
Effects of Chromium-L-Methionine in Combination with a Zinc Amino Acid Complex or Selenomethionine on Growth Performance, Intestinal Morphology, and Antioxidative Enzymes in Red Tilapia Oreochromis spp.


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Diets Description
2.3. Experimental Condition
2.4. Data Collection
2.4.1. Growth Performance
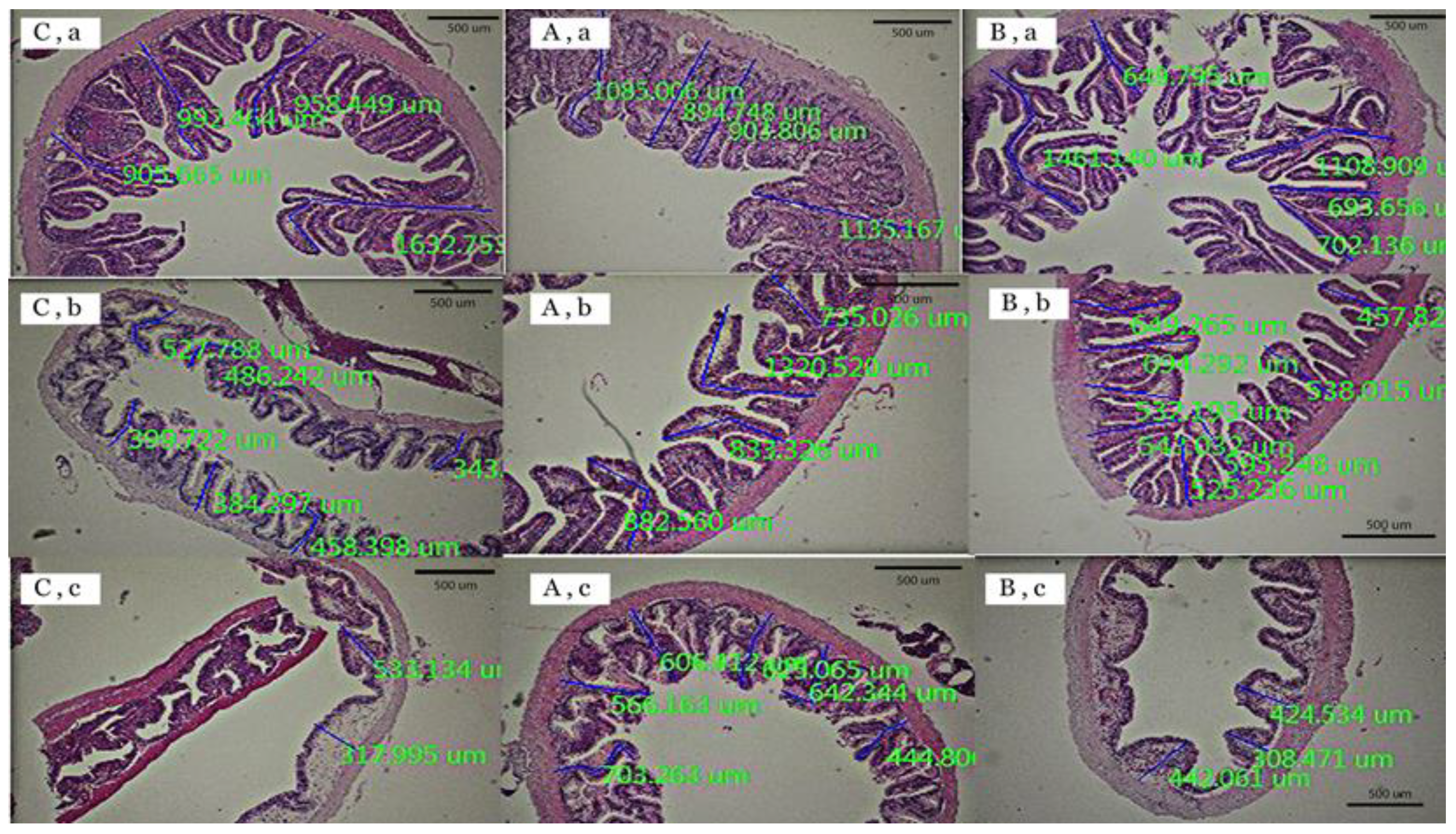
2.4.2. Measurement of Intestinal Villus Height
2.4.3. Hematological Parameters and Antioxidative Enzymes
2.5. Statistical Analysis
3. Results and Discussion
3.1. Growth Performance
3.2. Measurement of Villus Height
3.3. Hematological Parameters and Immune Response
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Department of Fisheries. Annual Thailand Export-Import of Aquatic Animals/Products. 2019. Available online: http://www4.fisheries.go.th/dof_en (accessed on 15 February 2021).
