

Effects of N Acetylcysteine on the Expression of Genes Associated with Reproductive Performance in the Goat Uterus during Early Gestation

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Sample Collection
2.3. Total RNA Extraction and RNA Sequencing
2.4. Quantitative Reverse Transcription PCR
2.5. Statistical Analysis
3. Results
3.1. Effect of N-Acetylcysteine on the Reproductive Performance of Qianbei Ma Goats
3.2. Sequence Quality and Differential Gene Expression Profiling
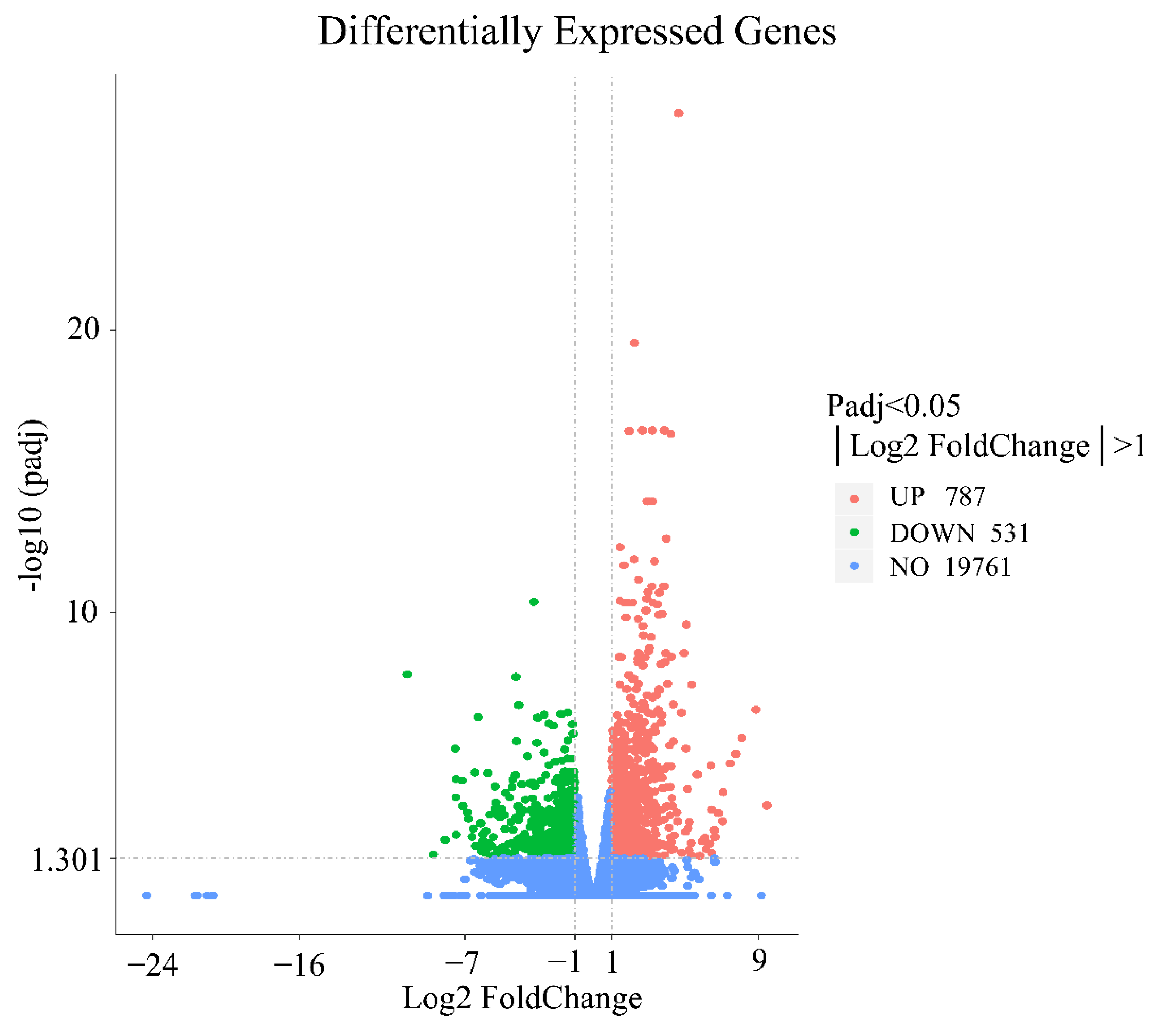
3.3. Screening of DEGs
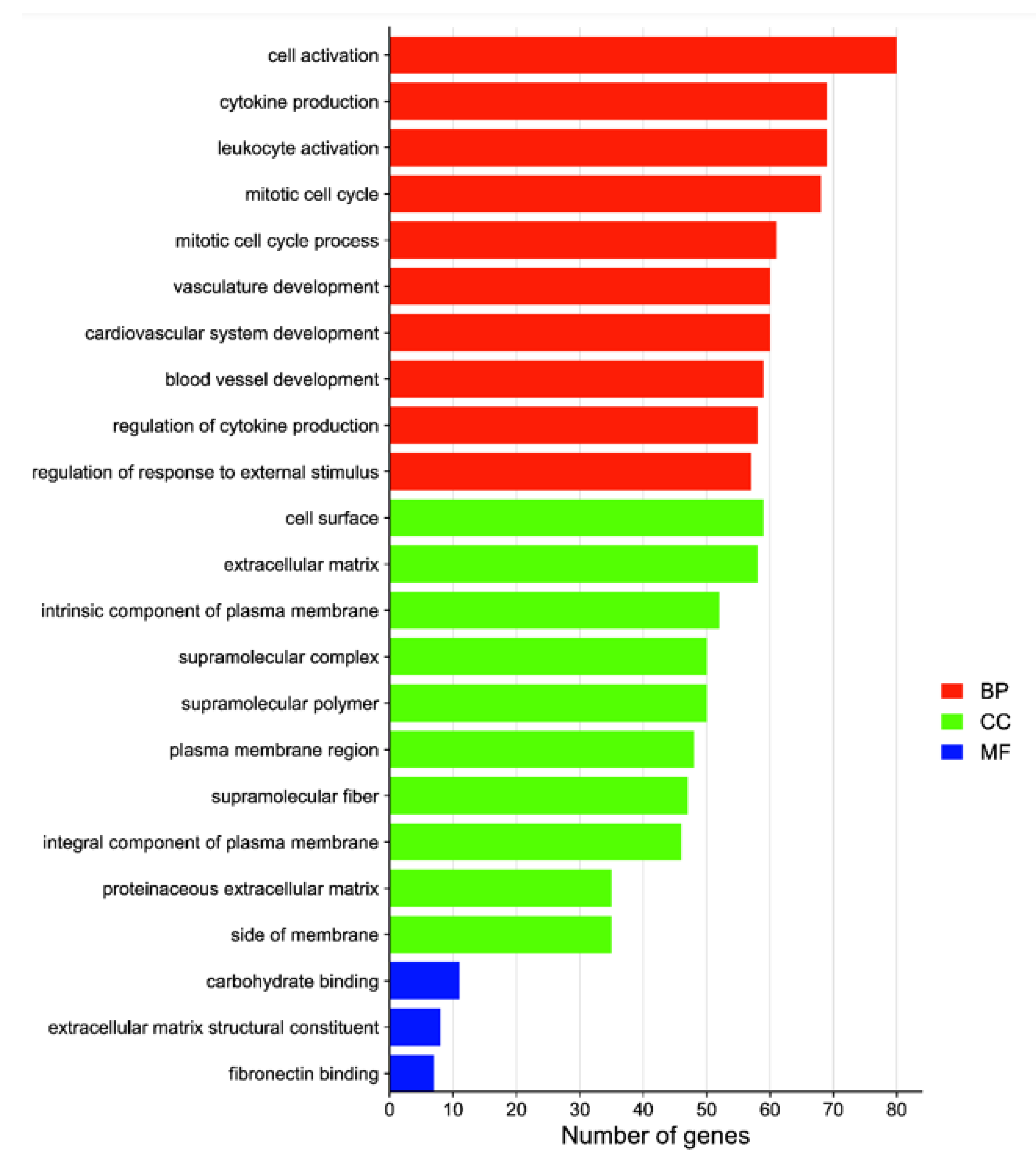
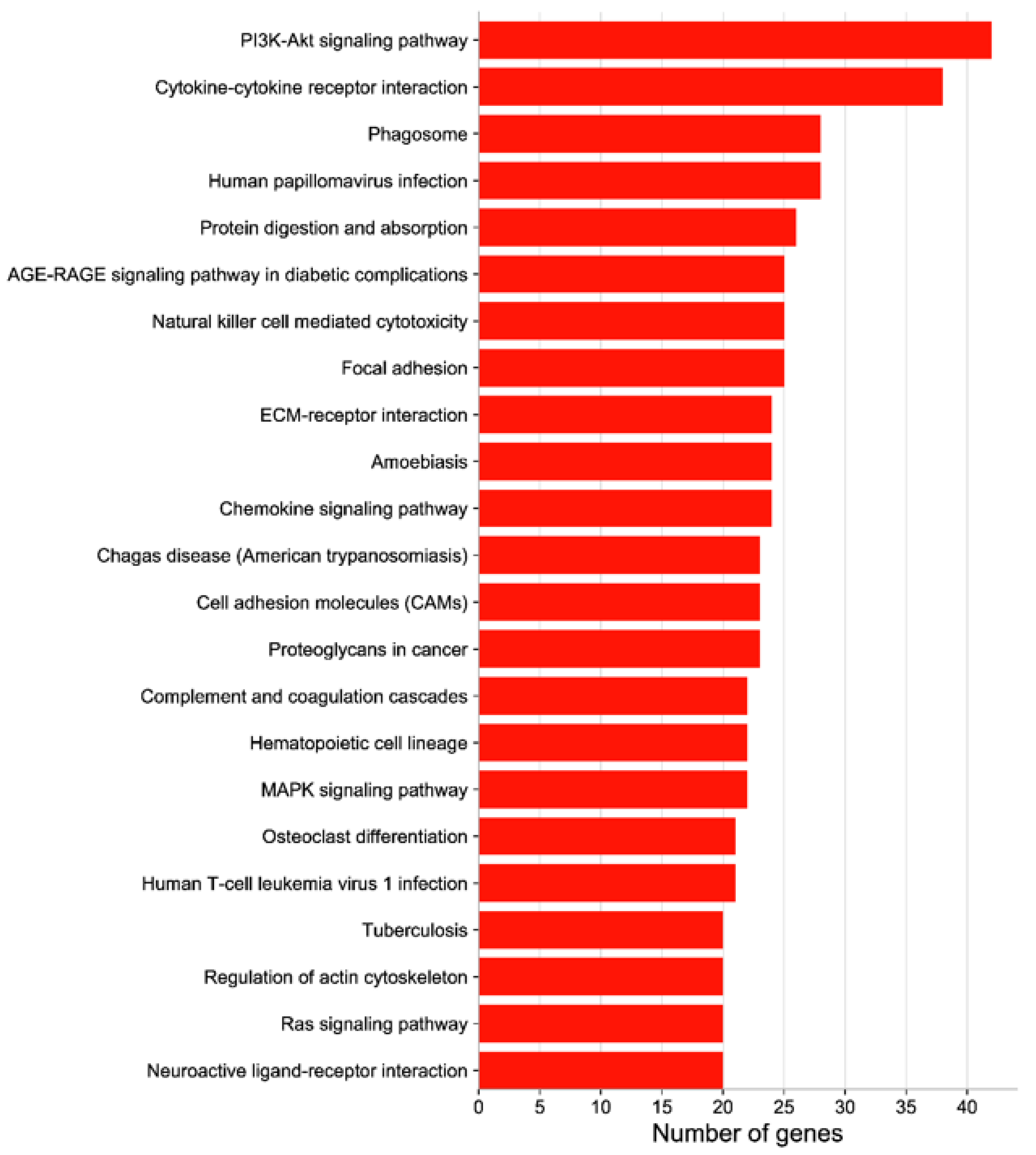
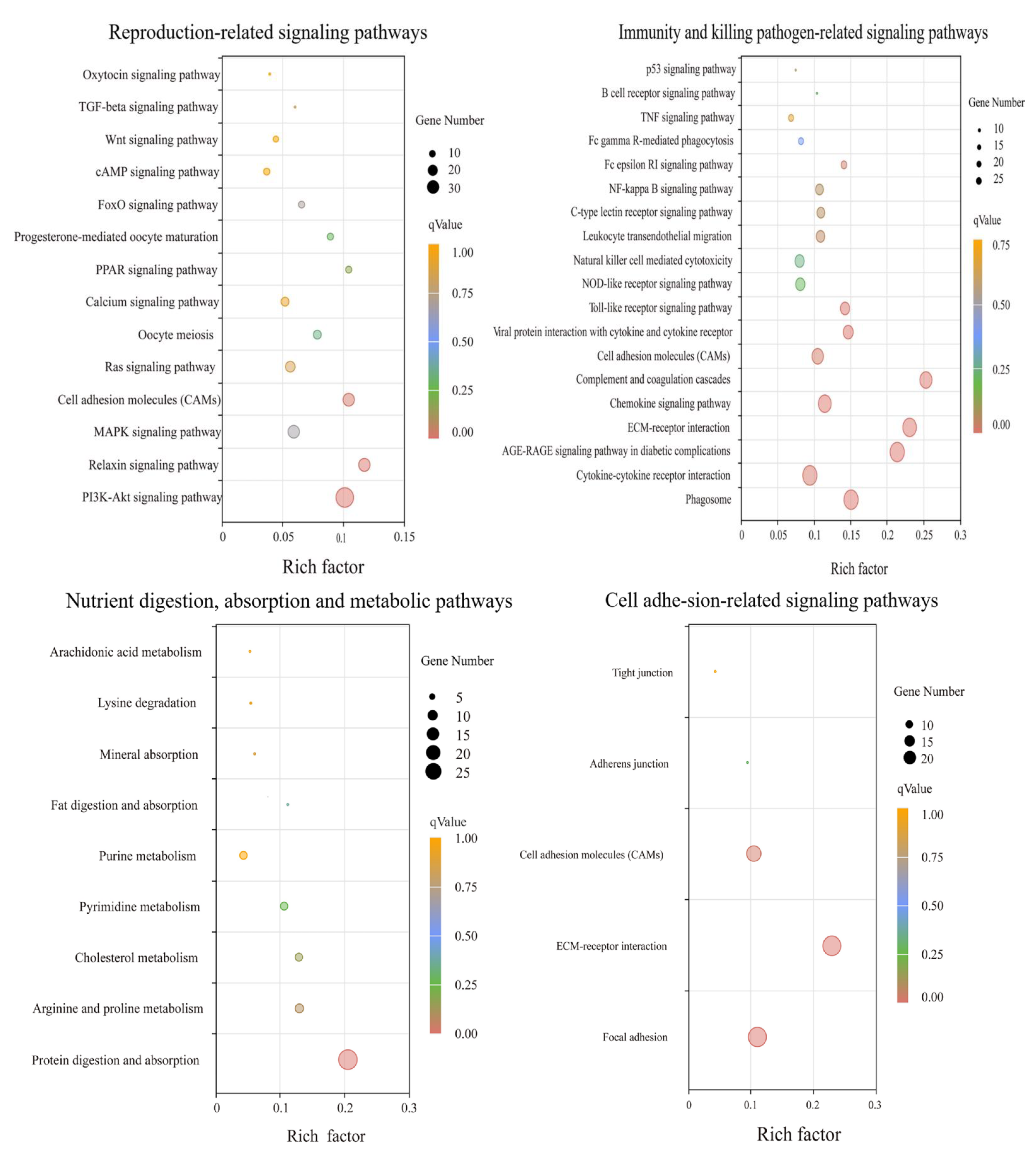
3.4. GO Function and KEGG Enrichment Analyses
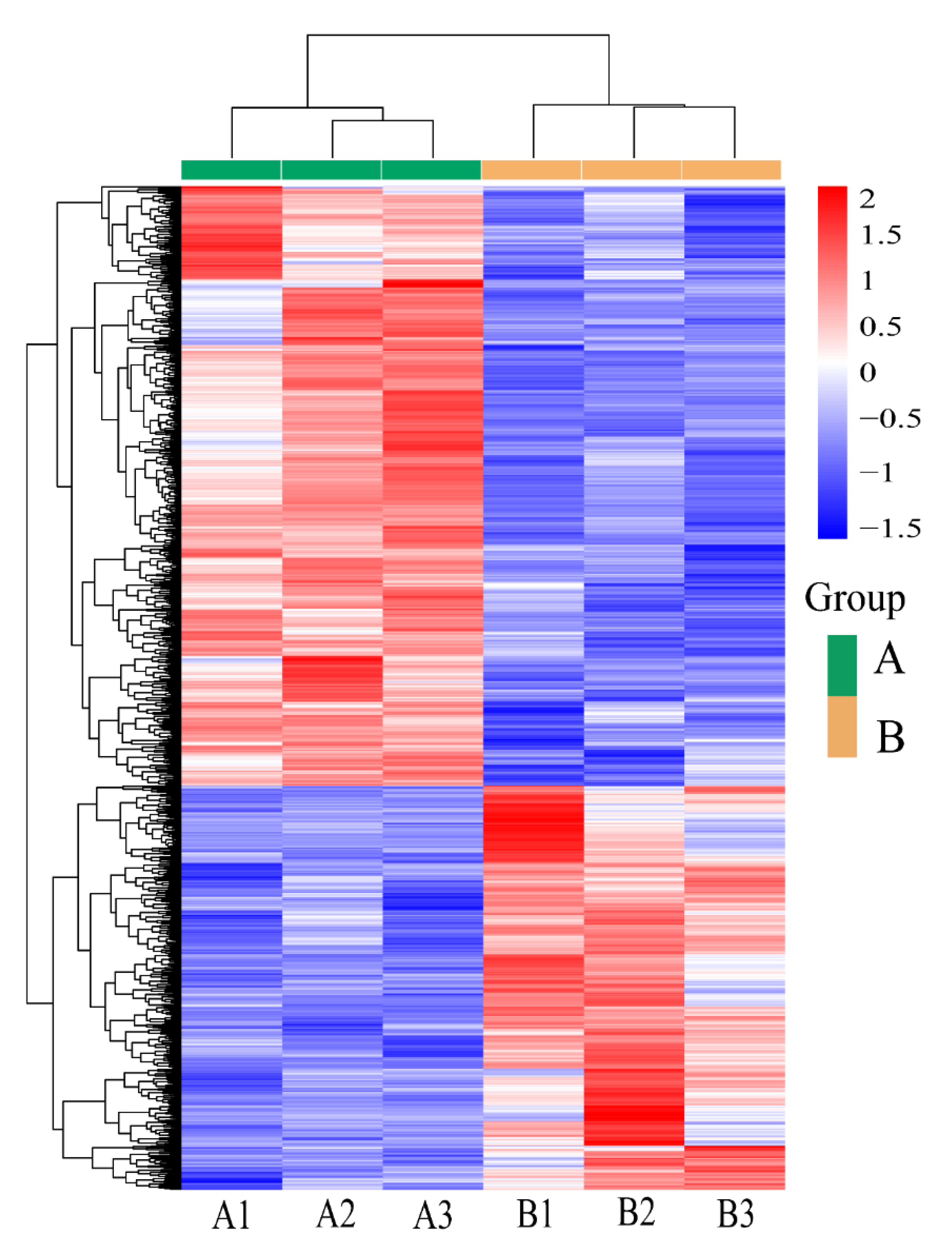
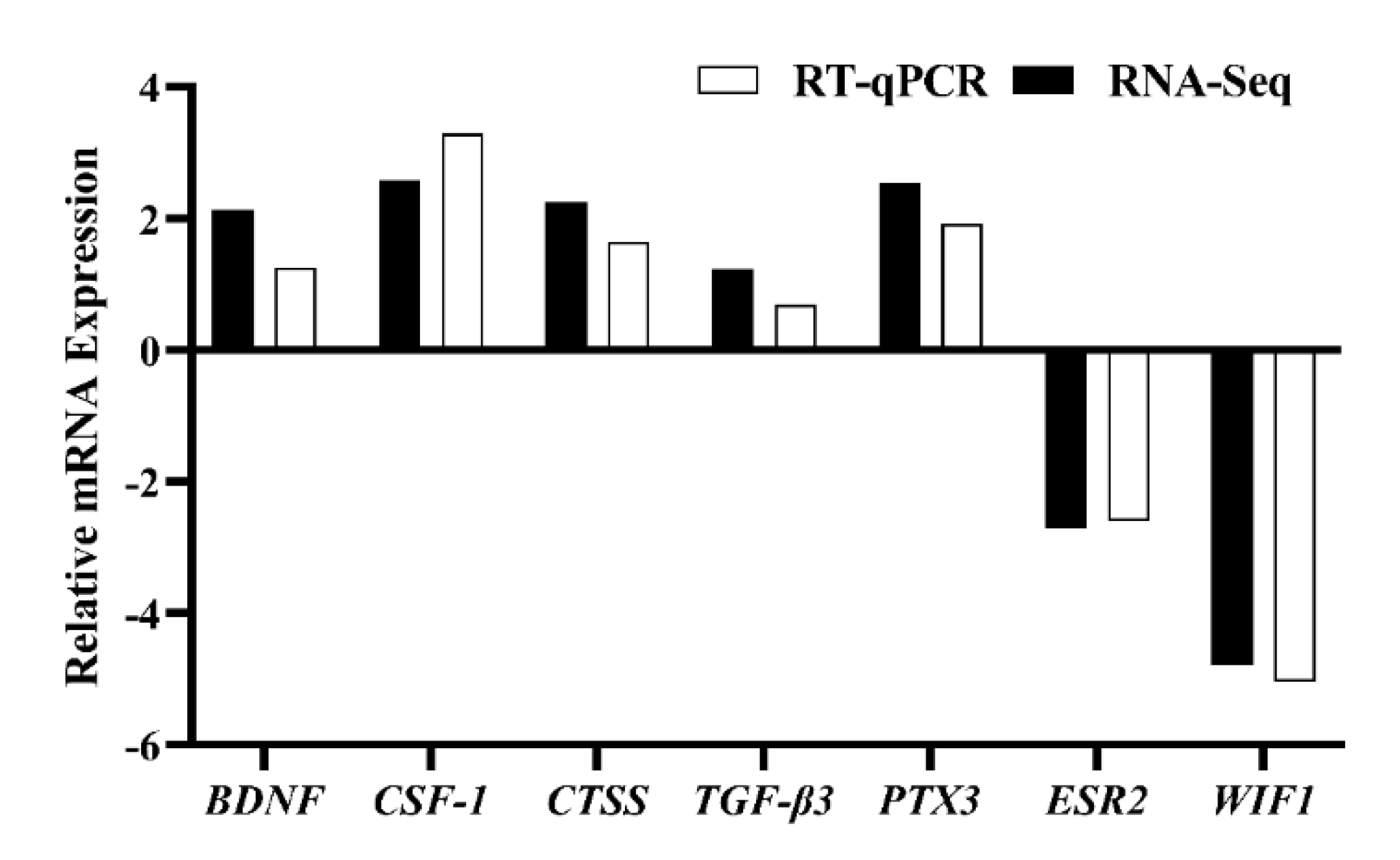
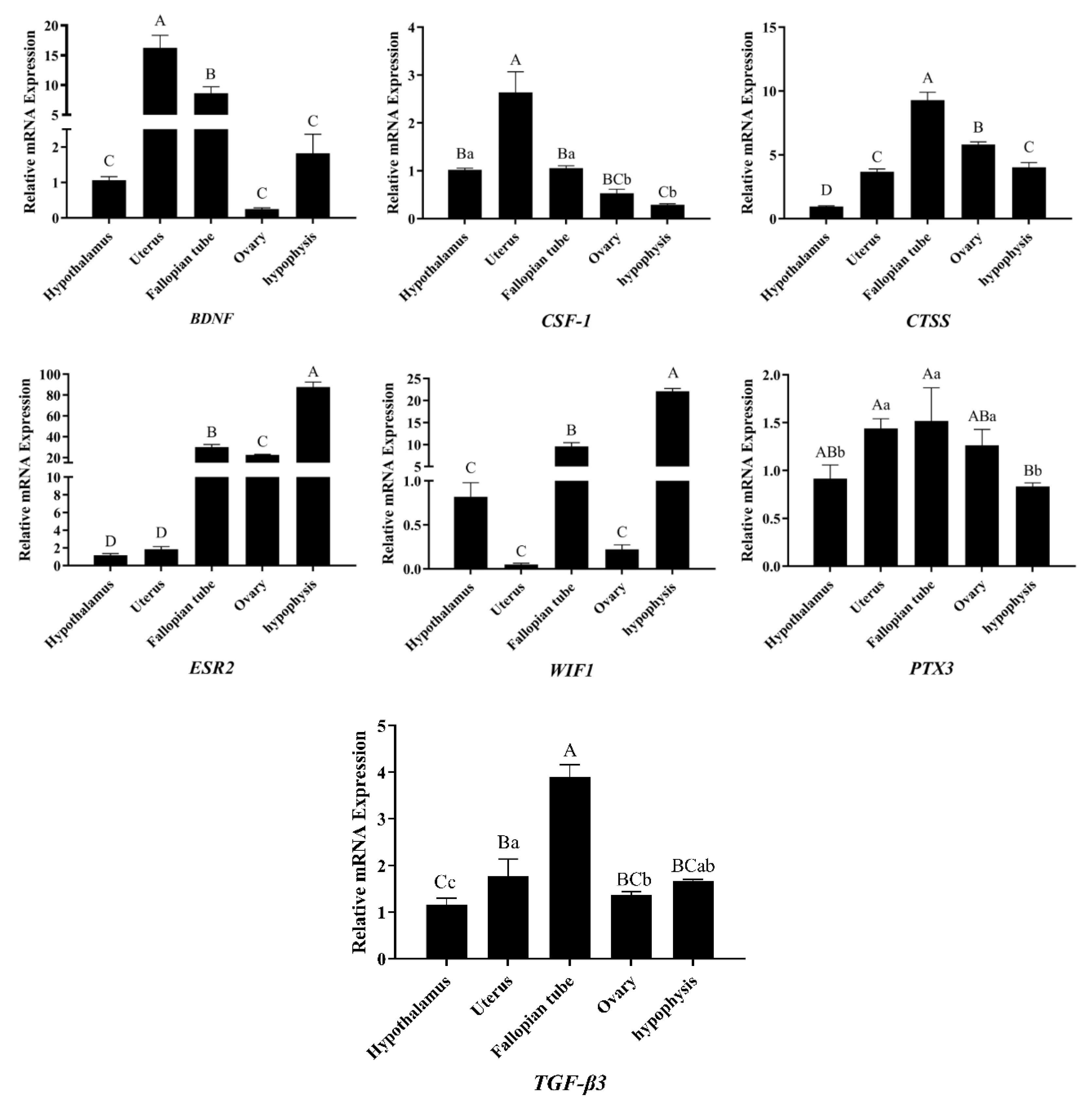
3.5. Analysis of DEGs Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jonker, F.H. Fetal death: Comparative aspects in large domestic animals. Anim. Reprod. Sci. 2004, 82–83, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Spencer, T.E.; Wu, G. Mechanisms for the establishment and maintenance of pregnancy: Synergies from scientific collaborations. Biol. Reprod. 2018, 99, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.A.; Burghardt, R.C. Cellular mechanisms of implantation in domestic farm animals. Semin. Cell Dev. Biol. 2000, 11, 93–104. [Google Scholar] [CrossRef]
- Johns, D.N.; Lucas, C.G.; Pfeiffer, C.A.; Chen, P.R.; Meyer, A.E.; Perry, S.D.; Spate, L.D.; Cecil, R.F.; Fudge, M.A.; Samuel, M.S.; et al. Conceptus interferon gamma is essential for establishment of pregnancy in the pig. Biol. Reprod. 2021, 105, 1577–1590. [Google Scholar] [CrossRef]
- Hussain, T.; Murtaza, G.; Metwally, E.; Kalhoro, D.H.; Kalhoro, M.S.; Rahu, B.A.; Sahito, R.; Yin, Y.; Yang, H.; Chughtai, M.I.; et al. The role of oxidative stress and antioxidant balance in pregnancy. Mediators Inflamm. 2021, 2021, 9962860. [Google Scholar] [CrossRef]
- Maltepe, E.; Fisher, S.J. Placenta: The forgotten organ. Annu. Rev. Cell Dev. Biol. 2015, 31, 523–552. [Google Scholar] [CrossRef]
- Mullen, M.P.; Elia, G.; Hilliard, M.; Parr, M.H.; Diskin, M.G.; Evans, A.C.; Crowe, M.A. Proteomic characterization of histotroph during the preimplantation phase of the estrous cycle in cattle. J. Proteome Res. 2012, 11, 3004–3018. [Google Scholar] [CrossRef]
- Bazer, F.W.; Kim, J.; Ka, H.; Johnson, G.A.; Wu, G.; Song, G. Select nutrients in the uterine lumen of sheep and pigs affect conceptus development. J. Reprod. Dev. 2012, 58, 180–188. [Google Scholar] [CrossRef]
