

Fatty Acid Composition of M. Biceps Femoris of Edible Dormouse (Glis glis L.)

 ,
,  , ,
, ,  ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Body Mass and Fatty Acid Composition of Muscle
3.2. Nutritional Fatty Acid Sums and Ratios
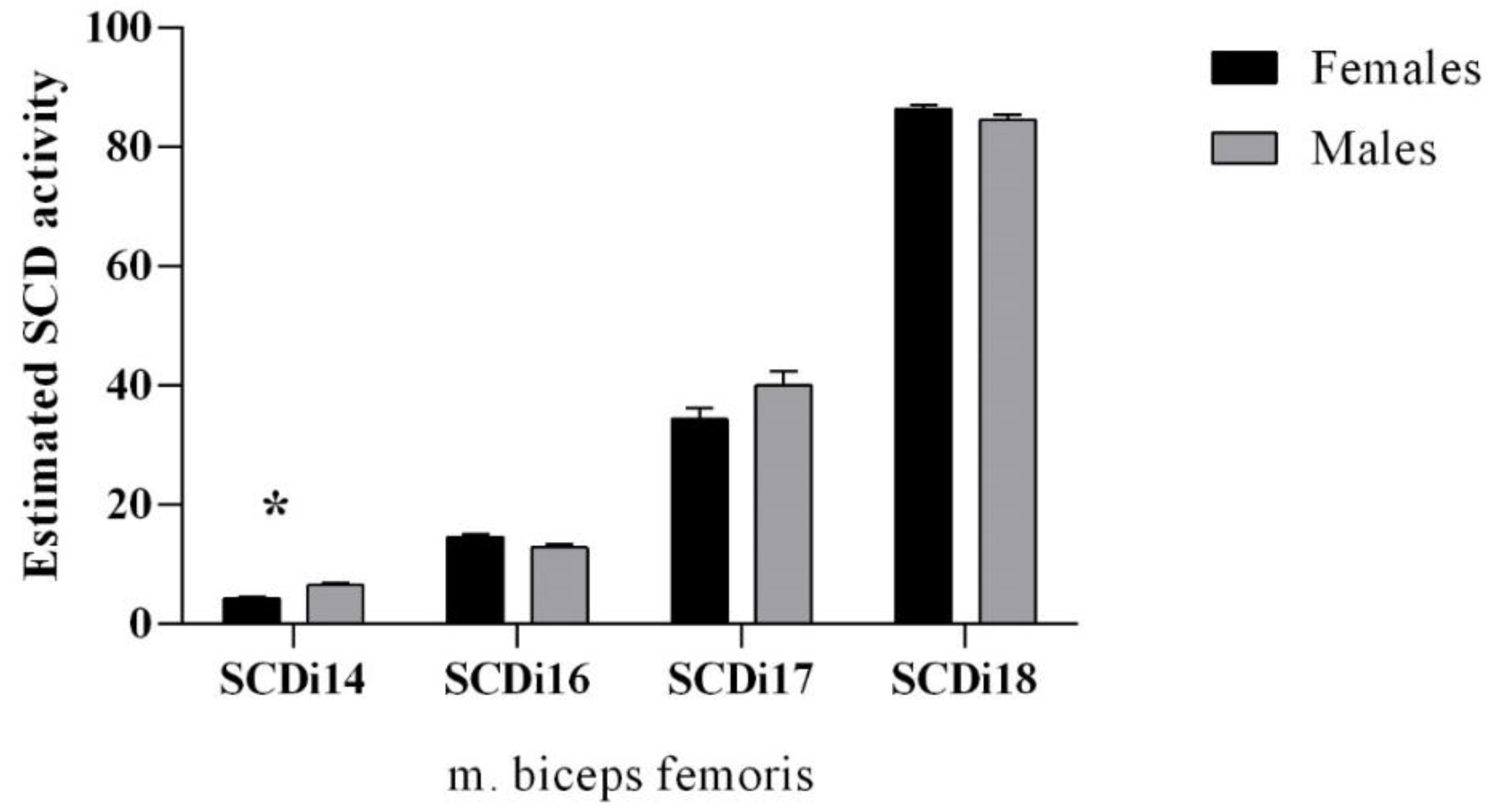
3.3. Stearoyl-CoA Desaturase Activity Indices
3.4. Figures and Tables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Storch, G. Gliridae. In Handbuch der Säugetiere Europas 1, Rodentia I; Niethammer, J., Krapp, F., Eds.; Akademische Verlagsgesellschaft: Wiesbaden, Germany, 1978; Volume 187, pp. 253–263. [Google Scholar]
- Forenbacher, S. Compendium of the Velebit Fauna; Veterinarski Fazkultet: Zagreb, Croatia, 2002; pp. 10–13. (In Croatian) [Google Scholar]
- Jurczyszyn, M.; Wolk, K. The present status of dormice (Myoxidae) in Poland. Nt. Croat. 1998, 7, 11–18. [Google Scholar]
- Grubešić, M.; Krapinec, K.; Glavaš, M.; Margaletić, J. Body measurements and harvesting dynamics of the fat dormouse (Glis glis, L.) in the mountainous part of Croatia. Acta Zool. Acad. Scie Hung. 2004, 50, 271–282. [Google Scholar]
- Konjević, D.; Krapinec, K. Fat dormouse (Glis glis, Linnaeus, 1766), from hunting to groceries. Meso 2004, 6, 59–63. [Google Scholar]
- Margaletić, J.; Grubešić, M.; Krapinec, K.; Kauzlarić, K.; Krajter, S. Dynamics and structure of fat dormouse (Glisglis L.) population in Croatia forests in the period from 2002 to 2004. Glas. Šum. Pokuse Posebno Izdanje 2006, 5, 377–386. [Google Scholar]
- Anonymous. Lovno gospodarska osnova za zajedničko otvoreno lovište broj XVII/135 (Hunting basis for common open hunting grounds number XVII/135). Vrgoračko jezero“ for period 01. Croatian Hunting Association, 10000 Zagreb Croatia, April 2008–31 March 2018. (In Croatian)
- Horvatić, S. Pregled vegetacije otoka Raba sa gledišta biljne sociologije (Overview of the vegetation of the island of Rab from the point of view of plant sociology). Prir. Istraž. Jugosl. Akad. 1939, 22, 1–96. (In Croatian) [Google Scholar]
- Horvatić, S. Tipološko raščlanjenje primorske vegetacije gariga i borovih šuma (Typology of coastal vegetation of garig and pine forests). Acta Bot Croat. 1958, 17, 1–98. (In Croatian) [Google Scholar]
- Tvrtković, N.; Dulić, B.; Grubešić, M. Distribution and habitats of Dormice in Croatia. Hystrix 1995, 6, 265–271. [Google Scholar]
- Cvrtila, Ž.; Konjević, D.; Kozačinski, L.; Hadziosmanović, M.; Slavica, A.; Margaletić, J. The chemical composition of the meat of fat dormice (Glis glis L.). Eur. J. Wildl. Res. 2004, 50, 90–91. [Google Scholar] [CrossRef]
