
Early Pregnancy Modulates Expression of the Nod-like Receptor Family in Lymph Nodes of Ewes


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. RNA Extraction and RT-qPCR Assay
2.3. Western Blot Analysis
2.4. Immunohistochemistry Analysis
2.5. Statistical Analysis
3. Results
3.1. Expression of NOD1, NOD2, CIITA, NAIP, NLRP1, NLRP3 and NLRP7 mRNA in Lymph Nodes
3.2. Expression of NOD1, NOD2, CIITA, NAIP, NLRP1, NLRP3 and NLRP7 Proteins in Lymph Nodes
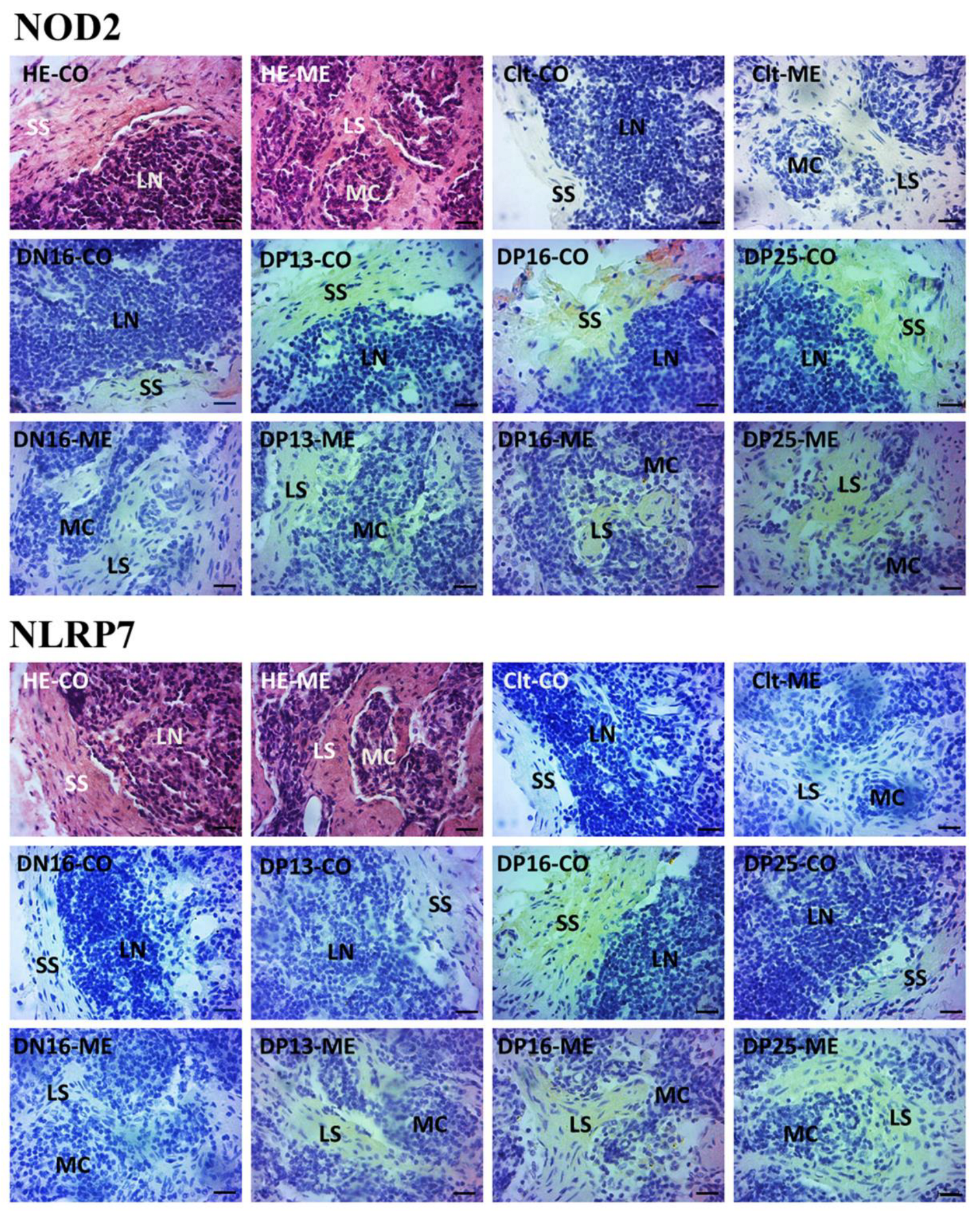
3.3. Immunohistochemistry for NOD2 and NLRP7 Proteins in Maternal Lymph Nodes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, L.; Cao, S.Q.; Lin, Z.M.; He, S.J.; Zuo, J.P. NOD-like receptors in autoimmune diseases. Acta Pharmacol. Sin. 2021, 42, 1742–1756. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, T.; Hovingh, E.S.; Foerster, E.G.; Abdel-Nour, M.; Philpott, D.J.; Girardin, S.E. NOD1 and NOD2 in inflammation, immunity and disease. Arch. Biochem. Biophys. 2019, 670, 69–81. [Google Scholar] [CrossRef]
- Cho, S.X.; Vijayan, S.; Yoo, J.S.; Watanabe, T.; Ouda, R.; An, N.; Kobayashi, K.S. MHC class I transactivator NLRC5 in host immunity, cancer and beyond. Immunology 2021, 162, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.C.; Sandall, C.F.; Carlson, D.A.; Ulke-Lemée, A.; Platnich, J.M.; Hughes, P.F.; Muruve, D.A.; Haystead, T.A.J.; MacDonald, J.A. Application of immobilized ATP to the study of NLRP inflammasomes. Arch. Biochem. Biophys. 2019, 670, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Morón-Calvente, V.; Romero-Pinedo, S.; Toribio-Castelló, S.; Plaza-Díaz, J.; Abadía-Molina, A.C.; Rojas-Barros, D.I.; Beug, S.T.; LaCasse, E.C.; MacKenzie, A.; Korneluk, R.; et al. Inhibitor of apoptosis proteins, NAIP, cIAP1 and cIAP2 expression during macrophage differentiation and M1/M2 polarization. PLoS ONE 2018, 13, e0193643. [Google Scholar] [CrossRef]
- Lim, R.; Lappas, M. NOD-like receptor pyrin domain-containing-3 (NLRP3) regulates inflammation-induced pro-labor mediators in human myometrial cells. Am. J. Reprod. Immunol. 2018, 79, e12825. [Google Scholar] [CrossRef]
- Triantafilou, K. Enigmatic inflammasomes—Sequel (Part 2). Immunology 2021, 163, 345–347. [Google Scholar] [CrossRef]
- Wang, W.; Sung, N.; Gilman-Sachs, A.; Kwak-Kim, J. T helper (Th) cell profiles in pregnancy and recurrent pregnancy losses: Th1/Th2/Th9/Th17/Th22/Tfh cells. Front. Immunol. 2020, 11, 2025. [Google Scholar] [CrossRef]
- Skarzynski, D.J.; Bazer, F.W.; Maldonado-Estrada, J.G. Editorial: Veterinary reproductive immunology. Front. Vet. Sci. 2022, 8, 823169. [Google Scholar] [CrossRef]
- Rocha, C.C.; da Silveira, J.C.; Forde, N.; Binelli, M.; Pugliesi, G. Conceptus-modulated innate immune function during early pregnancy in ruminants: A review. Anim. Reprod. 2021, 18, e20200048. [Google Scholar] [CrossRef]