- Cetin, I.; Berti, C.; Calabrese, S. Role of micronutrients in the periconceptional period. Hum. Reprod. Update 2010, 16, 80–95. [Google Scholar] [CrossRef]
- Zheng, P.; Qin, X.; Feng, R.; Li, Q.; Huang, F.; Li, Y.; Zhao, Q.; Huang, H. Alleviative effect of melatonin on the decrease of uterine receptivity caused by blood ammonia through ROS/NF-κB pathway in dairy cow. Ecotoxicol. Environ. Saf. 2022, 231, 113166. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, B.D.; Casey, S.J.; Taupier, R. N-acetyl-l-cysteine supplementation improves boar spermatozoa characteristics and subsequent fertilization and embryonic development. Reprod. Domest. Anim. 2012, 47, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, B.D.; Knight, J.W. Effects of N-acetyl-cysteine and N-acetyl-cysteine-amide supplementation on in vitro matured porcine oocytes. Reprod. Domest. Anim. 2010, 45, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.S.; Jang, H.; Park, M.R.; Oh, K.B.; Lee, H.; Hwang, S.; Xu, L.J.; Hwang, I.S.; Lee, J.W. N-acetyl-L-cysteine improves the developmental competence of bovine oocytes and embryos cultured in vitro by attenuating oxidative damage and apoptosis. Antioxidants 2021, 10, 860. [Google Scholar] [CrossRef] [PubMed]
- Rahil, J.; Kazem, P.; Hayati, R.N.; Hossein, N.M. Effects of N-acetyl-cysteine supplementation on sperm quality, chromatin integrity and level of oxidative stress in infertile men. Reprod. Biol. Endocrin. 2019, 17, 24. [Google Scholar] [CrossRef]
- Helal, M.A. The effects of N-acetyl-L-cysteine on the female reproductive performance and nephrotoxicity in rats. Ren. Fail. 2016, 38, 311–320. [Google Scholar] [CrossRef]
- Bhardwaj, J.K.; Saraf, P. N-acetyl cysteine-mediated effective attenuation of methoxychlor-induced granulosa cell apoptosis by counteracting reactive oxygen species generation in caprine ovary. Environ. Toxicol. 2017, 32, 156–166. [Google Scholar] [CrossRef]
- Wei, G.; Jin-Xiao, L.; Chi, M.; Jing-Yin, D.; Qiu, Y. The protective effect of N-Acetylcysteine on ionizing radiation induced ovarian failure and loss of ovarian reserve in female mouse. Biomed Res. Int. 2017, 2017, 4176170. [Google Scholar] [CrossRef]
- Witte, T.S.; Melkus, E.; Walter, I.; Senge, B.; Schwab, S.; Aurich, C.; Heuwieser, W. Effects of oral treatment with N-acetylcysteine on the viscosity of intrauterine mucus and endometrial function in estrous mares. Theriogenology 2012, 78, 1199–1208. [Google Scholar] [CrossRef]