- Bieber, C. Population dynamics, sexual activity, and reproductive failure in the fat dormouse (Myoxus glis). J. Zool. Lond. 1998, 244, 223–229. [Google Scholar] [CrossRef]
- Fietz, J.; Pflug, M.; Schlund, W.; Tataruch, F. Influences of the feeding ecology on body mass and possible implications for reproduction in the edible dormouse (Glis glis). J. Comp. Phys. B 2005, 175, 45–55. [Google Scholar] [CrossRef]
- Ruf, T.; Fietz, J.; Schlund, W.; Bieber, C. High survival in poor years: Life history tactics adapted to mast seeding in the edible dormouse. Ecology 2006, 87, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Schlund, W.; Scharfe, F.; Ganzhorn, J. Long-term comparison of food availability and reproduction in the edible dormouse (Glis glis). Z. Säugetierkd 2002, 67, 219–232. [Google Scholar] [CrossRef]
- Pilastro, A.; Gomiero, T.; Marin, G. Factors affecting body mass of young dormice (Glis glis) at weaning and by hibernation. J. Zool. 1994, 234, 13–23. [Google Scholar] [CrossRef]
- Bäumler, W.; Glavaš, M.; Margaletić, J. Schäden des Siebenschläfers an Waldbäumen. AFZ—DerWald 2002, 57, 204–206. [Google Scholar]
- Car, Z. Taxonomy, and biology of game. In Handbook of Hunting; Dragišić, P., Ed.; Lovačka Knjiga: Zagreb, Croatia, 1967; pp. 161–162. (In Croatian) [Google Scholar]
- Juškaitis, R.; Baltrūnaitė, L.; Augutė, V. Diet of the fat dormouse (Glis glis) on the northern periphery of its distributional range. Mamm. Res. 2015, 60, 155–161. [Google Scholar] [CrossRef]
- Perić, R. Značaj Puhova (por. Gliridae) u Šumama Hrvatske (The Significance of Dormice (por. gliridae) in the Forests of Croatia). Master’s Thesis, Faculty of Forestry, University of Zagreb, Department of Forestry, University Graduate Study, Zagreb, Croatia, 2006. (In Croatian). [Google Scholar]
- Turk, N.; Milas, Z.; Margaletić, J.; Turk, R.; Barbić, L.J.; Konjević, D.; Perić, S.; Stritof, Z.; Starešina, V. The role of fat dormouse (Glis glis L.) as reservoir host for spirochete Borrelia burgdorferi sensulato in the region of Gorski Kotar, Croatia. Eur. J. Wildl. Res. 2008, 54, 117–121. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Alves, S.P.; Raundrup, K.; Cabo, A.; Bessa, R.J.B.; Almeida, A.M. Fatty Acid Composition of Muscle, Adipose Tissue and Liver from Muskoxen (Ovibos moschatus) Living in West Greenland. PLoS ONE 2015, 10, e0145241. [Google Scholar] [CrossRef]