- Hilsdorf, A.W.; Penman, D.J.; Farias, E.C.; McAndrew, B. Melanophore appearance in wild and red tilapia embryos. Pigment Cell Res. 2002, 15, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Cnaani, A.; Hulata, G. Improving salinity tolerance in tilapias: Past experience and future prospects. Isr. J. Aquac. Bamidgeh 2011, 63, 20590. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020; Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Hardy, R.W. Utilization of plant proteins in fish diets: Effects of global demand and supplies of fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Connelly, P. Nutritional advantages and disadvantages of dietary phytates: A literature review Part 2. J. Aust. Tradit.-Med. Soc. 2011, 17, 16–20. [Google Scholar]
- Olsen, R.L.; Hasan, M.R. A limited supply of fishmeal: Impact on future increases in global aquaculture production. Trends Food Sci. Technol. 2012, 27, 120–128. [Google Scholar] [CrossRef]
- Lall, S.P.; Kaushik, S.J. Nutrition and metabolism of minerals in fish. Animals 2021, 11, 2711. [Google Scholar] [CrossRef]
- Vincent, J.B. The biochemistry of chromium. J. Nutr. 2000, 130, 715–718. [Google Scholar] [CrossRef]
- Le, K.T.; Fotedar, R. Bioavailability of selenium from different dietary sources in yellowtail kingfish (Seriola lalandi). Aquaculture 2014, 420, 457–462. [Google Scholar] [CrossRef]
- Vincent, J.B. New evidence against chromium as an essential trace element. J. Nutr. 2017, 147, 2212–2219. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on dietary reference values for chromium. EFSA J. 2014, 12, 1–27. [Google Scholar]
- Mehrim, A. Physiological, biochemical and histometric responses of Nile tilapia (Oreochromis niloticus L.) by dietary organic chromium (chromium picolinate) supplementation. J. Adv. Res. 2014, 5, 303–310. [Google Scholar] [CrossRef]
- Mehrim, A. Effect of Dietary Chromium Picolinate Supplementation on Growth Performance, Carcass Composition and Organs Indices of Nile Tilapia (Oreochromis niloticus L.) Fingerlings. J. Fish. Aquat. Sci. 2012, 7, 224–232. [Google Scholar] [CrossRef]
- Gatta, P.P.; Thompson, K.D.; Smullen, R.; Piva, A.; Testi, S.; Adams, A. Dietary organic chromium supplementation and its effect on the immune response of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2001, 11, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Sissener, N.H.; Julshamn, K.; Espe, M.; Lunestad, B.T.; Hemre, G.I.; Waagbø, R.; Måge, A. Surveillance of selected nutrients, additives and undesirables in commercial Norwegian fish feeds in the years 2000–2010. Aquac. Nutr. 2013, 19, 555–572. [Google Scholar] [CrossRef]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef]
- Glover, C.N.; Hogstrand, C. Amino acid modulation of in vivo intestinal zinc uptake in freshwater rainbow trout. J. Exp. Biol. 2002, 205, 151–158. [Google Scholar] [CrossRef]
- Prabhu, A.J.P.; Schrama, J.W.; Kaushik, S.J. Mineral requirements of fish: A systematic review. Rev. Aquac. 2016, 8, 172–219. [Google Scholar] [CrossRef]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- Arthur, J.R.; McKenzie, R.C.; Beckett, G.J. Selenium in the immune system. J. Nutr. 2003, 133, 1457–1459. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, N.N.; Shin, H.S.; Park, M.S.; Kil, G.S.; Choi, Y.C. Effects of waterborne selenium exposure on the antioxidant and immunological activity in the goldfish, Carassius auratus. Mol. Cell. Toxicol. 2013, 9, 365–373. [Google Scholar] [CrossRef]
- Dalgaard, T.S.; Briens, M.; Engberg, R.M.; Lauridsen, C. The influence of selenium and selenoproteins on immune responses of poultry and pigs. Anim. Feed Sci. Technol. 2018, 258, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Labunsky, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Queval, G.; Mhamdi, A.; Chaouch, S.; Foyer, C.H. Glutathione. Arab. Book 2011, 9, 142. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef]