- Rafiee, B.; Karbalay-Doust, S.; Tabei, S.; Azarpira, N.; Alaee, S.; Lohrasbi, P.; Bahmanpour, S. Effects of N-acetylcysteine and metformin treatment on the stereopathological characteristics of uterus and ovary. Eur. J. Transl. Myol. 2022, 32, 10409. [Google Scholar] [CrossRef]
- Hu, M.; Zhang, Y.; Ma, S.; Li, J.; Wang, X.; Liang, M.; Sferruzzi-Perri, A.N.; Wu, X.; Ma, H.; Brännström, M.; et al. Suppression of uterine and placental ferroptosis by N-acetylcysteine in a rat model of polycystic ovary syndrome. Mol. Hum. Reprod. 2021, 27, gaab067. [Google Scholar] [CrossRef]
- Omid, K.; Amirali, S.; Ahmad, K. N-Acetyl cysteine improves performance, reproduction, antioxidant status, immunity and maternal antibody transmission in breeder Japanese quail under heat stress condition. Livest. Sci. 2018, 217, 55–64. [Google Scholar] [CrossRef]
- Luo, J.; Ao, Z.; Duan, Z.; Ao, Y.; Wei, S.; Chen, W.; Chen, X. Effects of N-Acetylcysteine on the reproductive performance, oxidative stress and RNA sequencing of Nubian goats. Vet. Med. Sci. 2021, 7, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ying, Y.; Wang, S.; Li, J.; Xu, J.; Lv, P.; Chen, J.; Zhou, C.; Liu, Y.; Wu, Y.; et al. The effects and mechanisms of GM-CSF on endometrial regeneration. Cytokine 2020, 125, 154850. [Google Scholar] [CrossRef] [PubMed]
- Camargo-Díaz, F.; García, V.; Ocampo-Bárcenas, A.; González-Marquez, H.; López-Bayghen, E. Colony stimulating factor-1 and leukemia inhibitor factor expression from current-cycle cannula isolated endometrial cells are associated with increased endometrial receptivity and pregnancy. BMC Women’s Health 2017, 17, 63. [Google Scholar] [CrossRef] [PubMed]
- Chow, R.; Wessels, J.M.; Foster, W.G. Brain-derived neurotrophic factor (BDNF) expression and function in the mammalian reproductive Tract. Hum. Reprod. Update 2020, 26, 545–564. [Google Scholar] [CrossRef]
- Antonson, P.; Apolinário, L.M.; Shamekh, M.M.; Humire, P.; Poutanen, M.; Ohlsson, C.; Nalvarte, I.; Gustafsson, J.Å. Generation of an all-exon Esr2 deleted mouse line: Effects on fertility. Biochem. Biophys. Res. Commun. 2020, 529, 231–237. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Yu, W.; Sheng, X.; Zhang, H.; Han, Y.; Yuan, Z.; Weng, Q. Seasonal expressions of androgen receptor, estrogen receptors and cytochrome P450 aromatase in the uteri of the wild Daurian ground squirrels (Spermophilus dauricus). Eur. J. Histochem. 2018, 62, 2889. [Google Scholar] [CrossRef]
- Shooner, C.; Caron, P.L.; Fréchette-Frigon, G.; Leblanc, V.; Déry, M.C.; Asselin, E. TGF-beta expression during rat pregnancy and activity on decidual cell survival. Reprod. Biol. Endocrinol. 2005, 3, 20. [Google Scholar] [CrossRef]