- Giroud, S.; Evans, A.L.; Chery, I.; Bertile, F.; Georg Tascher, G.; Bertrand-Michel, J.; Gauquelin-Koch, G.; Arnemo, J.M.; Swenson, J.E.; Lefai, E.; et al. Seasonal changes in eicosanoid metabolism in the brown bear. Sci. Nat. 2018, 105, 58. [Google Scholar] [CrossRef]
- Vranković, L.; Delaš, I.; Reljić, S.; Huber, Đ.; Stojević, Z.; Aladrović, J. Insight into Croatian brown bears health status from liver and muscle fatty acid composition perspective. In Proceedings of the Book of Abstract of 26th International Conference of Bear Research and Management, Ljubljana, Slovenia, 16–21 September 2018; p. 120. [Google Scholar]
- Kulagina, T.P.; Popova, S.S.; Aripovsky, A.V. Seasonal Changes in the Content of Fatty Acids in the Myocardium and m. longissimus dorsi of the Long-Tailed Ground Squirrel Urocitellus undulatus. Biophysics 2021, 66, 1004–1010. [Google Scholar] [CrossRef]
- Speakman, J.R.; Talbot, D.A.; Selman, C.; Snart, S.; McLaren, J.S.; Redman, P.; Krol, E.; Jackson, D.M.; Johnson, M.S.; Brand, M.D. Uncoupled and surviving: Individual mice with high metabolism have greater mitochondrial uncoupling and live longer. Aging Cell 2004, 3, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, A.J.; Reinald, P.; Buffenstein, R.; Buttemer, W.A. Life and Death: Metabolic Rate, Membrane Composition, and Life Span of Animals. Phys. Rev. 2007, 87, 1175–1213. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, A.J.; Rana, T.; Couture, P. The acyl composition of mammalian phospholipids: An allometric analysis. Comp. Biochem. Phys. B 2002, 132, 515–527. [Google Scholar] [CrossRef]
- Hulbert, A.J. Life, death and membrane bilayers. J. Exp. Biol. 2003, 206, 2303–2311. [Google Scholar] [CrossRef]
- Hulbert, A.J. On the importance of fatty acid composition of membranes for aging. J. Biol. 2005, 234, 277–288. [Google Scholar] [CrossRef]
- Trout, R.C.; Brooks, S.; Morris, P. Nest box usage by old edible dormice (Glis glis) in breeding and non-breeding years. Folia Zool. 2015, 64, 320–324. [Google Scholar] [CrossRef]
- Andersson, A.; SjoDin, A.; Hedman, A.; Olsson, R.; Vessby, B. Fatty Acid Profile Of Skeletal Muscle Phospholipids In Trained And Untrained Young Men. Am. J. Phys. Endocrinol. Metab. 2000, 279, E744–E751. [Google Scholar] [CrossRef]
- Pan, D.A.; Lillioja, S.; Milner, M.R.; Kriketos, A.D.; Baur, L.A.; Bogardus, C.; Storlien, L.H. Skeletal muscle membrane lipid composition is related to adiposity and insulin action. J. Clin. Invest. 1995, 96, 2802–2808. [Google Scholar] [CrossRef] [PubMed]
- Renaville, B.; Prandi, A.; Fan, B.; Sepulcri, A.; Rothschild, M.F.; Piasentier, E. Candidate gene marker associations with fatty acid profiles in heavy pigs. Meat. Sci. 2013, 93, 495–500. [Google Scholar] [CrossRef]
