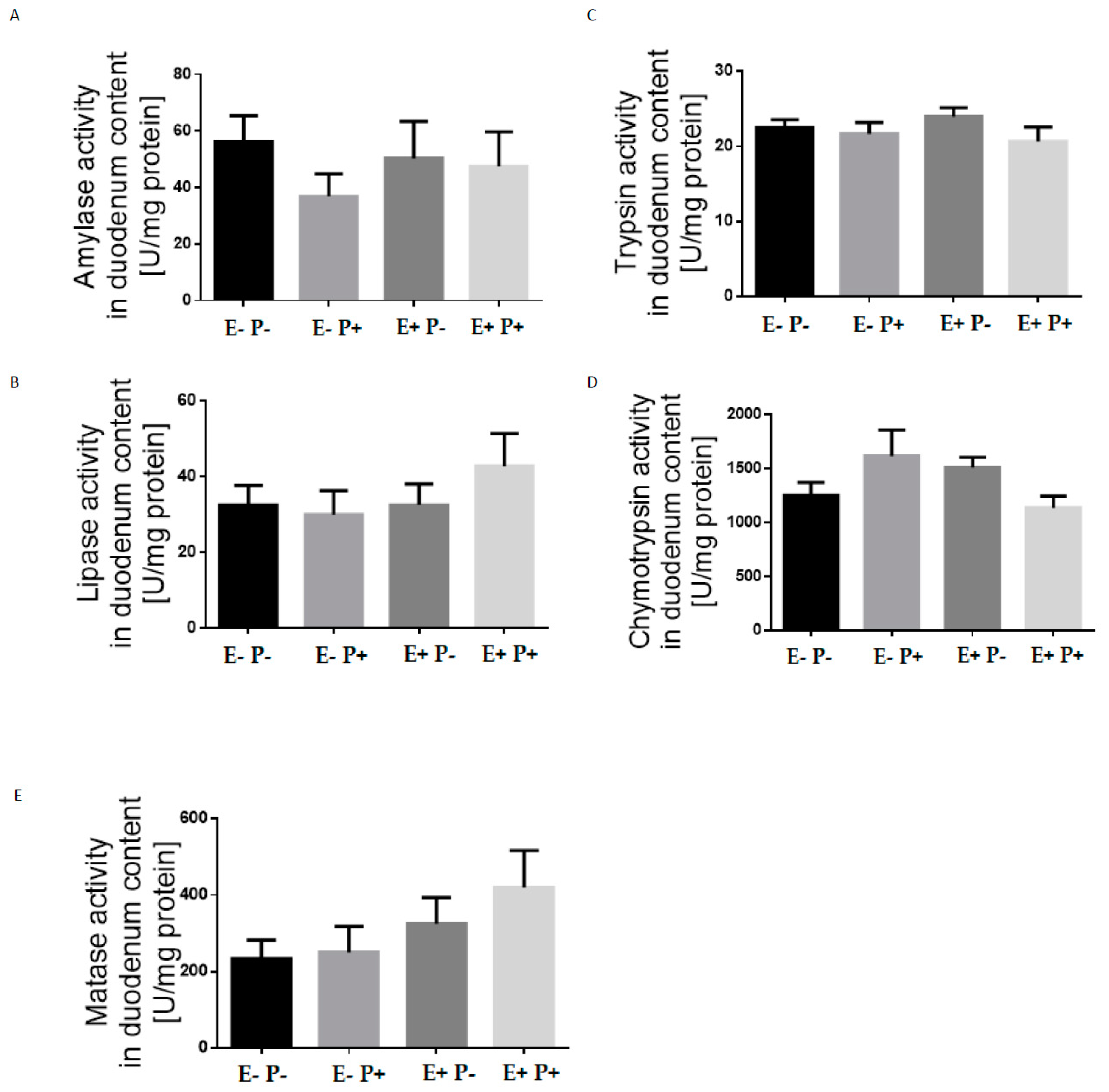{kind=link}
| Item (% DM) | Faba Bean | p | |
|---|---|---|---|
| Extrusion − | Extrusion + | ||
| Crude protein | 34.7 ± 1.6 | 32.7 ± 1.9 | NS |
| ADF | 11.8 ± 1.0 | 11.4 ± 0.9 | NS |
| NDF | 19.8 ± 1.4 | 12.9 ± 0.9 | ≤0.001 |
| Starch | 41.7 ± 4.2 | 41.1 ± 3.3 | NS |
| RS | 18.3 ± 0.6 | 0.9 ± 0.1 | ≤0.001 |
| Antinutritional factors | |||
| Oligosaccharides | 3.03 ± 0.2 | 3.01 ± 0.1 | NS |
| Tannins | 0.006 ± 0.001 | 0.006 ± 0.001 | NS |
| TIA | 0.060 ± 0.001 | 0.020 ± 0.001 | ≤0.001 |
| Phytic-P | 0.50 ± 0.03 | 0.19 ± 0.01 | 0.001 |
| Total NSP | 15.36 ± 0.93 | 15.21 ± 0.78 | NS |
| Essential amino acids (g/16g N) | |||
| Lysine | 5.3 ± 0.3 | 5.3 ± 0.4 | NS |
| Threonine | 3.1 ± 0.1 | 3.0 ± 0.2 | NS |
| Methionine | 0.5 ± 0.0 | 0.5 ± 0.1 | NS |
| Cysteine | 1.0 ± 0.0 | 1.0 ± 0.1 | NS |
| Isoleucine | 3.5 ± 0.4 | 3.5 ± 0.2 | NS |
| Valine | 4.0 ± 0.2 | 4.0 ± 0.1 | NS |
| Leucine | 6.5 ± 0.3 | 5.9 ± 0.4 | NS |
| Phenylalanine | 3.7 ± 0.2 | 3.7 ± 0.1 | NS |
| Histidine | 2.4 ± 0.1 | 2.4 ± 0.1 | NS |
| Arginine | 8.3 ± 0.5 | 8.2 ± 0.2 | NS |
| Glycine | 3.7 ± 0.3 | 3.6 ± 0.3 | NS |
| Non-essential amino acids (g/16g N) | |||
| Tyrosine | 2.7 ± 0.1 | 2.6 ± 0.2 | NS |
| Alanine | 3.5 ± 0.2 | 3.5 ± 0.1 | NS |
| Aspartic acid | 9.4 ± 0.5 | 9.4 ± 0.5 | NS |
| Glutamic acid | 16.9 ± 0.9 | 16.8 ± 1.0 | NS |
| Serine | 4.2 ± 0.3 | 4.1 ± 0.4 | NS |
| Proline | 3.2 ± 0.1 | 3.2 ± 0.1 | NS |
| Treatments | ||||
|---|---|---|---|---|
| Extrusion | − | − | + | + |
| Enzyme | − | + | − | + |
| Ingredients (%) | ||||
| Soybean meal | 16.300 | 12.600 | 16.300 | 12.600 |
| Faba bean | 9.100 | 9.100 | 9.100 | 9.100 |
| Wheat | 40.000 | 40.000 | 40.000 | 40.000 |
| Triticale | 28.065 | 32.715 | 28.065 | 32.715 |
| Premix 0.5% * | 0.500 | 0.500 | 0.500 | 0.500 |
| Phosphate 1-Ca | 1.150 | 1.150 | 1.150 | 1.150 |