- Giannubilo, S.R.; Landi, B.; Pozzi, V.; Sartini, D.; Cecati, M.; Stortoni, P.; Corradetti, A.; Saccucci, F.; Tranquilli, A.L.; Emanuelli, M. The involvement of inflammatory cytokines in the pathogenesis of recurrent miscarriage. Cytokine 2012, 58, 50–56. [Google Scholar] [CrossRef]
- Bottazzi, B.; Bastone, A.; Doni, A.; Garlanda, C.; Valentino, S.; Deban, L.; Maina, V.; Cotena, A.; Moalli, F.; Vago, L.; et al. The long pentraxin PTX3 as a link among innate immunity, inflammation, and female fertility. J. Leukoc. Biol. 2006, 79, 909–912. [Google Scholar] [CrossRef]
- Song, G.; Bazer, F.W.; Spencer, T.E. Differential expression of cathepsins and cystatin C in ovine uteroplacental tissues. Placenta 2007, 28, 1091–1098. [Google Scholar] [CrossRef]
- Zhang, L.; Li, W.; Song, W.; Ran, Y.; Yuan, Y.; Jia, L.; Liu, L.; Li, Y.; Cui, S.; Zhang, Z. Detection of WNT2B, WIF1 and β-catenin expression in preeclampsia by placenta tissue microarray. Clin. Chim. Acta 2018, 487, 179–185. [Google Scholar] [CrossRef]
- Cai, H.F.; Chen, Z.; Luo, W.X. Associations between polymorphisms of the GFI1B gene and growth traits of indigenous Chinese goats. Genet. Mol. Res. 2014, 13, 872–880. [Google Scholar] [CrossRef]
- Tian, X.Z.; Li, J.X.; Luo, Q.Y.; Wang, X.; Xiao, M.M.; Zhou, D.; Lu, Q.; Chen, X. Effect of supplementation with Selenium-Yeast on muscle antioxidant activity, meat quality, fatty acids and amino acids in goats. Front. Vet. Sci. 2021, 8, 813672. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Erratum: Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 888. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Robertson, S.A.; Care, A.S.; Moldenhauer, L.M. Regulatory T cells in embryo implantation and the immune response to pregnancy. J. Clin. Investig. 2018, 128, 4224–4235. [Google Scholar] [CrossRef]
- Ashary, N.; Tiwari, A.; Modi, D. Embryo implantation: War in times of love. Endocrinology 2018, 159, 1188–1198. [Google Scholar] [CrossRef] [Green Version]
- Rota, C.; Bergamini, S.; Daneri, F.; Tomasi, A.; Virgili, F.; Iannone, A. N-Acetylcysteine negatively modulates nitric oxide production in endotoxin-treated rats through inhibition of NF-kappaB activation. Antioxid. Redox Signal. 2002, 4, 221–226. [Google Scholar] [CrossRef]
- Schust, D.J.; Bonney, E.A.; Sugimoto, J.; Ezashi, T.; Roberts, R.M.; Choi, S.; Zhou, J. The immunology of syncytialized trophoblast. Int. J. Mol. Sci. 2021, 22, 1767. [Google Scholar] [CrossRef] [PubMed]