- Gol, S.; Pena, R.N.; Rothschild, M.F.; Tor, M.; Estany, J. A polymorphism in the fatty acid desaturase-2 gene is associated with the arachidonic acid metabolism in pigs. Sci. Rep. 2018, 8, 14336. [Google Scholar] [CrossRef]
- Blandine, C.; Storey, K.B.; Ziemianin, A.; Chanon, S.; Plume, M.; Chery, I.; Durand, C.; Evans, A.L.; Arnemo, J.M.; Zedrosser, A.; et al. Metabolic reprogramming involving glycolysis in the hibernating brown bear skeletal muscle. Front. Zool. 2019, 16, 12. [Google Scholar] [CrossRef]
- Murphy, M.G. Dietary fatty acids and membrane protein function. J. Nutr. Biochem. 1990, 1, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Peters, T.; Kubis, H.P.; Wetzel, P.; Sender, S.; Asmussen, G.; Fons, R.; Jürgens, K.D. Contraction parameters, myosin composition and metabolic enzymes of the skeletal muscles of the etruscan shrew Suncus etruscus and of the common European white-toothed shrew Crocidura russula (Insectivora: Soricidae). J. Exp. Biol. 1999, 202, 2461–2473. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-H.; Lee, Y.S.; Oh, Y.K.; Jung, N.-P.; Gwag, B.J.; Shin, H.C. Thermogenesis and motor recruitment of the pectoral muscle during shivering in arousing bats Murina leucogaster. Kor. J. Biol. Sci. 2001, 5, 31–35. [Google Scholar] [CrossRef]
- Lee, K.; So, H.; Gwag, T.; Ju, H.; Lee, J.-W.; Yamashita, M.; Choi, I. Molecular Mechanism underlying muscle mass retention in hibernating bats: Role of periodic arousal. J. Cell Phys. 2010, 222, 313–319. [Google Scholar] [CrossRef]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Phys. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef]
- Frank, C.L.; Dierenfeld, E.S.; Storey, K.B. The relationship between lipid peroxidation, hibernation, and food selection in mammals. Am. Zool. 1998, 38, 341–349. [Google Scholar] [CrossRef]
- Janovská, A.; Hatzinikolas, G.; Mano, M.; Wittert, G.A. The effect of dietary fat content on phospholipid fatty acid profile is muscle fiber type dependent. Am. J. Phys. Endocrinol. Metab. 2010, 298, E779–E786. [Google Scholar] [CrossRef]
- Wolff, R.L.; Pédrono, F.; Marpeau, A.M.; Christie, W.W.; Gunstone, F.D. The seed fatty acid composition and the distribution of ∆5- olefinic acids in the triacylglyerols of some Taxaceae (Taxus and Torreya). J. Am. Oil Chem. Soc. 1998, 75, 1637–1641. [Google Scholar] [CrossRef]
