
Effect of Supplementation with Organic Selenium or Turmeric and Rosemary Mixture on Beta-Defensin Content in Goat Milk

 , ,
, ,  , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Methods
2.3. Statistical Analysis
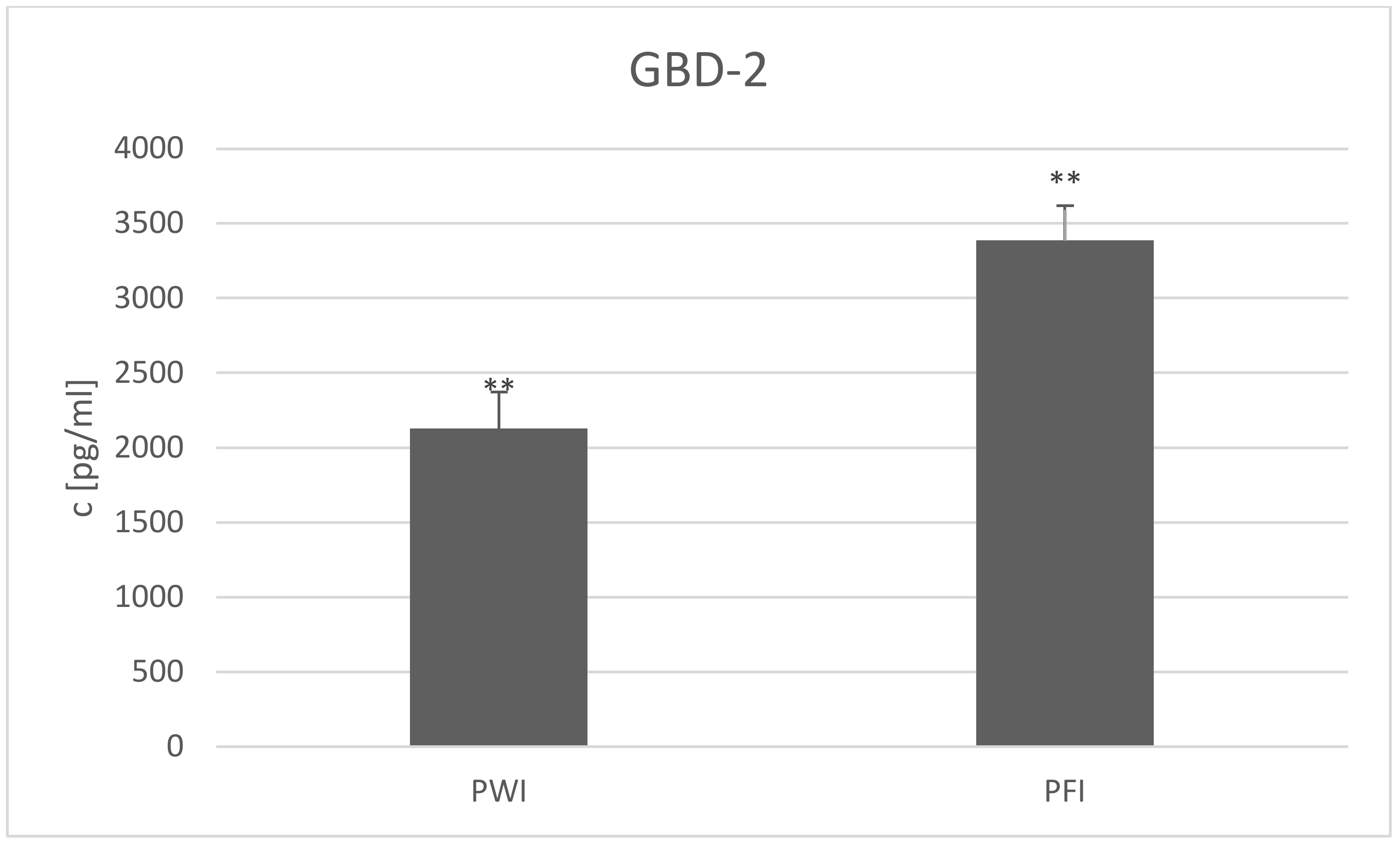
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Venkataraman, N.; Cole, A.L.; Ruchala, P.; Waring, A.J.; Lehrer, R.I.; Stuchlik, O.; Pohl, J.; Cole, A.M. Reawakening Retrocyclins: Ancestral Human Defensins Active Against HIV-1. PLoS Biol. 2009, 7, e1000095. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Lu, W. Defensins: A Double-Edged Sword in Host Immunity. Front. Immunol. 2020, 11, 764. [Google Scholar] [CrossRef] [PubMed]
- Bagnicka, E.; Strzałkowska, N.; Jóźwik, A.; Krzyżewski, J.; Horbańczuk, J.; Zwierzchowski, L. Expression and Polymorphism of Defensins in Farm Animals. Acta Biochim. Pol. 2010, 57, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Jarczak, J.; Kościuczuk, E.M.; Lisowski, P.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Defensins: Natural Component of Human Innate Immunity. Hum. Immunol. 2013, 74, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Riera Romo, M.; Pérez-Martínez, D.; Castillo Ferrer, C. Innate Immunity in Vertebrates: An Overview. Immunology 2016, 148, 125–139. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial Peptides of Innate Immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Selsted, M.E.; Ouellette, A.J. Mammalian Defensins in the Antimicrobial Immune Response. Nat. Immunol. 2005, 6, 551–557. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial Peptides: Pore Formers or Metabolic Inhibitors in Bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Kudryashova, E.; Quintyn, R.; Seveau, S.; Lu, W.; Wysocki, V.H.; Kudryashov, D.S. Human Defensins Facilitate Local Unfolding of Thermodynamically Unstable Regions of Bacterial Protein Toxins. Immunity 2014, 41, 709–721. [Google Scholar] [CrossRef]
- Harder, J.; Bartels, J.; Christophers, E.; Schröder, J.-M. A Peptide Antibiotic from Human Skin. Nature 1997, 387, 861. [Google Scholar] [CrossRef]
- Wei, G.; de Leeuw, E.; Pazgier, M.; Yuan, W.; Zou, G.; Wang, J.; Ericksen, B.; Lu, W.-Y.; Lehrer, R.I.; Lu, W. Through the Looking Glass, Mechanistic Insights from Enantiomeric Human Defensins. J. Biol. Chem. 2009, 284, 29180–29192. [Google Scholar] [CrossRef]
- Fjell, C.D.; Jenssen, H.; Fries, P.; Aich, P.; Griebel, P.; Hilpert, K.; Hancock, R.E.W.; Cherkasov, A. Identification of Novel Host Defense Peptides and the Absence of Alpha-Defensins in the Bovine Genome. Proteins 2008, 73, 420–430. [Google Scholar] [CrossRef]