- Mei, C.; Yang, W.; Wei, X.; Wu, K.; Huang, D. The unique microbiome and innate immunity during pregnancy. Front. Immunol. 2019, 10, 2886. [Google Scholar] [CrossRef] [PubMed]
- Mou, D.; Ding, D.; Li, S.; Yan, H.; Qin, B.; Li, Z.; Zhao, L.; Che, L.; Fang, Z.; Xu, S.; et al. Effect of maternal organic selenium supplementation during pregnancy on sow reproductive performance and long-term effect on their progeny. J. Anim. Sci. 2020, 98, a366. [Google Scholar] [CrossRef] [PubMed]
- Mou, D.; Ding, D.; Yan, H.; Qin, B.; Dong, Y.; Li, Z.; Che, L.; Fang, Z.; Xu, S.; Lin, Y.; et al. Maternal supplementation of organic selenium during gestation improves sows and offspring antioxidant capacity and inflammatory status and promotes embryo survival. Food Funct. 2020, 11, 7748–7761. [Google Scholar] [CrossRef]
- Habbeddine, M.; Verbeke, P.; Karaz, S.; Bobé, P.; Kanellopoulos-Langevin, C. Leukocyte population dynamics and detection of IL-9 as a major cytokine at the mouse fetal-maternal interface. PLoS ONE 2014, 9, e107267. [Google Scholar] [CrossRef]
- Yockey, L.J.; Iwasaki, A. Interferons and proinflammatory cytokines in pregnancy and fetal development. Immunity 2018, 49, 397–412. [Google Scholar] [CrossRef]
- Karizbodagh, M.P.; Rashidi, B.; Sahebkar, A.; Masoudifar, A.; Mirzaei, H. Implantation window and angiogenesis. J. Cell Biochem. 2017, 118, 4141–4151. [Google Scholar] [CrossRef]
- Samborski, A.; Graf, A.; Krebs, S.; Kessler, B.; Reichenbach, M.; Reichenbach, H.D.; Ulbrich, S.E.; Bauersachs, S. Transcriptome changes in the porcine endometrium during the preattachment phase. Biol. Reprod. 2013, 89, 134. [Google Scholar] [CrossRef]
- Li, A.; Li, S.; Zhang, C.; Fang, Z.; Sun, Y.; Peng, Y.; Wang, X.; Zhang, M. FPR2 serves a role in recurrent spontaneous abortion by regulating trophoblast function via the PI3K/AKT signaling pathway. Mol. Med. Rep. 2021, 24, 838. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Y.; Yu, Q. The PI3K/Akt signaling pathway exerts effects on the implantation of mouse embryos by regulating the expression of RhoA. Int. J. Mol. Med. 2014, 33, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fu, L.J.; Liu, X.Q.; Hu, Z.Y.; Jiang, Y.; Gao, R.F.; Feng, Q.; Lan, X.; Geng, Y.Q.; Chen, X.M.; et al. Nm23 regulates decidualization through the PI3K-Akt-mTOR signaling pathways in mice and humans. Hum. Reprod. 2016, 31, 2339–2351. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Feng, F.; Mao, Z.; Huang, J.Z.; Liu, Y.; Li, Y.L.; Jiang, R.X. Regulation mechanism of miR-494-3p on endometrial receptivity in mice via PI3K/AKT/mTOR pathway. Gen. Physiol. Biophys. 2021, 40, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Chen, L.; Zhuang, Y.; Han, Y.; Tang, W.; Xia, F. Fibronectin 1 inhibits the apoptosis of human trophoblasts by activating the PI3K/Akt signaling pathway. Int. J. Mol. Med. 2020, 46, 1908–1922. [Google Scholar] [CrossRef]