- Wolff, R.L.; Christie, W.W.; Aitzetmuller, K.; Pasquier, E.; Pedrono, F.; Destaillats, F.; Marpeau, A.M. Arachidonic and eicosapentaenoic acids in Araucariaceae, a unique feature among seed plants. Oléagineux Corps. Gras Lipides 2000, 7, 113–117. [Google Scholar] [CrossRef][Green Version]
- Berger, A.; Monnard, I.; Baur, M.; Charbonnet, C.; Safonova, I.; Jomard, A. Epidermal anti-Inflammatory properties of 5,11,14 20:3: Effects on mouse ear edema, PGE2 levels in cultured keratinocytes, and PPAR activation. Lipids Health Dis. 2002, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Ekman, R. New Polyenoic Fatty Acids in Norway Spruce Wood. Phytochemistry 1980, 19, 147–148. [Google Scholar] [CrossRef]
- Janicki, Z.; Slavica, A.; Konjević, D.; Severin, K. Game zoology [in Croatian]. Zavod za biologiju, patologiju i uzgoj divljači. Veterinarski fakultet. Sveučilište u Zagrebu. Croatia 2007, 2007, 73–76. [Google Scholar]
- Hoelzl, F.; Bieber, C.; Cornils, J.S.; Gerritsmann, H.; Stalder, G.L.; Walzer, C.; Ruf, T. How to spend the summer? Free-living dormice (Glis glis) can hibernate for 11 months in non-reproductive years. J. Comp. Phys. B 2015, 185, 931–939. [Google Scholar] [CrossRef]
- Prasad, R.B.N.; Gülz, P.-G. Composition of Lipids of Beech (Fagus sylvatica L.) Seed Oil. Z. Naturforsch. 1989, 44, 735–738. [Google Scholar] [CrossRef]
- Hulbert, A.J.; Abbott, S.K. Nutritional ecology of essential fatty acids: An evolutionary perspective. Aust. J. Zool. 2011, 59, 369–379. [Google Scholar] [CrossRef]
- Stumpfel, S.; Bieber, C.; Blanc, S.; Ruf, T.; Giroud, S. Differences in growth rates and prehibernation body mass gain between early and lateborn juvenile garden dormice. J. Comp. Phys. B 2017, 187, 1. [Google Scholar] [CrossRef]
- Frank, C.L. Polyunsaturate content and diet selection by ground squirrels (Spermophilus lateralis). Ecology 1994, 75, 458–463. [Google Scholar] [CrossRef]
- Frank, C.L. Short-term variations in diet fatty acid composition and torpor by ground squirrels. J. Mamm. 2002, 83, 1013–1019. [Google Scholar] [CrossRef]
- Geiser, F. The effect of unsaturated and saturated dietary lipids on the pattern of daily torpor and the fatty acid composition of tissues and membranes of the deer mouse Peromyscus maniculatus. J. Comp. Phys. B Biochem. Syst. Environ. Phys. 1991, 161, 590–597. [Google Scholar] [CrossRef]
- Frank, C.L. The influence of dietary fatty acids on hibernation by golden-mantled ground squirrels (Spermophilus lateralis). Phys. Zool. 1992, 65, 906–920. [Google Scholar] [CrossRef]
- DePeters, E.J.; Medrano, J.F.; Reed, B.A. Fatty acid composition of milk fat from three breeds of dairy cattle. Can. J. Anim. Sci. 1995, 75, 267–269. [Google Scholar] [CrossRef]