- Lisitsyn, N.A.; Bukurova, Y.A.; Nikitina, I.G.; Krasnov, G.S.; Sykulev, Y.; Beresten, S.F. Enteric Alpha Defensins in Norm and Pathology. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 1. [Google Scholar] [CrossRef]
- Zhang, L.; Xiao, H.; Huang, J.; Ouyang, L.; Li, S.; Tang, Y. Identification and Expression Analysis of the β-Defensin Genes in the Goat Small Intestine. Gene 2021, 801, 145846. [Google Scholar] [CrossRef]
- Bagnicka, E.; Prusak, B.; Kościuczuk, E.; Jarczak, J.; Kaba, J.; Strzałkowska, N.; Jóźwik, A.; Czopowicz, M.; Krzyżewski, J.; Zwierzchowski, L. A Note on the Organization and Expression of β-Defensin Genes in Polish Goats. J. Appl. Genet. 2013, 54, 125–127. [Google Scholar] [CrossRef][Green Version]
- Swanson, K.; Gorodetsky, S.; Good, L.; Davis, S.; Musgrave, D.; Stelwagen, K.; Farr, V.; Molenaar, A. Expression of a β-Defensin MRNA, Lingual Antimicrobial Peptide, in Bovine Mammary Epithelial Tissue Is Induced by Mastitis. Infect. Immun. 2004, 72, 7311–7314. [Google Scholar] [CrossRef]
- Zhao, C.; Nguyen, T.; Liu, L.; Shamova, O.; Brogden, K.; Lehrer, R.I. Differential Expression of Caprine Beta-Defensins in Digestive and Respiratory Tissues. Infect. Immun. 1999, 67, 6221–6224. [Google Scholar] [CrossRef]
- Roosen, S.; Exner, K.; Paul, S.; Schröder, J.-M.; Kalm, E.; Looft, C. Bovine Beta-Defensins: Identification and Characterization of Novel Bovine Beta-Defensin Genes and Their Expression in Mammary Gland Tissue. Mamm. Genome 2004, 15, 834–842. [Google Scholar] [CrossRef]
- Conner, A.E. Elevated Levels of Sodium and Chloride in Milk from Mastitic Breast. Pediatrics 1979, 63, 910–911. [Google Scholar] [CrossRef]
- Ma, Y.; Ryan, C.; Barbano, D.M.; Galton, D.M.; Rudan, M.A.; Boor, K.J. Effects of Somatic Cell Count on Quality and Shelf-Life of Pasteurized Fluid Milk. J. Dairy Sci. 2000, 83, 264–274. [Google Scholar] [CrossRef]
- Krzyzewski, J.; Bagnicka, E.; Horbanczuk, J.O. The Effect of Selenium Supplementation to the Diet of Dairy Cows and Goats on Production Traits and Animal Health—A Review. Anim. Sci. Pap. Rep. 2014, 32, 283–299. [Google Scholar]
- Arthur, J.R.; McKenzie, R.C.; Beckett, G.J. Selenium in the Immune System. J. Nutr. 2003, 133, 1457S–1459S. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, M.N.; Monahan, F.J.; Fallon, R.J.; Allen, P. Effects of Dietary Supplementation with Vitamin E and Organic Selenium on the Oxidative Stability of Beef. J. Anim. Sci. 2001, 79, 2827–2834. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M. Selenium-Dependent Regulation of Oxidative Stress and Immunity in Periparturient Dairy Cattle. Vet. Med. Int. 2013, 2013, 154045. [Google Scholar] [CrossRef] [PubMed]
