
Dietary Protected Butyrate Supplementation of Broilers Modulates Intestinal Tight Junction Proteins and Stimulates Endogenous Production of Short Chain Fatty Acids in the Caecum

 , ,
, ,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Performance Study
2.1.1. Animals and Housing
2.1.2. Experimental Design and Diets
2.1.3. Measurements
2.1.4. Statistical Analysis
2.2. Gut Mechanism Study
2.2.1. Animals and Housing
2.2.2. Experimental Design and Diets
2.2.3. Samplings
2.2.4. Assessment of Claudin-1, Claudin-3 and MCT-1 Protein Abundance
2.2.5. Measurement of SCFA Concentrations of Ingesta
2.2.6. Quantitative PCR Analysis of Targeted Genes in Caecal Ingesta
DNA Extraction
Quantitative PCR Analysis
2.2.7. Statistical Analyses
3. Results
3.1. Performance Study
3.2. Gut Mechanism Study
3.2.1. Claudin-1, Claudin-3 and MCT-1 Protein Abundance
3.2.2. SCFA Concentrations in Gut Digesta
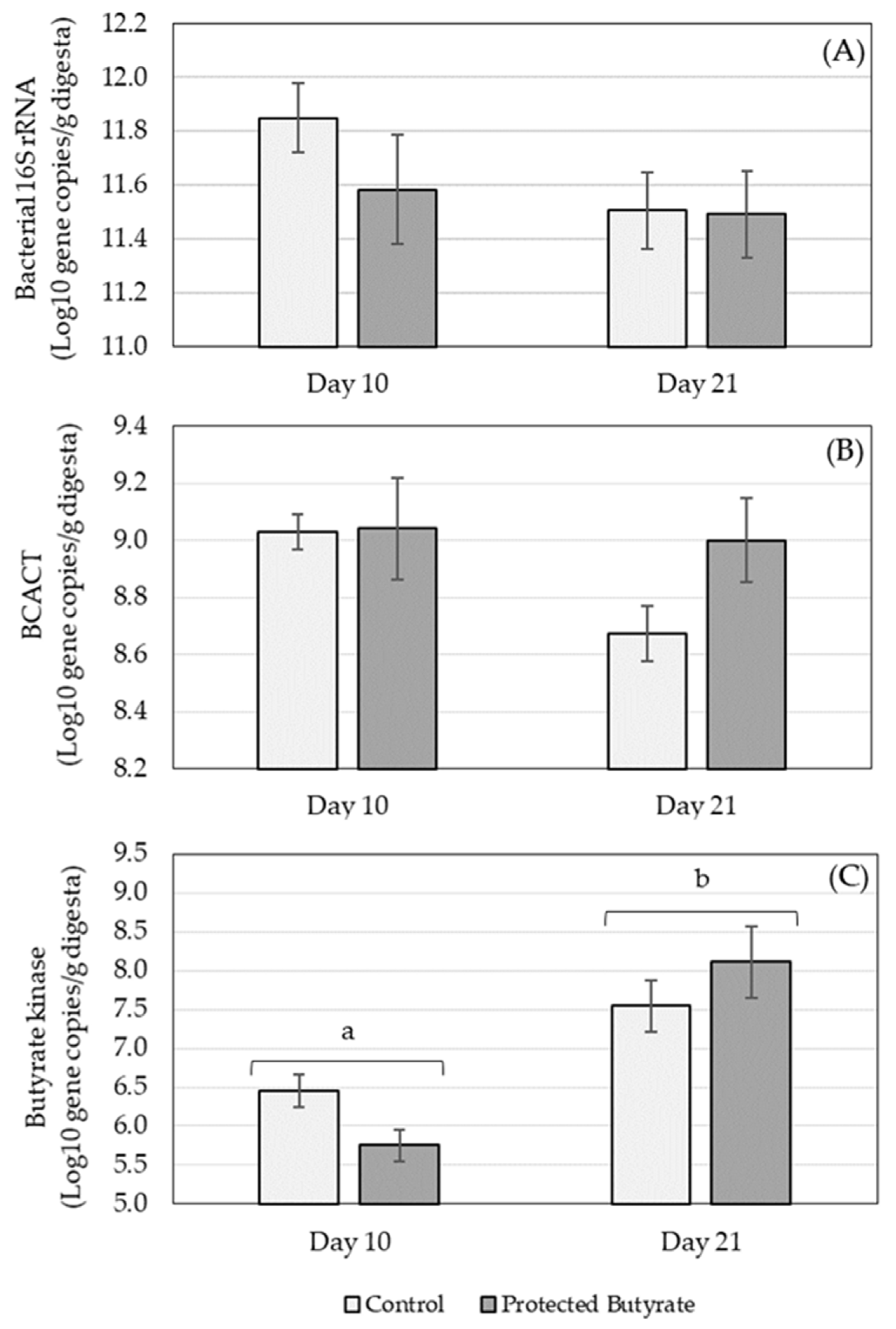
3.2.3. Targeted Caecal Microbiota Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Melaku, M.; Zhong, R.; Han, H.; Wan, F.; Yi, B.; Zhang, H. Butyric and Citric Acids and Their Salts in Poultry Nutrition: Effects on Gut Health and Intestinal Microbiota. Int. J. Mol. Sci. 2021, 22, 10392. [Google Scholar] [CrossRef] [PubMed]
- Eeckhaut, V.; van Immerseel, F.; Croubels, S.; de Baere, S.; Haesebrouck, F.; Ducatelle, R.; Louis, P.; Vandamme, P. Butyrate Production in Phylogenetically Diverse Firmicutes Isolated from the Chicken Caecum. Microb. Biotechnol. 2011, 4, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, A.; de Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borda-Molina, D.; Mátis, G.; Mackei, M.; Neogrády, Z.; Huber, K.; Seifert, J.; Camarinha-Silva, A. Caeca Microbial Variation in Broiler Chickens as a Result of Dietary Combinations Using Two Cereal Types, Supplementation of Crude Protein and Sodium Butyrate. Front. Microbiol. 2021, 11, 617800. [Google Scholar] [CrossRef] [PubMed]
- Salvi, P.S.; Cowles, R.A. Butyrate and the Intestinal Epithelium: Modulation of Proliferation and Inflammation in Homeostasis and Disease. Cells 2021, 10, 1775. [Google Scholar] [CrossRef]
- Panda, A.K.; Rao, S.V.R.; Raju, M.V.L.N.; Sunder, G.S. Effect of Butyric Acid on Performance, Gastrointestinal Tract Health and Carcass Characteristics in Broiler Chickens. Asian-Australas. J. Anim. Sci. 2009, 22, 1026–1031. [Google Scholar] [CrossRef]
- Zhang, W.H.; Jiang, Y.; Zhu, Q.F.; Gao, F.; Dai, S.F.; Chen, J.; Zhou, G.H. Sodium Butyrate Maintains Growth Performance by Regulating the Immune Response in Broiler Chickens. Br. Poult. Sci. 2011, 52, 292–301. [Google Scholar] [CrossRef]