- Ott, T.L. Immunological detection of pregnancy: Evidence for systemic immune modulation during early pregnancy in ruminants. Theriogenology 2020, 150, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, B.; Yan, X.; Zhang, L.; Gao, F.; Liu, Z. Expression of ISG15 in bone marrow during early pregnancy in ewes. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 767–772. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Mi, H.; Yan, J.K.; Yan, X.X.; Yang, L. Pregnancy-associated changes in expression of progesterone receptor and progesterone-induced blocking factor genes in bone marrow of ewes. Anim. Reprod. Sci. 2017, 186, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xue, J.; Wang, Q.; Lv, W.; Mi, H.; Liu, Y.; Yang, L. Changes in expression of ISG15, progesterone receptor and progesterone-induced blocking factor in ovine thymus during early pregnancy. Theriogenology 2018, 121, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, Z.; Wang, Y.; Li, N.; Cao, N.; Yang, L. Changes in expression of interferon-stimulated genes and ubiquitin activating enzyme E1-like in ovine thymus during early pregnancy. Anim. Reprod. 2020, 17, e20190134. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.; Lv, W.; Wang, P.; Wang, B.; Xue, J.; Zhang, L. Expression of interferon-stimulated gene 15-kDa protein, cyclooxygenase (COX) 1, COX-2, aldo-keto reductase family 1, member B1, and prostaglandin E synthase in the spleen during early pregnancy in sheep. Anim. Sci. J. 2018, 89, 1540–1548. [Google Scholar] [CrossRef]
- Yang, L.; Guo, R.; Yao, X.; Yan, J.; Bai, Y.; Zhang, L. Expression of progesterone receptor and progesterone-induced blocking factor in the spleen during early pregnancy in ewes. Livest. Sci. 2018, 209, 14–19. [Google Scholar] [CrossRef]
- Wang, Y.; Han, X.; Zhang, L.; Cao, N.; Cao, L.; Yang, L. Early pregnancy induces expression of STAT1, OAS1 and CXCL10 in ovine spleen. Animals 2019, 9, 882. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, L.; Yang, F.; Han, X.; Wang, Y.; Cao, N.; Yang, L. Relative abundance of interferon-stimulated genes STAT1, OAS1, CXCL10 and MX1 in ovine lymph nodes during early pregnancy. Anim. Reprod. Sci. 2020, 214, 106285. [Google Scholar] [CrossRef]
- Yang, L.; Zang, S.; Bai, Y.; Yao, X.; Zhang, L. Effect of early pregnancy on the expression of progesterone receptor and progesterone-induced blocking factor in ovine lymph node. Theriogenology 2017, 93, 78–83. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Q.; Liu, Y.; Zhang, L.; Lv, W.; Liu, B. Expression profiles of interferon-stimulated gene 15 and prostaglandin synthases in the ovine lymph nodes during early pregnancy. Mol. Reprod. Dev. 2019, 86, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Leal, J.M.; Huang, J.Y.; Kohli, K.; Stoltzfus, C.; Lyons-Cohen, M.R.; Olin, B.E.; Gale, M., Jr.; Gerner, M.Y. Innate cell microenvironments in lymph nodes shape the generation of T cell responses during type I inflammation. Sci. Immunol. 2021, 6, eabb9435. [Google Scholar] [CrossRef]
- Quirke, L.D.; Maclean, P.H.; Haack, N.A.; Edwards, S.J.; Heiser, A.; Juengel, J.L. Characterization of local and peripheral immune system in pregnant and nonpregnant ewes. J. Anim. Sci. 2021, 99, skab208. [Google Scholar] [CrossRef] [PubMed]