- Reczyńska, D.; Witek, B.; Jarczak, J.; Czopowicz, M.; Mickiewicz, M.; Kaba, J.; Zwierzchowski, L.; Bagnicka, E. The Impact of Organic vs. Inorganic Selenium on Dairy Goat Productivity and Expression of Selected Genes in Milk Somatic Cells. J. Dairy Res. 2019, 86, 48–54. [Google Scholar] [CrossRef]
- Tufarelli, V.; Laudadio, V. Dietary Supplementation with Selenium and Vitamin E Improves Milk Yield, Composition and Rheological Properties of Dairy Jonica Goats. J. Dairy Res. 2011, 78, 144–148. [Google Scholar] [CrossRef]
- Nurdin, E.; Amelia, T.; Makin, M. The Effects of Herbs on Milk Yield and Milk Quality of Mastitis Dairy Cow. J. Indones. Trop. Anim. Agric. 2011, 36, 104–108. [Google Scholar] [CrossRef][Green Version]
- Mamun, M.; Juyena, N.; Khan, M.A.H.N.A.; Islam, M.A. Comparative Effect of Turmeric (Curcuma longa) and Durba (Cynodon dactylon) on the Healing of Surgical Wounds in Cattle (Bos indicus). Medicine 2018, 4, 164–171. [Google Scholar] [CrossRef]
- Boutoial, K.; García, V.; Rovira, S.; Ferrandini, E.; Abdelkhalek, O.; López, M.B. Effect of Feeding Goats with Distilled and Non-Distilled Thyme Leaves (Thymus zygis subp. gracilis) on Milk and Cheese Properties. J. Dairy Res. 2013, 80, 448–456. [Google Scholar] [CrossRef]
- Tsugami, Y.; Ishiba, Y.; Suzuki, N.; Nii, T.; Kobayashi, K.; Isobe, N. Local Heat Treatment of Goat Udders Influences Innate Immune Functions in Mammary Glands. J. Mammary Gland Biol. Neoplasia 2021, 26, 387–397. [Google Scholar] [CrossRef]
- Jarczak, J.; Kościuczuk, E.; Ostrowska, M.; Lisowski, P.; Strzałkowska, N.; Jóźwik, A.; Krzyżewski, J.; Zwierzchowski, L.; Słoniewska, D.; Bagnicka, E. The Effects of Diet Supplementation with Yeast on the Expression of Selected Immune System Genes in the Milk Somatic Cells of Dairy Goats. Anim. Sci. Pap. Rep. 2014, 32, 41–53. [Google Scholar]
- Brzóska, F.; Kowalski, Z.M.; Osięgłowski, S.; Strzetelski, J. IZ PIB-INRA Zalecenia Żywieniowe dla Przeżuwaczy i Tabele Wartości Pokarmowej Pasz; Strzetelski, J., Ed.; Fundacja Instytutu Zootechniki Państwowego Instytutu Badawczego Patronus Animalium: Warsaw, Poland, 2014; ISBN 978-83-938377-0-0. [Google Scholar]
- Janštová, B.; Dračková, M.; Navrátilová, P.; Hadra, L.; Vorlová, L. Freezing Point of Raw and Heat-Treated Goat Milk. Czech. J. Anim. Sci. 2008, 52, 394–398. [Google Scholar] [CrossRef]
- Hanuš, O.; Hanušová, K.; Vyletělová, M.; Kopec, T.; Janů, L.; Kopecký, J. Selected Abiotic Factors That Influence Raw Cow Milk Freezing Point Depression. Acta Vet. Brno 2012, 81, 49–55. [Google Scholar] [CrossRef]
- Górecka-Bruzda, A.; Reczyńska, D.; Jastrzębska, E.; Barłowska, K.; Bagnicka, E. Behavioral and Physiological Measures in Dairy Goats with and without Small Ruminant Lentivirus Infection. J. Vet. Behav. 2019, 31, 67–73. [Google Scholar] [CrossRef]