- Abdelqader, A.; Al-Fataftah, A.R. Effect of Dietary Butyric Acid on Performance, Intestinal Morphology, Microflora Composition and Intestinal Recovery of Heat-Stressed Broilers. Livest. Sci. 2016, 183, 78–83. [Google Scholar] [CrossRef]
- Hu, Z.; Guo, Y. Effects of Dietary Sodium Butyrate Supplementation on the Intestinal Morphological Structure, Absorptive Function and Gut Flora in Chickens. Anim. Feed Sci. Technol. 2007, 132, 240–249. [Google Scholar] [CrossRef]
- Antongiovanni, M.; Buccioni, A.; Petacchi, F.; Leeson, S.; Minieri, S.; Martini, A.; Cecchi, R. Butyric Acid Glycerides in the Diet of Broiler Chickens: Effects on Gut Histology and Carcass Composition. Ital. J. Anim. Sci. 2007, 6, 19–25. [Google Scholar] [CrossRef]
- Chamba, F.; Puyalto, M.; Ortiz, A.; Torrealba, H.; Mallo, J.J.; Riboty, R. Effect of Partially Protected Sodium Butyrate on Performance, Digestive Organs, Intestinal Villi and E. Coli Development in Broilers Chickens. Int. J. Poult. Sci. 2014, 13, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Mátis, G.; Neogrády, Z.; Csikó, G.; Kulcsár, A.; Kenéz, Á.; Huber, K. Effects of Orally Applied Butyrate Bolus on Histone Acetylation and Cytochrome P450 Enzyme Activity in the Liver of Chicken—A Randomized Controlled Trial. Nutr. Metab. 2013, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Mátis, G.; Kulcsár, A.; Turowski, V.; Fébel, H.; Neogrády, Z.; Huber, K. Effects of Oral Butyrate Application on Insulin Signaling in Various Tissues of Chickens. Domest. Anim. Endocrinol. 2015, 50, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Kulcsár, A.; Mátis, G.; Molnár, A.; Petrilla, J.; Husvéth, F.; Huber, K.; Dublecz, K.; Neogrády, Z. Effects of Butyrate on the Insulin Homeostasis of Chickens Kept on Maize- or Wheat-Based Diets. Acta Vet. Hung. 2016, 64, 482–496. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Ajuwon, K.M. Butyrate Modifies Intestinal Barrier Function in IPEC-J2 Cells through a Selective Upregulation of Tight Junction Proteins and Activation of the Akt Signaling Pathway. PLoS ONE 2017, 12, e0179586. [Google Scholar] [CrossRef]
- Wang, H.B.; Wang, P.Y.; Wang, X.; Wan, Y.L.; Liu, Y.C. Butyrate Enhances Intestinal Epithelial Barrier Function via Up-Regulation of Tight Junction Protein Claudin-1 Transcription. Dig. Dis. Sci. 2012, 57, 3126–3135. [Google Scholar] [CrossRef]
- Song, B.; Li, H.; Wu, Y.; Zhen, W.; Wang, Z.; Xia, Z.; Guo, Y. Effect of Microencapsulated Sodium Butyrate Dietary Supplementation on Growth Performance and Intestinal Barrier Function of Broiler Chickens Infected with Necrotic Enteritis. Anim. Feed Sci. Technol. 2017, 232, 6–15. [Google Scholar] [CrossRef]