- Klossner, R.; Groessl, M.; Schumacher, N.; Fux, M.; Escher, G.; Verouti, S.; Jamin, H.; Vogt, B.; Mohaupt, M.G.; Gennari-Moser, C. Steroid hormone bioavailability is controlled by the lymphatic system. Sci. Rep. 2021, 11, 9666. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, P.; Mi, H.; Lv, W.; Liu, B.; Du, J.; Zhang, L. Comparison of Th1 and Th2 cytokines production in ovine lymph nodes during early pregnancy. Theriogenology 2019, 123, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Zhang, L.; Zhao, Z.; Li, N.; Wang, B.; Yang, L. Expression of melatonin receptors and CD4 in the ovine thymus, lymph node, spleen and liver during early pregnancy. Immunology 2020, 160, 52–63. [Google Scholar] [CrossRef]
- Cao, N.; Cao, L.; Gao, M.; Wang, H.; Zhang, L.; Yang, L. Changes in mRNA and protein levels of gonadotropin releasing hormone and receptor in ovine thymus, lymph node, spleen, and liver during early pregnancy. Domest. Anim. Endocrinol. 2021, 76, 106607. [Google Scholar] [CrossRef]
- Feng, P.; Wu, J.; Ren, Y.; Zhang, L.; Cao, J.; Yang, L. Early pregnancy regulates the expression of prolactin and its receptor in the thymus, the liver, the spleen and lymph nodes in sheep. Domest. Anim. Endocrinol. 2022, 81, 106731. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Q.; Zhang, L.; Feng, P.; Gao, M.; Zhao, Z.; Yang, L. Toll-like receptor signaling is changed in ovine lymph node during early pregnancy. Anim. Sci. J. 2021, 92, e13541. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, L.; Feng, P.; Han, X.; Yang, L. Complement regulation in ovine lymph nodes during early pregnancy. Exp. Ther. Med. 2022, 23, 166. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, T.; Yang, Z.; Cai, C.; Hao, S.; Yang, L. Expression of nuclear factor kappa B in ovine maternal inguinal lymph nodes during early pregnancy. BMC Vet. Res. 2022, 18, 266. [Google Scholar] [CrossRef] [PubMed]
- Mazrouei, S.; Ziaei, A.; Tanhaee, A.P.; Keyhanian, K.; Esmaeili, M.; Baradaran, A.; Salehi, M. Apoptosis inhibition or inflammation: The role of NAIP protein expression in Hodgkin and non-Hodgkin lymphomas compared to non-neoplastic lymph node. J. Inflamm. 2012, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, V.; González-Ramos, S.; Avendaño-Ortiz, J.; Martín-Sanz, P.; Delgado, C.; Castrillo, A.; Boscá, L. NOD1 splenic activation confers ferroptosis protection and reduces macrophage recruitment under pro-atherogenic conditions. Biomed. Pharmacother. 2022, 148, 112769. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, M.; Nie, X.; Zhang, Y.; Chen, Y.; Zhu, L.; Chen, X.; Chen, L.; Chen, H.; Zhang, J. NOD1 and NOD2 control the invasiveness of trophoblast cells via the MAPK/p38 signaling pathway in human first-trimester pregnancy. Placenta 2015, 36, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, I.; Mulla, M.J.; Myrtolli, K.; Sfakianaki, A.K.; Norwitz, E.R.; Tadesse, S.; Guller, S.; Abrahams, V.M. Nod1 activation by bacterial iE-DAP induces maternal-fetal inflammation and preterm labor. J. Immunol. 2011, 187, 980–986. [Google Scholar] [CrossRef]