- Kapusta, A.; Kuczyńska, B.; Puppel, K. Relationship between the Degree of Antioxidant Protection and the Level of Malondialdehyde in High-Performance Polish Holstein-Friesian Cows in Peak of Lactation. PLoS ONE 2018, 13, e0193512. [Google Scholar] [CrossRef]
- Puppel, K.; Kapusta, A.; Kuczyńska, B. The Etiology of Oxidative Stress in the Various Species of Animals, a Review. J. Sci. Food Agric. 2015, 95, 2179–2184. [Google Scholar] [CrossRef]
- Lu, C.D.; Kawas, J.R.; Mahgoub, O.G. Fibre Digestion and Utilization in Goats. Small Rumin. Res. 2005, 60, 45–52. [Google Scholar] [CrossRef]
- Santini, F.J.; Lu, C.D.; Potchoiba, M.J.; Coleman, S.W. Effects of Acid Detergent Fiber Intake on Early Postpartum Milk Production and Chewing Activities in Dairy Goats Fed Alfalfa Hay. Small Rumin. Res. 1991, 6, 63–71. [Google Scholar] [CrossRef]
- Zschiesche, M.; Mensching, A.; Sharifi, A.R.; Hummel, J. The Milk Fat-to-Protein Ratio as Indicator for Ruminal PH Parameters in Dairy Cows: A Meta-Analysis. Dairy 2020, 1, 259–268. [Google Scholar] [CrossRef]
- Bagnicka, E.; Hamann, H.; Distl, O. Structure and the Non-Genetic and Genetic Effects on Milk Traits in Polish Dairy Goat Population. Anim. Sci. Pap. Rep. 2015, 33, 59–69. [Google Scholar]
- Yoon, J.T.; Lee, J.H.; Kim, C.K.; Chung, Y.C.; Kim, C.-H. Effects of Milk Production, Season, Parity and Lactation Period on Variations of Milk Urea Nitrogen Concentration and Milk Components of Holstein Dairy Cows. Asian-Australas. J. Anim. Sci. 2004, 17, 479–484. [Google Scholar] [CrossRef]
- Zeng, S.S.; Escobar, E.N. Effect of Parity and Milk Production on Somatic Cell Count, Standard Plate Count and Composition of Goat Milk. Small Rumin. Res. 1995, 17, 269–274. [Google Scholar] [CrossRef]
- Bagnicka, E.; Łukaszewicz, M.; Ådnøy, T. Genetic Parameters of Somatic Cell Score and Lactose Content in Goat’s Milk. J. Anim. Feed Sci. 2016, 25, 210–215. [Google Scholar] [CrossRef]
- Goetsch, A.L.; Zeng, S.S.; Gipson, T.A. Factors Affecting Goat Milk Production and Quality. Small Rumin. Res. 2011, 101, 55–63. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, X.R.; Liu, J.Z.; An, X.P.; Zhou, Z.Q.; Cao, B.Y.; Song, Y.X. Supplemented Organic and Inorganic Selenium Affects Milk Performance and Selenium Concentration in Milk and Tissues in the Guanzhong Dairy Goat. Biol. Trace Elem. Res. 2018, 183, 254–260. [Google Scholar] [CrossRef]
- Rashnoo, M.; Rahmati, Z.; Azarfar, A.; Fadayifar, A. The Effects of Maternal Supplementation of Selenium and Iodine via Slow-Release Blouses in Late Pregnancy on Milk Production of Goats and Performance of Their Kids. Ital. J. Anim. Sci. 2020, 19, 502–513. [Google Scholar] [CrossRef]