- Chitu, V.; Stanley, E.R. Regulation of embryonic and postnatal development by the CSF-1 receptor. Curr. Top. Dev. Biol. 2017, 123, 229–275. [Google Scholar] [CrossRef]
- Armstrong, D.L.; McGowen, M.R.; Weckle, A.; Pantham, P.; Caravas, J.; Agnew, D.; Benirschke, K.; Savage-Rumbaugh, S.; Nevo, E.; Kim, C.J.; et al. The core transcriptome of mammalian placentas and the divergence of expression with placental shape. Placenta 2017, 57, 71–78. [Google Scholar] [CrossRef]
- Jang, H.; Choi, Y.; Yoo, I.; Han, J.; Kim, M.; Ka, H. Characterization of interferon α and β receptor IFNAR1 and IFNAR2 expression and regulation in the uterine endometrium during the estrous cycle and pregnancy in pigs. Theriogenology 2017, 88, 166–173. [Google Scholar] [CrossRef]
- Neirijnck, Y.; Papaioannou, M.D.; Nef, S. The Insulin/IGF system in mammalian sexual development and reproduction. Int. J. Mol. Sci. 2019, 20, 4440. [Google Scholar] [CrossRef]
- Burkin, H.R.; Rice, M.; Sarathy, A.; Thompson, S.; Singer, C.A.; Buxton, I.L. Integrin upregulation and localization to focal adhesion sites in pregnant human myometrium. Reprod. Sci. 2013, 20, 804–812. [Google Scholar] [CrossRef]
- Park, H.J.; Park, J.E.; Lee, H.; Kim, S.J.; Yun, J.I.; Kim, M.; Park, K.H.; Lee, S.T. Integrins functioning in uterine endometrial stromal and epithelial cells in estrus. Reproduction 2017, 153, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Saeed, A.M.; Saenz, D.J.M.L.; Marco, J.F.; Vicente, J.S. Oviductal and endometrial mRNA expression of implantation candidate biomarkers during early pregnancy in rabbit. Zygote 2015, 23, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Zhang, M.; Chen, X.; He, J.; Ding, Y.; Zhang, C.; Liu, X.; Wang, Y. Expression of KRAS in the endometrium of early pregnant mice and its effect during embryo implantation. Reprod. BioMed. Online 2015, 31, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Liang, J.; Feng, F.; Shi, S.; Tan, Q.; Wang, Z. MiR-183 impeded embryo implantation by regulating Hbegf and Lamc1 in mouse uterus. Theriogenology 2020, 158, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.G.; Hosoe, M.; Fujii, S.; Kanahara, H.; Sakumoto, R. Temporal expression and localization of vascular endothelial growth factor family members in the bovine uterus during peri-implantation period. Theriogenology 2019, 133, 56–64. [Google Scholar] [CrossRef]