- Inoue, H.; Nishio, H.; Takada, H.; Sakai, Y.; Nanishi, E.; Ochiai, M.; Onimaru, M.; Chen, S.J.; Matsui, T.; Hara, T. Activation of Nod1 Signaling Induces Fetal Growth Restriction and Death through Fetal and Maternal Vasculopathy. J. Immunol. 2016, 196, 2779–2787. [Google Scholar] [CrossRef]
- Gurugubelli Krishna, R.; Vishnu Bhat, B. Molecular mechanisms of intrauterine growth restriction. J. Matern. Fetal. Neonatal. Med. 2018, 31, 2634–2640. [Google Scholar] [CrossRef]
- Rakner, J.J.; Silva, G.B.; Mundal, S.B.; Thaning, A.J.; Elschot, M.; Ostrop, J.; Thomsen, L.C.V.; Bjørge, L.; Gierman, L.M.; Iversen, A.C. Decidual and placental NOD1 is associated with inflammation in normal and preeclamptic pregnancies. Placenta 2021, 105, 23–31. [Google Scholar] [CrossRef]
- Alipoor, S.D.; Mirsaeidi, M. Inborn Errors in the LRR Domain of Nod2 and Their Potential Consequences on the Function of the Receptor. Cells 2021, 10, 2031. [Google Scholar] [CrossRef] [PubMed]
- Costello, M.J.; Joyce, S.K.; Abrahams, V.M. NOD protein expression and function in first trimester trophoblast cells. Am. J. Reprod. Immunol. 2007, 57, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, C.; Fu, S.; Chen, X.; Zhang, S.; Li, Y.; Du, M.; Zhang, J. Different expression of NOD2 in decidual stromal cells between normal and unexplained recurrent spontaneous abortion women during first trimester gestation. Int. J. Clin. Exp. Pathol. 2014, 7, 8784–8790. [Google Scholar]
- van Rijn, B.B.; Franx, A.; Steegers, E.A.; de Groot, C.J.; Bertina, R.M.; Pasterkamp, G.; Voorbij, H.A.; Bruinse, H.W.; Roest, M. Maternal TLR4 and NOD2 gene variants, pro-inflammatory phenotype and susceptibility to early-onset preeclampsia and HELLP syndrome. PLoS ONE 2008, 3, e1865. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Gourley, T.S.; Sisk, T.J. Function and regulation of class II transactivator in the immune system. Immunol. Res. 2002, 25, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Yoo, I.; Kim, D.; Han, J.; Lee, S.; Hong, M.; Jeon, B.Y.; Kim, J.M.; Ka, H. Transcriptomic analysis of interferon-γ-regulated genes in endometrial explants and their possible role in regulating maternal endometrial immunity during the implantation period in pigs, a true epitheliochorial placentation species. Theriogenology 2020, 155, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Heuberger, C.; Pott, J.; Maloy, K.J. Why do intestinal epithelial cells express MHC class II? Immunology 2021, 162, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, H.; Akizawa, H.; Sada, A.; Kishi, Y.; Yamanaka, K.; Takuma, T.; Sasaki, K.; Yamauchi, N.; Yanagawa, Y.; Nagano, M.; et al. Comparing spatial expression dynamics of bovine blastocyst under three different procedures: In-vivo, in-vitro derived, and somatic cell nuclear transfer embryos. Jpn. J. Vet. Res. 2015, 63, 159–171. [Google Scholar] [CrossRef]