- Kachuee, R.; Abdi-Benemar, H.; Mansoori, Y.; Sánchez-Aparicio, P.; Seifdavati, J.; Elghandour, M.M.M.Y.; Guillén, R.J.; Salem, A.Z.M. Effects of Sodium Selenite, L-Selenomethionine, and Selenium Nanoparticles During Late Pregnancy on Selenium, Zinc, Copper, and Iron Concentrations in Khalkhali Goats and Their Kids. Biol. Trace Elem. Res. 2019, 191, 389–402. [Google Scholar] [CrossRef]
- Chiofalo, V.; Liotta, L.; Fiumanò, R.; Riolo, E.B.; Chiofalo, B. Influence of Dietary Supplementation of Rosmarinus Officinalis L. on Performances of Dairy Ewes Organically Managed. Small Rumin. Res. 2012, 104, 122–128. [Google Scholar] [CrossRef]
- Hashemzadeh-Cigari, F.; Khorvash, M.; Ghorbani, G.R.; Kadivar, M.; Riasi, A.; Zebeli, Q. Effects of Supplementation with a Phytobiotics-Rich Herbal Mixture on Performance, Udder Health, and Metabolic Status of Holstein Cows with Various Levels of Milk Somatic Cell Counts. J. Dairy Sci. 2014, 97, 7487–7497. [Google Scholar] [CrossRef]
- Abdullah, M.A.M.; Daghash, M.W.H.; Bakheet, M.G.M.; Moneeb, A.H.M. Effect of Dietary Lemongrass (Cymbopogon citratus) and Roselle (Hibiscus sabdariffa) to Saidi Ewes on: (A) Milk Production, Rumen Fermentation and Lamb’s Performance. Egypt. J. Anim. Prod. 2020, 57, 47–53. [Google Scholar] [CrossRef]
- Abd EL Tawab, A.M.; Kholif, A.M.; El-Bordeny, N.E.; Elsayed, H.M.; Selim, N.A.H. Feed Utilization and Lactational Performance of Damascus Goats Fed a Diet Supplemented with Marjoram or Basil Essential Oils. Egypt. J. Chem. 2022, 65, 407–418. [Google Scholar] [CrossRef]
- Kholif, A.E.; Hamdon, H.A.; Kassab, A.Y.; Farahat, E.S.A.; Azzaz, H.H.; Matloup, O.H.; Mohamed, A.G.; Anele, U.Y. Chlorella Vulgaris Microalgae and/or Copper Supplementation Enhanced Feed Intake, Nutrient Digestibility, Ruminal Fermentation, Blood Metabolites and Lactational Performance of Boer Goat. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1595–1605. [Google Scholar] [CrossRef]
- Ghoneem, W.; Mahmoud, A. Impact of Incorporating Thymus Vulgaris as Leaves or Essential Oil in Damascus Goats Ration on Lactation Performance. Int. J. Dairy Sci. 2022, 17, 1–12. [Google Scholar] [CrossRef]
- Amosu, S.D.; Oluwatosin, B.O.; Fasae, O.A.; Ajadi, T.A.; Oderinwale, O.A.; Jolaosho, O.O. Performance Characteristics of Pregnant Goats Fed Diets Containing Turmeric (Curcuma longa) Powder Supplementation. Agric. Sci. Dig. 2020, 40, 178–183. [Google Scholar]
- Oderinwale, O.A.; Oluwatosin, B.O.; Onagbesan, M.O.; Adekunle, E.O.; Shuaibu, A.Y.; Amosu, S.D.; Adeyemo, A.J.; Kuye, O.M.; Olalere, J.O.; Ajewole, I.T. Effects of Dietary Inclusion of Turmeric (Curcuma longa L.) Powder on Oxidative Stress and Cortisol Concentration in Goats during Pregnancy and Onset of Postpartum. Niger. J. Anim. Prod. 2021, 48, 374–390. [Google Scholar] [CrossRef]