- Zhao, X.; Du, F.; Liu, X.; Ruan, Q.; Wu, Z.; Lei, C.; Deng, Y.; Luo, C.; Jiang, J.; Shi, D.; et al. Brain-derived neurotrophic factor (BDNF) is expressed in buffalo (Bubalus bubalis) ovarian follicles and promotes oocyte maturation and early embryonic development. Theriogenology 2019, 130, 79–88. [Google Scholar] [CrossRef]
- Arceci, R.J.; Shanahan, F.; Stanley, E.R.; Pollard, J.W. Temporal expression and location of colony-stimulating factor 1 (CSF-1) and its receptor in the female reproductive tract are consistent with CSF-1-regulated placental development. Proc. Natl. Acad. Sci. USA 1989, 86, 8818–8822. [Google Scholar] [CrossRef]
- Ding, J.; Yang, C.; Zhang, Y.; Wang, J.; Zhang, S.; Guo, D.; Yin, T.; Yang, J. M2 macrophage-derived G-CSF promotes trophoblasts EMT, invasion and migration via activating PI3K/Akt/Erk1/2 pathway to mediate normal pregnancy. J. Cell Mol. Med. 2021, 25, 2136–2147. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, Y.; Lu, J.; Wang, Q.; Wang, S.; Cao, Y.; Wang, H.; Duan, E. Embryo–uterine cross-talk during implantation: The role of Wnt signaling. Mol. Hum. Reprod. 2009, 15, 215–221. [Google Scholar] [CrossRef]
- Poggi, L.; Casarosa, S.; Carl, M. An Eye on the Wnt Inhibitory Factor Wif1. Front. Cell Dev. Biol. 2018, 6, 167. [Google Scholar] [CrossRef]
- Tan, J.; Paria, B.C.; Dey, S.K.; Das, S.K. Differential uterine expression of estrogen and progesterone receptors correlates with uterine preparation for implantation and decidualization in the mouse1. Endocrinology 1999, 140, 5310–5321. [Google Scholar] [CrossRef]
- Hummitzsch, K.; Hatzirodos, N.; Irving-Rodgers, H.F.; Hartanti, M.D.; Perry, V.; Anderson, R.A.; Rodgers, R.J. Morphometric analyses and gene expression related to germ cells, gonadal ridge epithelial-like cells and granulosa cells during development of the bovine fetal ovary. PLoS ONE 2019, 14, e214130. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Winuthayanon, W. Oviduct: Roles in fertilization and early embryo development. J. Endocrinol. 2017, 232, R1–R26. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients % | Content | Chemical Composition % | Content |
|---|---|---|---|
| Corn silage | 50 | DM | 60.13 |
| Chinese wildrye | 20 | Metabolic energy (MJ/kg DM) | 12.51 |
| Corn | 15 | Crude protein, DM | 13.42 |
| Soybean meal | 8.02 | Organic matter | 86.38 |