- Li, G.; Xie, P.; Li, H.; Chen, J.; Hao, L.; Xiong, Q. Quantitative profiling of mRNA expression of glutathione S-transferase superfamily genes in various tissues of bighead carp (Aristichthys nobilis). J. Biochem. Mol. Toxicol. 2010, 24, 250–259. [Google Scholar] [CrossRef]
- Ammerman, C.B. Methods for estimation of mineral bioavailability. In Bioavailability of Nutrients for Animals: Amino Acids, Minerals and Vitamins; Ammerman, C.B., Baker, D.J., Lewis, A.J., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 83–94. [Google Scholar]
- Gatlin, D.M., III; Wilson, R.P. Dietary selenium requirement of fingerling channel catfish. J. Nutr. 1984, 114, 627–633. [Google Scholar] [CrossRef]
- Wang, C.; Lovell, R.T. Organic selenium sources, selenomethionine and selenoyeast, have higher bioavailability than an inorganic selenium source, sodium selenite, in diets for channel catfish (Ictalurus puncatatus). Aquaculture 1997, 152, 223–234. [Google Scholar] [CrossRef]
- Katya, K.; Lee, S.; Bharadwaj, A.S.; Browdy, C.L.; Vazquez-Anon, M.; Bai, S.C. Effects of inorganic and chelated trace mineral (Cu, Zn, Mn and Fe) premixes in marine rockfish, Sebastes schlegeli (Hilgendorf), fed diets containing phytic acid. Aquac. Res. 2017, 48, 4165–4173. [Google Scholar] [CrossRef]
- Paripatananont, T.; Lovell, R.T. Comparative net absorption of chelated and inorganic trace minerals in channel catfish Ictalurus punctatus diets. J. World Aquac. Soc. 1997, 28, 62–67. [Google Scholar] [CrossRef]
- Ashmead, H.D.; Zunino, H. Factors which affect the intestinal absorption of minerals. In The Roles of Amino Acid Chelates in Animal Nutrition; Noyes Publications: Westwood, NJ, USA, 1992; pp. 21–46. [Google Scholar]
- Shao, X.P.; Liu, W.B.; Xu, W.N.; Xia, W.; Jiang, Y.Y. Effects of dietary copper sources and levels on performance, copper status, plasma antioxidant activities and relative copper bioavailability in Carassius auratus gibelio. Aquaculture 2010, 308, 60–65. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirement of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Pirarat, N.; Pinpimai, K.; Endo, M.; Katagiri, T.; Ponpornpisit, A.; Chansue, N.; Maita, M. Modulation of intestinal morphology and immunity in nile tilapia (Oreochromis niloticus) by Lactobacillus rhamnosus GG. Res. Vet. Sci. 2011, 91, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Feldman, A.; Wolfe, D. Tissue Processing and Hematoxylin and Eosin Staining. Methods Mol. Biol. 2014, 1180, 31–43. [Google Scholar] [PubMed]
- Morris, M.W.; Davey, F.R. Basic examination of blood. In Clinical Diagnosis and Management by Laboratory Methods, 19th ed.; Henry, J.B., Ed.; WB Saunders: Philadelphia, PA, USA, 1996; pp. 549–593. [Google Scholar]
- Drabkin, D.L.; Austin, J.H. Spectrophotometric studies; Spectrophotometric constants for common hemoglobin derivatives in human, dog and rabbit blood. J. Biol. Chem. 1932, 98, 719–733. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, N.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Takahashi, Y.; Itami, T.; Konegawa, K. Enzymatic Properties of Partially Purified Lysozyme from the Skin Mucus of Carp. Nippon Suisan Gakkaishi 1986, 52, 1209–1214. [Google Scholar] [CrossRef][Green Version]
- Tsukatani, T.; Suenaga, H.; Higuchi, T.; Akao, T.; Ishiyama, M.; Ezoe, K.; Matsumoto, K. Colorimetric cell proliferation assay for microorganisms in microtiter plate using water-soluble tetrazolium salts. J. Microbiol. Methods 2008, 75, 109–116. [Google Scholar] [CrossRef]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- MacDonald, R.S. The role of zinc in growth and cell proliferation. J. Nutr. 2000, 130, 1500–1508. [Google Scholar] [CrossRef]
- Dawood, M.A.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Tantikitti, C.; Supamas, K. The Effect of Diet Supplementation with Different Zinc Sources and Levels on Growth Performance, Immune Response, Disease Resistance and Histopathology of Oreochromis spp. Available online: http://kb.psu.ac.th/psukb/handle/2010/8631 (accessed on 15 February 2020).