- Barroso, F.G.; Alados, C.L.; Boza, J. Social Hierarchy in the Domestic Goat: Effect on Food Habits and Production. Appl. Anim. Behav. Sci. 2000, 69, 35–53. [Google Scholar] [CrossRef]
- Jarczak, J.; Kościuczuk, E.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Effect of Organic vs. Inorganic Selenium Supplementation on Expression of Selected Defensin Genes in Somatic Milk Cells of Dairy Cows. J. Agric. Sci. Technol. A 2014, 4, 686–695. [Google Scholar]
- Maier, T.; Güell, M.; Serrano, L. Correlation of MRNA and Protein in Complex Biological Samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef]
- McInturff, J.E.; Modlin, R.L.; Kim, J. The Role of Toll-like Receptors in the Pathogenesis and Treatment of Dermatological Disease. J. Investig. Dermatol. 2005, 125, 1–8. [Google Scholar] [CrossRef]
- Romano Carratelli, C.; Mazzola, N.; Paolillo, R.; Sorrentino, S.; Rizzo, A. Toll-like Receptor-4 (TLR4) Mediates Human Beta-Defensin-2 (HBD-2) Induction in Response to Chlamydia Pneumoniae in Mononuclear Cells. FEMS Immunol. Med. Microbiol. 2009, 57, 116–124. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, A.; Kumar, A.; Dev, K. Characterization of Goat Lingual Antimicrobial Peptide CDNA. J. Immunol. Immunopathol. 2010, 12, 42–51. [Google Scholar]
- Plawinska-Czarnak, J.; Zarzynska, J.; Majewska, A.; Jank, M.; Kaba, J.; Bogdan, J.; Anusz, K.; Bagnicka, E. Selected Tissues of Two Polish Goat Breeds Do Not Differ on Genomic Level. Anim. Sci. Pap. Rep. 2019, 37, 53–64. [Google Scholar]
- Chen, J.; Qi, S.; Guo, R.; Yu, B.; Chen, D. Different Messenger RNA Expression for the Antimicrobial Peptides Beta-Defensins between Meishan and Crossbred Pigs. Mol. Biol. Rep. 2010, 37, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Ryniewicz, Z.; Zwierzchowski, L.; Bagnicka, E.; Krzyzewski, J.; Strzalkowska, N. Preliminary Investigations on the Polymorphism of Defensin Genes in Cattle—Relation with Milk Somatic Cell Count. Anim. Sci. Pap. Rep. 2002, 20. [Google Scholar]
- Le Maréchal, C.; Thiéry, R.; Vautor, E.; Le Loir, Y. Mastitis Impact on Technological Properties of Milk and Quality of Milk Products—A Review. Dairy Sci. Technol. 2011, 91, 247–282. [Google Scholar] [CrossRef]
- Ryniewicz, Z.; Zwierzchowski, L.; Bagnicka, E.; Flisikowski, K.; Maj, A.; Krzyżewski, J.; Strzałkowska, N. Association of the Polymorphism at Defensin Gene Loci with Dairy Production Traits and Milk Somatic Cell Count in Black-and-White Cows. Anim. Sci. Pap. Rep. 2003, 21, 209–222. [Google Scholar]