| Wheat bran | 4.98 | NDF | 38.67 |
| Calcium bicarbonate | 0.5 | ADF | 31.07 |
| Sodium chloride | 0.5 | Ca | 0.68 |
| Premix * | 1 | P | 0.49 |
| Total | 100 |
| Gene Name | Primer Sequences (5′→3′) | GenBank ID | Product Size/bp | Tm/°C |
|---|---|---|---|---|
| BDNF | F: GTCCTTGAAAAAGTCCCCG R: CTATCCGAATGAACCGCCA | XM_005690025.2 | 205 | 61 |
| CSF-1 | F: AGGTGTCGGAGAACTGTAGC R: TTGGGGGTGTTGTCTTTGAA | XM_013962557.2 | 214 | 56 |
| CTSS | F: ACTGGAGAGAGAAGGGGTGT R: ATGGCTTTGTAGGGATAGGA | XM_005677657.3 | 268 | 59.4 |
| PTX3 | F: CTGGTCGCTGATGCTGT R: GCCATTCTTTTCTTGCC | XM_018048475.1 | 183 | 59.4 |
| TGF-β3 | F: GCCAAGCAGCGGTAT R: GCAAGAGCCATTCACG | XM_005686141.3 | 100 | 57.9 |
| WIF1 | F: GTGGCAGCATTTGAAGTGAAC R: ATCCATCAGGACATTCGCAG | XM_005680217.2 | 172 | 56 |
| ESR2 | F: GACAGACCACAAGCCCAAA R: GGCACAACTGCTCCCACTA | NM_001285688.1 | 191 | 60 |
| β-actin | F: TGATATTGCTGCGCTCGTGGT R: GTCAGGATGCCTCTCTTGCTC | XM_018039831.1 | 189 | All |
| Items | Treatment Group | Control Group |
|---|---|---|
| No. of test does | 30 | 30 |
| No. of pregnant goats | 23 | 21 |
| No. of total kids | 54 | 45 |
| No. of kids in slaughtered does | 7 | 5 |
| No. of average kids | 2.35 ± 0.71 | 2.14 ± 0.73 |
| Conception rate | 76.67% | 70% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, K.; Chen, X.; Guo, W.; Zhou, Z.; Zhang, Y.; Ji, T.; Yang, P.; Tian, X.; Wang, W.; Zou, Y. Effects of N Acetylcysteine on the Expression of Genes Associated with Reproductive Performance in the Goat Uterus during Early Gestation. Animals 2022, 12, 2431. https://doi.org/10.3390/ani12182431
Fu K, Chen X, Guo W, Zhou Z, Zhang Y, Ji T, Yang P, Tian X, Wang W, Zou Y. Effects of N Acetylcysteine on the Expression of Genes Associated with Reproductive Performance in the Goat Uterus during Early Gestation. Animals. 2022; 12(18):2431. https://doi.org/10.3390/ani12182431
Chicago/Turabian StyleFu, Kaibin, Xiang Chen, Wei Guo, Zhinan Zhou, Yan Zhang, Taotao Ji, Peifang Yang, Xingzhou Tian, Weiwei Wang, and Yue Zou. 2022. "Effects of N Acetylcysteine on the Expression of Genes Associated with Reproductive Performance in the Goat Uterus during Early Gestation" Animals 12, no. 18: 2431. https://doi.org/10.3390/ani12182431
APA StyleFu, K., Chen, X., Guo, W., Zhou, Z., Zhang, Y., Ji, T., Yang, P., Tian, X., Wang, W., & Zou, Y. (2022). Effects of N Acetylcysteine on the Expression of Genes Associated with Reproductive Performance in the Goat Uterus during Early Gestation. Animals, 12(18), 2431. https://doi.org/10.3390/ani12182431