| Limestone | 0.850 | 0.850 | 0.850 | 0.850 |
| NaCl | 0.350 | 0.350 | 0.350 | 0.350 |
| Rapeseed oil | 2.500 | 1.500 | 2.500 | 1.500 |
| L-Lysine HCl | 0.440 | 0.440 | 0.440 | 0.440 |
| DL-Methionine | 0.150 | 0.150 | 0.150 | 0.150 |
| DL-Tryptophan | 0.025 | 0.025 | 0.025 | 0.025 |
| L-Threonine | 0.270 | 0.270 | 0.270 | 0.270 |
| TiO2 | 0.300 | 0.300 | 0.300 | 0.300 |
| Protease | − | 0.050 | − | 0.050 |
| Feed nutritional value (calculated) in dry matter | ||||
| Dry matter (%) | 88.60 | 88.92 | 89.10 | 89.49 |
| ME (MJ/kg) | 13.72 | 13.72 | 13.72 | 13.72 |
| CP% | 17.90 | 16.88 | 17.90 | 16.88 |
| Dig. CP% | 14.82 | 14.82 | 14.81 | 14.82 |
| Dig. Lys% | 1.150 | 1.150 | 1.150 | 1.150 |
| Dig. Met% | 0.363 | 0.363 | 0.363 | 0.363 |
| Dig. Trp% | 0.259 | 0.259 | 0.259 | 0.259 |
| Dig. Thr% | 0.722 | 0.722 | 0.722 | 0.722 |
| Ca% | 0.759 | 0.759 | 0.759 | 0.759 |
| P% | 0.654 | 0.654 | 0.654 | 0.654 |
| Dig. P% | 0.433 | 0.433 | 0.433 | 0.433 |
| Na% | 0.152 | 0.152 | 0.152 | 0.152 |
| Extrusion | − | − | + | + | p | |||
|---|---|---|---|---|---|---|---|---|
| Protease | − | + | − | + | p | Extrusion | Protease | Interaction |
| IBW 0d (kg) | 9.36 | 9.43 | 9.29 | 9.64 | 0.960 | 0.885 | 0.666 | 0.773 |
| BW 14d (kg) | 14.14 | 13.86 | 14.00 | 14.50 | 0.908 | 0.703 | 0.870 | 0.551 |
| FBW 28d (kg) | 23.43 | 22.79 | 24.57 | 24.71 | 0.595 | 0.196 | 0.830 | 0.737 |
| DWG (kg/day) | ||||||||
| 0–14 days | 0.34 | 0.32 | 0.34 | 0.35 | 0.804 | 0.593 | 0.741 | 0.453 |
| 15–28 days | 0.66 | 0.64 | 0.75 | 0.73 | 0.325 | 0.078 | 0.613 | 0.998 |
| 0–28 days | 0.50 | 0.48 | 0.55 | 0.54 | 0.325 | 0.084 | 0.580 | 0.766 |
| DFI (kg) | ||||||||
| 0–14 days | 0.63 | 0.65 | 0.68 | 0.69 | 0.349 | 0.109 | 0.427 | 0.897 |
| 15–28 days | 1.05 | 1.12 | 1.22 | 1.27 | 0.203 | 0.048 | 0.420 | 0.892 |