- Bagnicka, E.; Winnicka, A.; Jóźwik, A.; Rzewuska, M.; Strzałkowska, N.; Kościuczuk, E.; Prusak, B.; Kaba, J.; Horbańczuk, J.; Krzyżewski, J. Relationship between Somatic Cell Count and Bacterial Pathogens in Goat Milk. Small Rumin. Res. 2011, 100, 72–77. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Herd I | Herd II | ||
|---|---|---|---|---|
| LSM | SE | LSM | SE | |
| Milk yield [kg/morning milking] | 2.35 A | 0.17 | 1.46 B | 0.13 |
| LnSCC# ≈ SCC | 6.98 ≈ 1.08 × 106 A | 0.41 | 5.04 ≈ 1.55 × 105 B | 0.31 |
| Fat [%] | 4.31 A | 0.23 | 2.17 B | 0.18 |
| Protein [%] | 2.79 A | 0.08 | 3.20 B | 0.06 |
| Casein [%] | 2.13 | 0.08 | 2.38 | 0.06 |
| Lactose [%] | 4.66 A | 0.06 | 4.99 B | 0.05 |
| TS [%] | 12.31 A | 0.27 | 10.64 B | 0.21 |
| SNF [%] | 8.25 | 0.12 | 8.47 | 0.09 |
| Urea [mg/L] | 157 A | 26 | 445 B | 20 |
| Citric acid [%] | 0.11 | 0.07 | 0.10 | 0.05 |
| FPD [°C] | −636 A | 6.01 | −596 B | 6.00 |
| FFA [mEKV/L] | 1.10 A | 0.06 | 0.57 B | 0.05 |
| Density [mg/mL] | 1024 A | 0.58 | 1027 B | 0.48 |
| Acidity [T] * | 16.68 | 0.59 | 16.25 | 0.59 |
| Trait | Sampling Time | Control Group | Experimental Group | SE |
|---|---|---|---|---|
| LSM | LSM | |||
| Milk yield [kg/morning milking] | beginning | 1.92 A | 2.06 A | 0.10 |
| end | 2.46 Ba | 2.80 Bb | ||
| LnSCC# ≈ SCC | beginning | 5.85 ≈ 3.50 × 105 | 5.55 ≈ 2.58 × 105 | 0.10 |
| end | 6.17 a ≈ 4.80 × 105 | 5.22 b ≈ 1.85 × 105 | ||
| Fat [%] | beginning | 5.21 A | 4.85 A | 0.14 |
| end | 3.94 | 3.80 B | ||
| Protein [%] | beginning | 3.09 | 3.01 | 0.06 |
| end | 2.99 | 2.97 | ||
| Casein [%] | beginning | 2.35 | 2.30 | 0.05 |
| end | 2.29 | 2.24 | ||
| Lactose [%] | beginning | 4.79 A | 4.77 A | 0.04 |
| end | 4.63 B | 4.46 B | ||
| TS [%] | beginning | 13.76 A | 13.29 A | 0.18 |
| end | 12.12 B | 11.80 B | ||
| SNF [%] | beginning | 8.70 A | 8.62 A | 0.08 |
| end | 8.40 B | 8.1 B | ||
| Urea [mg/L] | beginning | 91.60 A | 72.42 A | 12.9 |
| end | 155.55 B | 146.21 B | ||
| Citric acid [%] | beginning | 0.09 | 0.09 | 0.005 |
| end | 0.10 | 0.09 | ||
| FPD [−°C] | beginning | −598 A | −589 A | 4.83 |
| end | −630 Ba | −609 Bb | ||
| FFA [mEKV/L] | beginning | 1.35 A | 1.26 A | 0.07 |
| end | 0.96 B | 0.85 B | ||
| Density [mg/mL] | beginning | 1025.7 | 1025.4 A | 0.32 |
| end | 1024.8 | 1024.1 B | ||
| Acidity [T] * | beginning | 15.28 | 15.54 | 0.47 |
| end | 16.28 | 16.05 |
| Trait | Sampling Time | Control Group | Experimental Group | SE |
|---|---|---|---|---|
| LSM | LSM | |||
| Milk yield [kg/morning milking] | beginning | 1.24 | 1.26 | 0.13 |
| end | 1.38 | 1.37 | ||
| LnSCC# ≈ SCC | beginning | 5.33 ≈ 2.07 × 105 | 4.75 ≈ 1.16 × 105 | 0.31 |