- Huang, F.; Jiang, M.; Wen, H.; Wu, F.; Liu, W.; Tian, J.; Yang, C. Dietary zinc requirement of adult Nile tilapia (Oreochromis niloticus) fed semi-purified diets, and effects on tissue mineral composition and antioxidant responses. Aquaculture 2015, 439, 53–59. [Google Scholar] [CrossRef]
- Akram, Z.; Fatima, M.; Shah, S.Z.H.; Afzal, M.; Hussain, S.M.; Hussain, M.; Khan, Z.I.; Akram, K. Dietary zinc requirement of Labeo rohita juveniles fed practical diets. J. Appl. Anim. Res. 2019, 47, 223–229. [Google Scholar] [CrossRef]
- Zhao, H.X.; Cao, J.M.; Liu, X.H.; Zhu, X.; Chen, S.C.; Lan, H.B.; Wang, A.L. Effect of supplemental dietary zinc sources on the growth and carbohydrate utilization of tilapia Smith 1840, Oreochromis niloticus × Oreochromis aureus. Aquac. Nutr. 2011, 17, 64–72. [Google Scholar] [CrossRef]
- Mertz, W. Chromium in human nutrition: A review. J. Nutr. 1993, 123, 626–633. [Google Scholar] [CrossRef]
- Chanda, S.; Paul, B.; Ghosh, K.; Giri, S. Dietary essentiality of trace minerals in aquaculture-A Review. Agric. Rev. 2015, 36, 100–112. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, J.; Zhu, T.; Zhang, X.; Jin, M.; Jiao, L.; Meng, F.; Figueiredo-Silva, C.; Hong, U.; Zhou, Q. Dietary chromium could improve growth, antioxidant capacity, chromium accumulation in tissues and expression of genes involved into glucose and lipid metabolism in juvenile mud crab Scylla paramamosain. Aquac. Rep. 2022, 23, 101088. [Google Scholar] [CrossRef]
- Rakhmawati, R.; Suprayudi, M.A.; Setiawati, M.; Widanarni, W.; Junior, M.Z.; Jusadi, D. Bioefficacy of dietary chromium picolinate and chromium yeast on growth performance and blood biochemical in Red Tilapia, Oreochromis niloticus (Linnaeus). Aquac. Res. 2018, 49, 839–846. [Google Scholar] [CrossRef]
- Kegley, E.; Spears, J. Immune response, glucose metabolism, and performance of stressed feeder calves fed inorganic or organic chromium. J. Anim. Sci. 1995, 73, 2721–2726. [Google Scholar]
- Liu, T.; Wen, H.; Jiang, M.; Yuan, D.; Gao, P.; Zhao, Y.; Wu, F.; Liu, W. Effect of dietary chromium picolinate on growth performance and blood parameters in grass carp fingerling (Ctenopharyngodon idellus). Fish Physiol. Biochem. 2009, 36, 565–572. [Google Scholar] [CrossRef]
- Bureau, D.P.; Kirkland, J.B.; Cho, C.Y. The effects of dietary chromium supplementation on performance, carcass yield and blood glucose of rainbow trout Oncorhynchus mykiss fed two practical diets. J. Anim. Sci. 1995, 73, 194. [Google Scholar]
- Ebrahimzadeh, S.; Farhoomand, P.; Noori, K. Effects of chromium methionine supplementation on performance, carcass traits, and the Ca and P metabolism of broiler chickens under heat-stress conditions. J. Appl. Poult. Res. 2013, 22, 382–387. [Google Scholar] [CrossRef]
- Xu, X.; Liu, L.; Long, S.-F.; Piao, X.-S.; Ward, T.L.; Ji, F. Effects of Chromium Methionine Supplementation with Different Sources of Zinc on Growth Performance, Carcass Traits, Meat Quality, Serum Metabolites, Endocrine Parameters, and the Antioxidant Status in Growing-Finishing Pigs. Biol. Trace Elem. Res. 2017, 179, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Van der Marel, M.; Pröpsting, M.; Battermann, F.; Jung-Schroers, V.; Hübner, A.; Rombout, J.; Steinhagen, D. Differences between intestinal segments and soybean meal–induced changes in intestinal mucus composition of common carp Cyprinus carpio L. Aquac. Nutr. 2014, 20, 12–24. [Google Scholar] [CrossRef]