- Kryštufek, B. Glis glis (Rodentia: Gliridae). Mammal. Spec. 2000, 42, 195–206. [Google Scholar] [CrossRef]
- Čanády, A.; Mošanský, L.; Krišovský, P. Sexual size monomorphism and body variation in the fat dormouse Glis glis in Slovakia. Biologia 2016, 71, 1061–1066. [Google Scholar] [CrossRef]
- Nandini, R. Evolution of Sexual Size Dimorphism in Squirrels. Doctoral Dissertation, Smithsonian Institution, Washington, DC, USA, Auburn University, Auburn, AL, USA, 2011; 121p. [Google Scholar]
- Rieger, J.F. Body size, litter size, timing of reproduction, and juvenile survival in the Uinta ground squirrel, Spermophilus armatus. Oecologia 1996, 107, 463–468. [Google Scholar] [CrossRef]
- Schaefer, A.; Piquard, F.; Haberey, F. Food self-selection during spontaneous body weight variations in the dormouse (Glis glis L.). Comp. Biochem. Phys. A 1976, 55, 115–118. [Google Scholar] [CrossRef]
- Dark, J. Annual lipid cycles in hibernators: Integration of physiology and behavior. Annu. Rev. Nutr. 2005, 25, 469–497. [Google Scholar] [CrossRef]
- FAO. Fats and Fatty Acids in Human Nutrition (FAO Report of an Expert Consultation); FAO: Rome, Italy, 2010. [Google Scholar]
- ISSFAL. Global Recommendations. 2004. Available online: http://www.issfal.org.uk/recommendations-of-others.Html (accessed on 15 February 2022).
- Kang, M.J.; Shin, M.S.; Park, J.N.; Lee, S.S. The effects of polyunsaturated:saturated fatty acids ratios and peroxidisability index values of dietary fats on serum lipid profiles and hepatic enzyme activities in rats. Brit. J. Nutr. 2005, 94, 526–532. [Google Scholar] [CrossRef]
- Hayes, K.C. Dietary fat and heart health: In search of the ideal fat. Asia Pac. J. Clin. Nutr. 2002, 11, S394–S400. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Females (n = 16) | Males (n = 14) | p-Value | |
|---|---|---|---|
| Saturated FA | |||
| 14:0 | 0.78 ± 0.18 | 0.89 ± 0.15 | 0.402 |
| 15:0 | 0.018 ± 0.004 | 0.019 ± 0.004 | 0.567 |
| 16:0 | 13.2 ± 0.85 | 17.7 ± 0.74 | 0.782 |
| 17:0 | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.743 |
| 18:0 | 5.90 ± 1.26 | 8.84 ± 1.10 | 0.746 |
| 20:0 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.254 |
| Monounsaturated FA | |||
| 14:1cis-9 | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.567 |
| 16:1cis-7 | 0.36 ± 0.08 | 0.20 ± 0.07 | 0.600 |
| 16:1cis-9 | 2.73 ± 0.61 | 2.51 ± 0.53 | 0.840 |
| 17:1cis-9 | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.510 |