- Song, J.; Li, Q.; Li, P.; Liu, R.R.; Cui, H.; Zheng, M.; Everaert, N.; Zhao, G.; Wen, J. The Effects of Inulin on the Mucosal Morphology and Immune Status of Specific Pathogen-Free Chickens. Poult. Sci. 2018, 97, 3938–3946. [Google Scholar] [CrossRef]
- Günzel, D.; Yu, A.S.L. Claudins and the Modulation of Tight Junction Permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [Green Version]
- StatSoft Inc. Statistica (Data Analysis Software System); StatSoft Inc.: Tulsa, OK, USA, 2016. [Google Scholar]
- Aviagen. Aviagen Broiler Ross 308 Performance Objectives; Aviagen: Chester, UK, 2014. [Google Scholar]
- Kettunen, H.; Vuorenmaa, J.; Rinttilä, T.; Grönberg, H.; Valkonen, E.; Apajalahti, J. Natural Resin Acid –Enriched Composition as a Modulator of Intestinal Microbiota and Performance Enhancer in Broiler Chicken. J. Appl. Anim. Nutr. 2015, 3, e2. [Google Scholar] [CrossRef]
- Mátis, G.; Petrilla, J.; Kulcsár, A.; van den Bighelaar, H.; Boomsma, B.; Neogrády, Z.; Fébel, H. Effects of Dietary Butyrate Supplementation and Crude Protein Level on Carcass Traits and Meat Composition of Broiler Chickens. Arch. Anim. Breed. 2019, 62, 527–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeson, S.; Namkung, H.; Antongiovanni, M.; Lee, E.H. Effect of Butyric Acid on the Performance and Carcass Yield of Broiler Chickens. Poult. Sci. 2005, 84, 1418–1422. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; McCartney, E.; Knox, A.; Francesch, M.; Oka, K.; Wada, K.; Ideno, M.; Uno, K.; Kozłowski, K.; Jankowski, J.; et al. Effects of the Butyric Acid-Producing Strain Clostridium butyricum MIYAIRI 588 on Broiler and Piglet Zootechnical Performance and Prevention of Necrotic Enteritis. Anim. Sci. J. 2018, 89, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.D.; Bayir, H.O.; Cosby, D.E.; Cox, N.A.; Williams, S.M.; Fowler, J. Evaluation of Encapsulated Sodium Butyrate on Growth Performance, Energy Digestibility, Gut Development, and Salmonella Colonization in Broilers. Poult. Sci. 2017, 96, 3638–3644. [Google Scholar] [CrossRef]
- Feng, W.; Wu, Y.; Chen, G.; Fu, S.; Li, B.; Huang, B.; Wang, D.; Wang, W.; Liu, J. Sodium Butyrate Attenuates Diarrhea in Weaned Piglets and Promotes Tight Junction Protein Expression in Colon in a GPR109A-Dependent Manner. Cell. Physiol. Biochem. 2018, 47, 1617–1629. [Google Scholar] [CrossRef]
- Silva, J.P.B.; Navegantes-Lima, K.C.; Oliveira, A.L.B.; Rodrigues, D.V.S.; Gaspar, S.L.F.; Monteiro, V.V.S.; Moura, D.P.; Monteiro, M.C. Protective Mechanisms of Butyrate on Inflammatory Bowel Disease. Curr. Pharm. Des. 2018, 24, 4154–4166. [Google Scholar] [CrossRef]