- Ewtushik, A.; Bertolo, R.; Ball, R.O. Intestinal development of early-weaned piglets receiving diets supplemented with selected amino acids or polyamines. Can. J. Anim. Sci. 2000, 80, 653–662. [Google Scholar] [CrossRef]
- Sklan, D.; Prag, T.; Lupatsch, I. Structure and function of the small intestine of the tilapia Oreochromis niloticus × Oreochromis aureus (Teleostei, Cichlidae). Aquac. Res. 2004, 35, 350–357. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Hemre, G.I.; Mommsen, T. Carbohydrates in fish nutrition: Digestion and absorption in postlarval stages. Aquac. Nutr. 2005, 11, 103–122. [Google Scholar] [CrossRef]
- Ramos, M.A.; Batista, S.; Pires, M.A.; Silva, A.P.; Pereira, L.F.; Saavedra, M.J.; Ozório, R.O.A.; Rema, P. Dietary probiotic supplementation improves growth and the intestinal morphology of Nile tilapia. Animal 2017, 11, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Xiao, K.; Jiao, L.F.; Song, J. Effects of zinc oxide supported on zeolite on growth performance, intestinal barrier function and digestive enzyme activities of Nile tilapia. Aquac. Nutr. 2014, 20, 486–493. [Google Scholar] [CrossRef]
- Ibrahim, M.S.; El-Gendi, G.M.I.; Ahmed, A.I.; El-Haroun, E.R.; Hassaan, M.S. Nano Zinc Versus Bulk Zinc Form as Dietary Supplied: Effects on Growth, Intestinal Enzymes and Topography, and Hemato-biochemical and Oxidative Stress Biomarker in Nile Tilapia (Oreochromis niloticus Linnaeus, 1758). Biol. Trace Elem. Res. 2022, 200, 1347–1360. [Google Scholar] [CrossRef]
- Hardy, R.W.; Shearer, K.D. Effect of dietary calcium phosphate and zinc supplementation on whole body zinc concentration of rainbow trout (Salmo gairdneri). Can. J. Fish. Aquat. Sci. 1985, 42, 181–184. [Google Scholar] [CrossRef]
- Yuan, Y.; Luo, J.; Zhu, T.; Jin, M.; Jiao, L.; Sun, P.; Ward, T.L.; Ji, F.; Xu, G.; Zhou, Q. Alteration of growth performance, meat quality, antioxidant and immune capacity of juvenile Litopenaeus vannamei in response to different dietary dosage forms of zinc: Comparative advantages of zinc amino acid complex. Aquaculture 2020, 522, 735120. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Z.; Ren, Y.; Zhou, Z.; Wang, W.-X.; Huang, Y.; Shu, X. Improving Heat Resistance of Nile Tilapia (Oreochromis niloticus) by Dietary Zinc Supplementation. Aquac. Nutr. 2022, 2022, 6323789. [Google Scholar] [CrossRef]
- Subbotkina, T.A.; Subbotkin, M. Lysozyme of Cyprinidae in different climate zones. Probl. Fish. 2015, 16, 118–125. [Google Scholar] [CrossRef]
- Magnadottir, B. Innate immunity of fish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Bukharin, O.V.; Valyshev, A.V. Microbial inhibitors of lysozyme. Zh. Mikrobiol. Epidemiol. Immunobiol. 2006, 4, 8–13. [Google Scholar]
- Saurabh, S.; Sahoo, P. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Alvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant Defenses in Fish: Biotic and Abiotic Factors. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Silva, C.F. Managing Stress in Aquaculture Production with Performance Trace Minerals. Available online: https://essentialfeed.zinpro.com/2020/01/managing-stress-in-aquaculture-production-with-performance-trace-minerals/ (accessed on 13 February 2020).