| 18:1cis-9 | 63.0 ± 6.50 | 47.7 ± 5.63 | 0.164 |
| 18:1cis-11 | 2.04 ± 0.20 | 2.32 ± 0.18 | 0.783 |
| Polyunsaturated FA | |||
| 18:2 n-6 | 8.56 ± 3.57 | 11.4 ± 3.09 | 0.063 |
| 18:3 n-3 | 1.48 ± 0.22 | 0.93 ± 0.19 | 0.480 |
| 20:2 n-6 | 0.12 ± 0.05 | 0.14 ± 0.04 | 0.244 |
| 20:3 ∆5,∆11,∆14 | 0.10 ± 0.10 | 0.14 ± 0.08 | 0.835 |
| 20:3 n-6 | 0.11 ± 0.12 | 0.36 ± 0.10 | 0.998 |
| 20:3 n-3 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.196 |
| 20:4 n-6 (ARA) | 0.89 ± 1.67 | 4.03 ± 1.45 | 0.380 |
| 20:5 n-3 (EPA) | 0.02 ± 0.03 | 0.09 ± 0.02 | 0.226 |
| 22:4 n-6 | 0.04 ± 0.03 | 0.11 ± 0.03 | 0.890 |
| 22:5 n-6 | 0.03 ± 0.03 | 0.13 ± 0.02 | 0.190 |
| 22:5 n-3 (DPA) | 0.11 ± 0.15 | 0.41 ± 0.13 | 0.423 |
| 22:6 n-3 (DHA) | 0.38 ± 0.80 | 1.97 ± 0.69 | 0.406 |
| Females (n = 16) | Males (n = 14) | p-Value | |
|---|---|---|---|
| Sums | |||
| SFA | 20.0 ± 1.27 | 27.6 ± 1.10 | 0.835 |
| UFA | 79.6 ± 1.27 | 72.5 ± 1.10 | 0.835 |
| MUFA | 68.2 ± 6.83 | 52.8 ± 5.92 | 0.189 |
| PUFA | 11.8 ± 6.07 | 19.6 ± 5.25 | 0.099 |
| n-6 PUFA | 9.76 ± 5.25 | 16.2 ± 4.54 | 0.119 |
| n-3 PUFA | 2.01 ± 0.90 | 3.43 ± 0.78 | 0.058 |
| VLn-3PUFA | 0.97 ± 0.24 | 1.73 ± 0.21 | 0.102 |
| DHA + EPA | 0.76 ± 0.16 | 1.42 ± 0.14 | 0.133 |
| Ratios | |||
| UFA/SFA | 4.00 ± 0.20 | 2.65 ± 0.17 | 0.835 |
| PUFA/SFA | 0.66 ± 0.09 | 0.91 ± 0.09 | 0.059 |
| 18:1/18:2 | 8.2 ± 1.22 | 4.45 ± 1.30 | 0.068 |
| AA/EPA | 131 ±66.37 | 59.6 ± 57.48 | 0.624 |
| AA/DHA | 2.38 ± 0.29 | 2.06 ± 0.25 | 0.065 |
| EPA/DHA | 0.05 ± 0.03 | 0.07 ± 0.03 | 0.522 |
| n-6/n-3 | 4.69 ± 0.92 | 4.80 ± 0.80 | 0.888 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pađen, L.; Alves, S.P.; Bessa, R.J.B.; Almeida, A.M.; Bujanić, M.; Konjević, D. Fatty Acid Composition of M. Biceps Femoris of Edible Dormouse (Glis glis L.). Animals 2022, 12, 3284. https://doi.org/10.3390/ani12233284
Pađen L, Alves SP, Bessa RJB, Almeida AM, Bujanić M, Konjević D. Fatty Acid Composition of M. Biceps Femoris of Edible Dormouse (Glis glis L.). Animals. 2022; 12(23):3284. https://doi.org/10.3390/ani12233284
Chicago/Turabian StylePađen, Lana, Susana P. Alves, Rui J. B. Bessa, André M. Almeida, Miljenko Bujanić, and Dean Konjević. 2022. "Fatty Acid Composition of M. Biceps Femoris of Edible Dormouse (Glis glis L.)" Animals 12, no. 23: 3284. https://doi.org/10.3390/ani12233284
APA StylePađen, L., Alves, S. P., Bessa, R. J. B., Almeida, A. M., Bujanić, M., & Konjević, D. (2022). Fatty Acid Composition of M. Biceps Femoris of Edible Dormouse (Glis glis L.). Animals, 12(23), 3284. https://doi.org/10.3390/ani12233284