- Bauer, R.; Rauch, I. The NAIP/NLRC4 inflammasome in infection and pathology. Mol. Aspects Med. 2020, 76, 100863. [Google Scholar] [CrossRef]
- Ka, H.; Hunt, J.S. Temporal and spatial patterns of expression of inhibitors of apoptosis in human placentas. Am. J. Pathol. 2003, 163, 413–422. [Google Scholar] [CrossRef]
- Zhao, L.; Liang, X.; Ma, Y.; Li, J.; Liao, S.; Chen, J.; Wang, C. AK002210 promotes the proliferation, migration and invasion of trophoblast cell through regulating miR-590/NAIP signal axis. Arch. Biochem. Biophys. 2020, 688, 108366. [Google Scholar] [CrossRef]
- Kummer, J.A.; Broekhuizen, R.; Everett, H.; Agostini, L.; Kuijk, L.; Martinon, F.; van Bruggen, R.; Tschopp, J. Inflammasome components NALP 1 and 3 show distinct but separate expression profiles in human tissues suggesting a site-specific role in the inflammatory response. J. Histochem. Cytochem. 2007, 55, 443–452. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Sandstrom, A.; Vance, R.E. The NLRP1 inflammasome: New mechanistic insights and unresolved mysteries. Curr. Opin. Immunol. 2019, 60, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Meihe, L.; Shan, G.; Minchao, K.; Xiaoling, W.; Peng, A.; Xili, W.; Jin, Z.; Huimin, D. The ferroptosis-NLRP1 inflammasome: The vicious cycle of an adverse pregnancy. Front. Cell. Dev. Biol. 2021, 9, 707959. [Google Scholar] [CrossRef] [PubMed]
- Nunes, P.R.; Romao-Veiga, M.; Matias, M.L.; Ribeiro, V.R.; de Oliveira, L.; Peracoli, J.C.; Peracoli, M.T.S. Vitamin D decreases expression of NLRP1 and NLRP3 inflammasomes in placental explants from women with preeclampsia cultured with hydrogen peroxide. Hum. Immunol. 2022, 83, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Yan, Y.; Jiang, X.; Mai, J.; Chen, N.C.; Wang, H.; Yang, X.F. Inflammasomes are differentially expressed in cardiovascular and other tissues. Int. J. Immunopathol. Pharmacol. 2009, 22, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Alatshan, A.; Benkő, S. Nuclear Receptors as Multiple Regulators of NLRP3 Inflammasome Function. Front. Immunol. 2021, 12, 630569. [Google Scholar] [CrossRef]
- Weel, C.I.; Romão-Veiga, M.; Matias, M.L.; Fioratti, E.G.; Peraçoli, J.C.; Borges, V.T.; Araujo, J.P., Jr.; Peraçoli, M.T. Increased expression of NLRP3 inflammasome in placentas from pregnant women with severe preeclampsia. J. Reprod. Immunol. 2017, 123, 40–47. [Google Scholar] [CrossRef]
- Shirasuna, K.; Karasawa, T.; Takahashi, M. Role of the NLRP3 Inflammasome in Preeclampsia. Front. Endocrinol. 2020, 11, 80. [Google Scholar] [CrossRef]
- Cheng, X.; Zhang, Y.; Ma, J.; Wang, S.; Ma, R.; Ge, X.; Zhao, W.; Xue, T.; Chen, L.; Yao, B. NLRP3 promotes endometrial receptivity by inducing epithelial-mesenchymal transition of the endometrial epithelium. Mol. Hum. Reprod. 2021, 27, gaab056. [Google Scholar] [CrossRef]