- Nakano, T.; Sato, M.; Takeuchi, M. Glutathione peroxidase of fish. J. Food Sci. 1992, 57, 1116–1119. [Google Scholar] [CrossRef]
- Stenersen, J.; Øien, N. Glutathione S-transferases in earthworms (Lumbricidae). Substrate specificity, tissue and species distribution and molecular weight. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1981, 69, 243–252. [Google Scholar] [CrossRef]
- Ribas, V.; García-Ruiz, C.; Fernández-Checa, J.C. Glutathione and mitochondria. Front. Pharmacol. 2014, 5, 151. [Google Scholar] [CrossRef]
- Nagai, T.; Inada, J.; Hamada, M.; Kai, N.; Tanoue, Y.; Kaminishi, Y.; Nakagawa, H.; Fujiki, K.; Nakao, M.; Yano, T. Distribution of glutathione peroxidase activity in fish. Fish. Sci. 1999, 65, 665–666. [Google Scholar] [CrossRef]
- Poston, H.A.; Gerald, F.C.; Leibovitz, L. Vitamin E and Selenium Interrelations in the Diet of Atlantic Salmon (Salmo salar), Gross, Histological and Biochemical Deficiency Signs. J. Nutr. 1976, 106, 892–904. [Google Scholar] [CrossRef]
- Lushchak, V.I. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef]
- Bell, J.G.; Cowey, C.; Adron, J.; Shank, A. Some effects of vitamin E and selenium deprivation on tissue enzyme levels and indices of tissue peroxidation in rainbow trout (Salmo gairdneri). Br. J. Nutr. 1985, 53, 149–157. [Google Scholar] [CrossRef]
- Bell, J.G.; Pirie, B.; Adron, J.; Cowey, C. Some effects of selenium deficiency on glutathione peroxidase (EC 1.11.1.9) activity and tissue pathology in rainbow trout (Salmo gairdneri). Br. J. Nutr. 1986, 55, 305–311. [Google Scholar] [CrossRef]
- Bell, J.G.; Cowey, C.; Adron, J.; Pirie, B. Some effects of selenium deficiency on enzyme activities and indices of tissue peroxidation in Atlantic salmon (Salmo salar). Aquaculture 1987, 65, 43–54. [Google Scholar] [CrossRef]
- Thorarinsson, R.; Landolt, M.L.; Elliott, D.G.; Pascho, R.J.; Hardy, R.W. Effect of dietary vitamin E and selenium on growth, survival and the prevalence of Renibacterium salmoninarum infection in chinook salmon (Oncorhynchus tshawytscha). Aquaculture 1994, 121, 343–358. [Google Scholar] [CrossRef]
- Sallam, A.; Mansour, A.T.; Alsaqufi, A.; Salem, M.; El-Feky, M. Growth performance, antioxidative status, innate immunity, and ammonia stress resistance of Siganus rivulatus fed diet supplemented with zinc and zinc nanoparticles. Aquac. Rep. 2020, 18, 100410. [Google Scholar] [CrossRef]
- Lin, S.; Lin, X.; Yang, Y.; Li, F.; Luo, L. Comparison of chelated zinc and zinc sulfate as zinc sources for growth and immune response of shrimp (Litopenaeus vannamei). Aquaculture 2013, 406, 79–84. [Google Scholar] [CrossRef]
- Houng-Yung, C.; Yu-Chun, C.; Li-Chi, H.; Meng-Hsien, C. Dietary zinc requirements of juvenile grouper, Epinephelus malabaricus. Aquaculture 2014, 432, 360–364. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Alagawany, M.; Sewilam, H. The Role of Zinc Microelement in Aquaculture: A Review. Biol. Trace Elem. Res. 2022, 200, 3841–3853. [Google Scholar] [CrossRef]