| 0–28 days | 0.84 | 0.89 | 0.95 | 0.98 | 0.220 | 0.055 | 0.429 | 0.894 |
| FCR (kg/kg) | ||||||||
| 0–14 days | 1.85 | 2.09 | 2.08 | 2.06 | 0.454 | 0.419 | 0.368 | 0.285 |
| 15–28 days | 1.58 | 1.78 | 1.63 | 1.76 | 0.223 | 0.863 | 0.045 | 0.651 |
| 0–28 days | 1.67 b | 1.87 a | 1.75 a,b | 1.83 a | 0.035 | 0.735 | 0.008 | 0.221 |
| Extrusion | − | − | + | + | p | |||
|---|---|---|---|---|---|---|---|---|
| Protease | − | + | − | + | p | Extrusion | Protease | Interaction |
| Crude protein (%) | 73.64 | 72.74 | 74.20 | 74.81 | 0.797 | 0.391 | 0.784 | 0.784 |
| Dry matter (%) | 76.43 | 77.15 | 77.36 | 77.45 | 0.942 | 0.636 | 0.623 | 0.623 |
| Extrusion | − | − | + | + | p | |||
|---|---|---|---|---|---|---|---|---|
| Protease | − | + | − | + | p | Extrusion | Protease | Interaction |
| Liver weight (g) | 735.33 | 748.00 | 757.67 | 831.00 | 0.439 | 0.248 | 0.343 | 0.501 |
| Liver/BWG ratio (kg/kg) | 0.032 | 0.031 | 0.032 | 0.030 | 0.828 | 0.756 | 0.417 | 0.756 |
| pH | ||||||||
| Ileum | 5.90 | 5.56 | 5.79 | 5.56 | 0.647 | 0.796 | 0.225 | 0.958 |
| Cecum | 5.16 b | 5.13 b | 5.42 a | 5.53 a | ≤0.001 | ≤0.001 | 0.509 | 0.430 |
| Ammonia (µmol/g digesta) | ||||||||
| Ileum | 13.81 | 15.71 | 11.96 | 11.18 | 0.443 | 0.144 | 0.791 | 0.529 |
| Cecum | 15.80 | 16.42 | 18.92 | 18.13 | 0.059 | 0.209 | 0.897 | 0.622 |
| Dry matter (%) | ||||||||
| Ileum | 12.28 a | 11.53 a,b | 10.66 b,c | 9.97 c | 0.02 | 0.004 | 0.161 | 0.958 |
| Cecum | 9.14 | 10.73 | 10.08 | 10.30 | 0.605 | 0.763 | 0.298 | 0.430 |
| Viscosity (cP) | ||||||||
| Ileum | 1.11 | 1.10 | 1.09 | 1.19 | 0.758 | 0.702 | 0.553 | 0.423 |
| Extrusion | − | − | + | + | p | |||
|---|---|---|---|---|---|---|---|---|
| Protease | − | + | − | + | p | Extrusion | Protease | Interaction |
| Acetic acid | 55.17 b | 57.88 a,b | 58.65 a | 59.68 a | 0.033 | 0.016 | 0.085 | 0.475 |