- Soczewski, E.; Grasso, E.; Gallino, L.; Hauk, V.; Fernández, L.; Gori, S.; Paparini, D.; Perez Leirós, C.; Ramhorst, R. Immunoregulation of the decidualization program: Focus on the endoplasmic reticulum stress. Reproduction 2020, 159, R203–R211. [Google Scholar] [CrossRef]
- Tsai, P.Y.; Chen, K.R.; Li, Y.C.; Kuo, P.L. NLRP7 is involved in the differentiation of the decidual macrophages. Int. J. Mol. Sci. 2019, 20, 5994. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Yu, P.H.; Li, Y.C.; Kuo, P.L. NLRP7 contributes to in vitro decidualization of endometrial stromal cells. Reprod. Biol. Endocrinol. 2017, 15, 66. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tian, X.; Lv, D.; Zhang, L.; Zhang, Z.; Wang, J.; Yang, M.; Tao, J.; Ma, T.; Wu, H.; et al. NLRP7 is expressed in the ovine ovary and associated with in vitro pre-implantation embryo development. Reproduction 2019, 158, 415–427. [Google Scholar] [CrossRef]
- Mahadevan, S.; Wen, S.; Wan, Y.W.; Peng, H.H.; Otta, S.; Liu, Z.; Iacovino, M.; Mahen, E.M.; Kyba, M.; Sadikovic, B.; et al. NLRP7 affects trophoblast lineage differentiation, binds to overexpressed YY1 and alters CpG methylation. Hum. Mol. Genet. 2014, 23, 706–716. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence | Size (bp) | Accession Numbers |
|---|---|---|---|---|
| NOD1 | Forward | CCTTGGCTGTCAGAGATTGGCTTC | 94 | XM_042248630.1 |
| Reverse | GCTTCTGGCTGTATCTGCTCACTG | |||
| NOD2 | Forward | TGCCATCCTCGCTCAGACATCTC | 117 | XM_042231601.1 |
| Reverse | CAGCCACACTGCCCTCTTTGC | |||
| CIITA | Forward | GCACCTCCTTCCAGTTCCTTGTTG | 119 | XM_042239890.1 |
| Reverse | CCTGTCCCAGTCCCTGAGATCG | |||
| NAIP | Forward | TTGTCCAGCAGTGTCAGCATCTTC | 82 | XM_012096791.3 |
| Reverse | ATTTCCACCACGCTGTCATCATCC | |||
| NLRP1 | Forward | AAGGAGGTGACCGAGATGCTGAG | 143 | XM_012185551.4 |
| Reverse | TGCCGCTTGAGTGAGGATGTATTG | |||
| NLRP3 | Forward | CTCTGGTTGGTCAGTTGCTGTCTC | 81 | XM_042250402.1 |
| Reverse | GGTCAGGGAATGGTTGGTGCTTAG | |||
| NLRP7 | Forward | GCCTGCTACTCGTTCATCCATCTC | 90 | XM_004015893.5 |
| Reverse | CCCTTCCTCCTCCTGCTCTTCC | |||
| GAPDH | Forward | GGGTCATCATCTCTGCACCT | 176 | NM_001190390.1 |
| Reverse | GGTCATAAGTCCCTCCACGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Li, Y.; Cao, J.; Fang, H.; Zhang, L.; Yang, L. Early Pregnancy Modulates Expression of the Nod-like Receptor Family in Lymph Nodes of Ewes. Animals 2022, 12, 3285. https://doi.org/10.3390/ani12233285
Zhao Z, Li Y, Cao J, Fang H, Zhang L, Yang L. Early Pregnancy Modulates Expression of the Nod-like Receptor Family in Lymph Nodes of Ewes. Animals. 2022; 12(23):3285. https://doi.org/10.3390/ani12233285
Chicago/Turabian StyleZhao, Zhenyang, Yuanjing Li, Jianhua Cao, Hongxu Fang, Leying Zhang, and Ling Yang. 2022. "Early Pregnancy Modulates Expression of the Nod-like Receptor Family in Lymph Nodes of Ewes" Animals 12, no. 23: 3285. https://doi.org/10.3390/ani12233285
APA StyleZhao, Z., Li, Y., Cao, J., Fang, H., Zhang, L., & Yang, L. (2022). Early Pregnancy Modulates Expression of the Nod-like Receptor Family in Lymph Nodes of Ewes. Animals, 12(23), 3285. https://doi.org/10.3390/ani12233285