- Hackney, A.C. Emerging issues in analytical chemistry. In Doping, Performance-Enhancing Drugs, and Hormones in Sport; Mechanisms of Action and Methods of Detection; Thomas, B., Ed.; Elsevier: Amsterdam, The Netherlands; RTI Press: Research Triangle Park, NC, USA, 2017; Volume 18, pp. 37–48. [Google Scholar]
- Hertz, Y.; Madar, Z.; Hepher, B.; Gertler, A. Glucose metabolism in the common carp (Cyprinus carpio L.): The effects of cobalt and chromium. Aquaculture 1989, 76, 255–267. [Google Scholar] [CrossRef]
- Sa, M.V.; Pezzato, L.; Barros, M.; Padilha, P. Relative bioavailability of zinc in supplemental inorganic and organic sources for Nile tilapia (Oreochromis niloticus) fingerlings. Aquac. Nutr. 2005, 11, 273–281. [Google Scholar]
- Asad, F.; Mubarik, M.S.; Ali, T.; Zahoor, M.K.; Ashrad, R.; Qamer, S. Effect of organic and in-organic chromium supplementation on growth performance and genotoxicity of Labeo rohita. Saudi J. Biol. Sci. 2019, 26, 1140–1145. [Google Scholar] [CrossRef]
- Watanabe, T.; Kiron, V.; Satoh, S. Trace minerals in fish nutrition. Aquaculture 1997, 151, 185–207. [Google Scholar] [CrossRef]
- Shankar, A.H.; Prasad, A.S. Zinc and immune function: The biological basis of altered resistance to infection. Am. J. Clin. Nutr. 1998, 68 (Suppl. 2), 447–463. [Google Scholar] [CrossRef]
- Naiel, M.A.E.; Negm, S.S.; Abd El-hameed, S.A.A.; Abdel-Latif, H.M.R. Dietary organic selenium improves growth, serum biochemical indices, immune responses, antioxidative capacity, and modulates transcription of stress-related genes in Nile tilapia reared under sub-optimal temperature. J. Therm. Biol. 2021, 99, 102999. [Google Scholar] [CrossRef]
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It’s role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Ning, L.; Yuwen, T.; Wang, W.; Wu, S.; Chen, F.; Zhang, H.; Pan, Q. Optimum selenium requirement of juvenile Nile tilapia, Oreochromis niloticus. Aquac. Nutr. 2020, 26, 528–535. [Google Scholar] [CrossRef]
- Pacitti, D.; Lawan, M.M.; Sweetman, J.; Martin, S.A.; Feldmann, J.; Secombes, C.J. Selenium Supplementation in Fish: A Combined Chemical and Biomolecular Study to Understand Sel-Plex Assimilation and Impact on Selenoproteome Expression in Rainbow Trout (Oncorhynchus mykiss). PLoS ONE 2015, 10, 1–17. [Google Scholar]
- Efsa, N. Panel: Scientific opinion on dietary reference values for selenium. EFSA J. 2014, 12, 3846. [Google Scholar]


{kind=link}
| Raw Materials (%) | Control | T1 (Zn-Cr) | T2 (Se-Cr) |
|---|---|---|---|
| Fishmeal (60%) | 5.00 | 5.00 | 5.00 |
| Corn gluten meal | 8.00 | 8.00 | 8.00 |
| Poultry meal (65% CP) | 7.00 | 7.00 | 7.00 |
| Rape seed | 7.00 | 7.00 | 7.00 |
| Fish hydrolysate | 4.00 | 4.00 | 4.00 |
| Soybean meal, dehulled | 25.50 | 25.50 | 25.50 |
| Wheat bran | 8.00 | 8.00 | 8.00 |
| Corn meal | 4.00 | 4.00 | 4.00 |
| Tapioca pulp | 4.00 | 4.00 | 4.00 |
| Tapioca chip | 25.00 | 25.00 | 25.00 |
| Lysine HCl | 0.43 | 0.43 | 0.43 |
| DL-methionine | 0.17 | 0.17 | 0.17 |
| Choline chloride | 0.30 | 0.30 | 0.30 |
| Mono-dicalcium phosphate | 0.50 | 0.50 | 0.50 |
| Salt | 0.10 | 0.10 | 0.10 |
| Micronutrient preservation premix | 1.00 | 1.00 | 1.00 |