| end | 5.44 ≈ 2.32 × 105 | 5.17 ≈ 1.76 × 105 | ||
| Fat [%] | beginning | 1.58 A | 1.92 a | 0.17 |
| end | 2.36 B | 2.57 b | ||
| Protein [%] | beginning | 3.08 | 3.03 | 0.05 |
| end | 3.15 | 3.06 | ||
| Casein [%] | beginning | 2.24 | 2.32 | 0.05 |
| end | 2.35 | 2.27 | ||
| Lactose [%] | beginning | 5.34 A | 5.24 A | 0.05 |
| end | 4.97 B | 4.95 B | ||
| TS [%] | beginning | 10.23 | 10.45 | 0.17 |
| end | 10.78 | 10.89 | ||
| SNF [%] | beginning | 8.76 a | 8.64 a | 0.07 |
| end | 8.42 b | 8.32 b | ||
| Urea [mg/L] | beginning | 307 A | 327 A | 15.48 |
| end | 455 B | 426 B | ||
| Citric acid [%] | beginning | 0.14 A | 0.13 A | 0.005 |
| end | 0.10 B | 0.09 B | ||
| FPD [°C] | beginning | −628 A | −615 a | 3.32 |
| end | −599 B | −601 b | ||
| FFA [mEKV/L] | beginning | 0.50 | 0.55 | 0.04 |
| end | 0.60 | 0.59 | ||
| Density [mg/mL] | beginning | 1027.4 | 1026.9 | 0.40 |
| end | 1026.8 | 1026.4 | ||
| Acidity [T] * | beginning | 13.82 A | 13.88 A | 0.41 |
| end | 16.29 B | 15.85 B |
| Trait | GBD-2 |
|---|---|
| Milk yield [kg/morning milking] | 0.42 ** |
| LnSCC # | - |
| Fat [%] | 0.56 ** |
| Protein [%] | −0.33 * |
| Casein [%] | - |
| Lactose [%] | −0.66 ** |
| TS [%] | 0.43 ** |
| SNF [%] | - |
| Urea [mg/L] | −0.48 ** |
| Citric acid [%] | - |
| FPD [°C] | 0.31 * |
| FFA [mEKV/L] | 0.49 ** |
| Density [mg/mL] | −0.47 ** |
| Acidity [T] ## | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalewska, M.; Kapusta, A.; Kawecka-Grochocka, E.; Urbańska, D.M.; Czopowicz, M.; Kaba, J.; Brzozowska, P.; Bagnicka, E. Effect of Supplementation with Organic Selenium or Turmeric and Rosemary Mixture on Beta-Defensin Content in Goat Milk. Animals 2022, 12, 2948. https://doi.org/10.3390/ani12212948
Zalewska M, Kapusta A, Kawecka-Grochocka E, Urbańska DM, Czopowicz M, Kaba J, Brzozowska P, Bagnicka E. Effect of Supplementation with Organic Selenium or Turmeric and Rosemary Mixture on Beta-Defensin Content in Goat Milk. Animals. 2022; 12(21):2948. https://doi.org/10.3390/ani12212948
Chicago/Turabian StyleZalewska, Magdalena, Aleksandra Kapusta, Ewelina Kawecka-Grochocka, Daria M. Urbańska, Michał Czopowicz, Jarosław Kaba, Paulina Brzozowska, and Emilia Bagnicka. 2022. "Effect of Supplementation with Organic Selenium or Turmeric and Rosemary Mixture on Beta-Defensin Content in Goat Milk" Animals 12, no. 21: 2948. https://doi.org/10.3390/ani12212948
APA StyleZalewska, M., Kapusta, A., Kawecka-Grochocka, E., Urbańska, D. M., Czopowicz, M., Kaba, J., Brzozowska, P., & Bagnicka, E. (2022). Effect of Supplementation with Organic Selenium or Turmeric and Rosemary Mixture on Beta-Defensin Content in Goat Milk. Animals, 12(21), 2948. https://doi.org/10.3390/ani12212948