| Propionic acid | 27.17 a | 23.55 b | 23.15 b | 21.00 b | ≤0001 | ≤0.0001 | 0.003 | 0.376 |
| Isobutyric acid | 2.75 | 2.70 | 1.90 | 1.80 | 0.396 | 0.094 | 0.882 | 0.960 |
| Butyric acid | 10.50 a | 8.70 b | 6.18 c | 4.73 d | ≤0.0001 | ≤0.0001 | ≤0.0001 | 0.643 |
| Isovaleric acid | 1.75 a | 0.70 b | 0.62 b | 0.35 b | ≤0.0001 | ≤0.0001 | ≤0.0001 | 0.072 |
| Valeric acid | 2.82 a | 1.55 b | 1.08 b | 3.37 a | ≤0.0001 | 0.901 | 0.140 | ≤0.0001 |
| Acetoacetic acid | 0.00 c | 5.20 b | 8.58 a | 9.35 a | ≤0.0001 | ≤0.0001 | 0.011 | 0.060 |
| Extrusion | − | − | + | + | p | |||
|---|---|---|---|---|---|---|---|---|
| Protease | − | + | − | + | p | Extrusion | Protease | Interaction |
| Villus length (µm) | 413.31 | 449.51 | 393.18 | 409.08 | 0.707 | 0.393 | 0.462 | 0.773 |
| Villus width (µm) | 104.99 | 108.73 | 104.24 | 105.04 | 0.911 | 0.649 | 0.643 | 0.764 |
| Crypt depth (µm) | 160.47 | 163.43 | 168.16 | 166.07 | 0.901 | 0.506 | 0.956 | 0.744 |
| Villus/crypt ratio | 2.56 | 2.77 | 2.35 | 2.49 | 0.562 | 0.250 | 0.420 | 0.875 |
| Extrusion | − | − | + | + | p | |||
|---|---|---|---|---|---|---|---|---|
| Protease | − | + | − | + | p | Extrusion | Protease | Interaction |
| Glucose (mg/dL) | 61.10 b | 59.19 b | 69.86 a | 63.80 a,b | 0.030 | 0.013 | 0.122 | 0.424 |
| TG (mg/dL) | 76.94 | 80.71 | 85.39 | 73.52 | 0.270 | 0.886 | 0.358 | 0.082 |
| Total cholesterol (mg/dL) | 91.13 | 84.90 | 92.33 | 87.95 | 0.658 | 0.644 | 0.253 | 0.841 |
| Albumin (g/dL) | 2.96 | 2.89 | 3.11 | 2.92 | 0.329 | 0.315 | 0.156 | 0.524 |
| Total protein (g/dL) | 4.75 | 4.69 | 4.69 | 4.54 | 0.335 | 0.411 | 0.400 | 0.687 |
| ALT (IU/L) | 5.17 c | 7.21 b | 8.72 a,b | 9.31 a | ≤0.001 | 0.001 | 0.058 | 0.284 |
| AST (IU/L) | 28.88 | 27.84 | 30.5 | 33.59 | 0.515 | 0.205 | 0.719 | 0.474 |
| ALP (IU/L) | 88.17 | 91.72 | 91.58 | 102.38 | 0.591 | 0.367 | 0.358 | 0.640 |