| Zinc (zinc amino acid complex; 12%) | 0.00 | 0.05 | 0.00 |
| Chromium (Cr-L-methionine; 1000 ppb) | 0.00 | 0.05 | 0.05 |
| Selenium (selenomethionine; 1000 ppb) | 0.00 | 0.00 | 0.03 |
| Growth Parameters | Control | T1 (Zn-Cr) | T2 (Se-Cr) |
|---|---|---|---|
| Final weight (g/fish) Weight gain (g/fish) ADG (g/fish) | 547.72 ± 13.66 a | 586.58 ± 22.91 b | 569.49 ± 13.63 a |
| 355.15 ± 8.60 a | 394.07 ± 25.91 b | 377.19 ± 18.71 a | |
| 6.34 ± 0.15 a | 7.04 ± 0.46 b | 6.74 ± 0.33 a | |
| SGR (%/day) FCR | 1.87 ± 0.05 | 1.99 ± 0.11 | 1.94 ± 0.11 |
| 1.55 ± 0.04 b | 1.36 ± 0.10 a | 1.44 ± 0.04 b |
| Intestinal Villus Height (μm) | Control | T1 (Zn-Cr) | T2 (Se-Cr) |
|---|---|---|---|
| Anterior part (foregut) | 1197.89 ± 45.02 b | 1104.33 ± 36.98 b | 926.14 ± 77.38 a |
| Middle part (midgut) | 501.84 ± 104.95 a | 830.96 ± 88.57 b | 474.96 ± 42.93 a |
| Posterior part (hindgut) | 372.57 ± 22.50 a | 522.40 ± 57.40 b | 321.45 ± 72.09 a |
| Hematological Parameters | Control | T1 (Zn-Cr) | T2 (Se-Cr) |
|---|---|---|---|
| RBC (106 cell/mL) | 0.80 ± 0.23 | 0.84 ± 0.19 | 0.83 ± 0.20 |
| WBC (105 cell/mL) | 0.13 ± 0.05 | 0.16 ± 0.09 | 0.15 ± 0.00 |
| Hct (%) | 28.50 ± 3.70 | 33.75 ± 9.98 | 34.25 ± 4.19 |
| Hb (g/dL) | 6.39 ± 0.14 | 6.95 ± 0.86 | 6.52 ± 0.84 |
| Serum protein (mg/dL) | 6.25 ± 0.44 | 8.11 ± 1.14 | 6.80 ± 1.12 |
| IgM (g/L) | 0.49 ± 0.04 | 0.62 ± 0.10 | 0.54 ± 0.10 |
| Lysozyme activity (units/mL) | 138.75 ± 14.38 | 159.63 ± 25.32 | 163.00 ± 41.36 |
| Superoxide dismutase (units/mL) | 4.09 ± 1.31 | 4.72 ± 0.35 | 3.96 ± 0.25 |
| Glutathione (nmol) | 6.71 ± 0.33 | 8.42 ± 0.53 | 9.65 ± 2.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limwachirakhom, R.; Triwutanon, S.; Chumkam, S.; Jintasataporn, O. Effects of Chromium-L-Methionine in Combination with a Zinc Amino Acid Complex or Selenomethionine on Growth Performance, Intestinal Morphology, and Antioxidative Enzymes in Red Tilapia Oreochromis spp. Animals 2022, 12, 2182. https://doi.org/10.3390/ani12172182
Limwachirakhom R, Triwutanon S, Chumkam S, Jintasataporn O. Effects of Chromium-L-Methionine in Combination with a Zinc Amino Acid Complex or Selenomethionine on Growth Performance, Intestinal Morphology, and Antioxidative Enzymes in Red Tilapia Oreochromis spp. Animals. 2022; 12(17):2182. https://doi.org/10.3390/ani12172182
Chicago/Turabian StyleLimwachirakhom, Rawiwan, Supawit Triwutanon, Srinoy Chumkam, and Orapint Jintasataporn. 2022. "Effects of Chromium-L-Methionine in Combination with a Zinc Amino Acid Complex or Selenomethionine on Growth Performance, Intestinal Morphology, and Antioxidative Enzymes in Red Tilapia Oreochromis spp." Animals 12, no. 17: 2182. https://doi.org/10.3390/ani12172182
APA StyleLimwachirakhom, R., Triwutanon, S., Chumkam, S., & Jintasataporn, O. (2022). Effects of Chromium-L-Methionine in Combination with a Zinc Amino Acid Complex or Selenomethionine on Growth Performance, Intestinal Morphology, and Antioxidative Enzymes in Red Tilapia Oreochromis spp. Animals, 12(17), 2182. https://doi.org/10.3390/ani12172182