| GGT (IU/L) | 14.37 a | 9.21 a,b | 4.79 c | 10.32 a,b | 0.043 | 0.069 | 0.935 | 0.024 |
| LDH (IU/L) | 283.21 | 286.67 | 283.21 | 306.86 | 0.711 | 0.551 | 0.424 | 0.551 |
| Creatinine (mg/dL) | 0.69 | 0.66 | 0.66 | 0.62 | 0.890 | 0.581 | 0.581 | 0.999 |
| BUN (mg/dL) | 16.26 | 15.16 | 17.58 | 15.00 | 0.370 | 0.616 | 0.119 | 0.524 |
| Urea (mg/dL) | 34.8 | 32.44 | 37.62 | 32.01 | 0.370 | 0.616 | 0.119 | 0.524 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaworska-Zakrzewska, A.; Kasprowicz-Potocka, M.; Ciołek, K.; Pruszyńska-Oszmałek, E.; Stuper-Szablewska, K.; Rutkowski, A. The Effects of Protease Supplementation and Faba Bean Extrusion on Growth, Gastrointestinal Tract Physiology and Selected Blood Indices of Weaned Pigs. Animals 2022, 12, 563. https://doi.org/10.3390/ani12050563
Zaworska-Zakrzewska A, Kasprowicz-Potocka M, Ciołek K, Pruszyńska-Oszmałek E, Stuper-Szablewska K, Rutkowski A. The Effects of Protease Supplementation and Faba Bean Extrusion on Growth, Gastrointestinal Tract Physiology and Selected Blood Indices of Weaned Pigs. Animals. 2022; 12(5):563. https://doi.org/10.3390/ani12050563
Chicago/Turabian StyleZaworska-Zakrzewska, Anita, Małgorzata Kasprowicz-Potocka, Klaudia Ciołek, Ewa Pruszyńska-Oszmałek, Kinga Stuper-Szablewska, and Andrzej Rutkowski. 2022. "The Effects of Protease Supplementation and Faba Bean Extrusion on Growth, Gastrointestinal Tract Physiology and Selected Blood Indices of Weaned Pigs" Animals 12, no. 5: 563. https://doi.org/10.3390/ani12050563
APA StyleZaworska-Zakrzewska, A., Kasprowicz-Potocka, M., Ciołek, K., Pruszyńska-Oszmałek, E., Stuper-Szablewska, K., & Rutkowski, A. (2022). The Effects of Protease Supplementation and Faba Bean Extrusion on Growth, Gastrointestinal Tract Physiology and Selected Blood Indices of Weaned Pigs. Animals, 12(5), 563. https://doi.org/10.3390/ani12050563