- Grilli, E.; Tugnoli, B.; Foerster, C.J.; Piva, A. Butyrate Modulates Inflammatory Cytokines and Tight Junctions Components along the Gut of Weaned Pigs. J. Anim. Sci. 2016, 94, 433–436. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Miao, W.; Wu, X.; Wang, K.; Wang, W.; Wang, Y.; Li, Z.; Liu, J.; Li, L.; Peng, L. Sodium Butyrate Promotes Reassembly of Tight Junctions in Caco-2 Monolayers Involving Inhibition of MLCK/MLC2 Pathway and Phosphorylation of PKCβ2. Int. J. Mol. Sci. 2016, 17, 1696. [Google Scholar] [CrossRef] [Green Version]
- Cuff, M.A.; Lambert, D.W.; Shirazi-Beechey, S.P. Substrate-Induced Regulation of the Human Colonic Monocarboxylate Transporter, MCT1. J. Physiol. 2002, 539, 361–371. [Google Scholar] [CrossRef]
- Moquet, P.C.A. Butyrate in Broiler Diets: Impact of Butyrate Presence in Distinct Gastrointestinal Tract Segments on Digestive Function, Microbiota Composition and Immune Responses. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2018. [Google Scholar]
- Kulcsár, A.; Mátis, G.; Molnár, A.; Petrilla, J.; Wágner, L.; Fébel, H.; Husvéth, F.; Dublecz, K.; Neogrády, Z. Nutritional Modulation of Intestinal Drug-Metabolizing Cytochrome P450 by Butyrate of Different Origin in Chicken. Res. Vet. Sci. 2017, 113, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; van Immerseel, F. From the Gut to the Peripheral Tissues: The Multiple Effects of Butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, A.; Hess, C.; Pál, L.; Wágner, L.; Awad, W.A.; Husvéth, F.; Hess, M.; Dublecz, K. Composition of Diet Modifies Colonization Dynamics of Campylobacter Jejuni in Broiler Chickens. J. Appl. Microbiol. 2015, 118, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, Receptor for Niacin and the Commensal Metabolite Butyrate, Suppresses Colonic Inflammation and Carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Parameter | Starter (1–14 days) | Grower (15–35 days) |
|---|---|---|
| Ingredients (%): | ||
| Corn | 59.31 | 62.64 |
| Soybean meal | 34.36 | 29.66 |
| Soybean oil | 2.00 | 3.68 |
| Sodium bicarbonate | 0.15 | 0.15 |
| Sodium chloride | 0.21 | 0.21 |
| Limestone | 1.30 | 1.16 |
| Monocalcium phosphate | 1.82 | 1.64 |
| Choline chloride | 0.09 | 0.09 |
| DL-Methionine | 0.26 | 0.23 |
| L-Lysine | 0.16 | 0.19 |
| L-Threonine | 0.09 | 0.10 |
| Vitamins & trace minerals | 0.25 | 0.25 |
| Calculated Nutritional Values: | ||
| ME (kcal/kg) | 2950 | 3100 |
| Crude protein (g/kg) | 215.0 | 195.0 |
| Lysine (g/kg) | 12.50 | 11.50 |
| Methionine (g/kg) | 5.86 | 5.34 |
| Met + Cys (g/kg) | 9.05 | 8.70 |
| Threonine (g/kg) | 9.50 | 8.30 |
| Tryptophan (g/kg) | 2.51 | 2.25 |
| Calcium (g/kg) | 10.00 | 9.00 |
| Phosphorus (g/kg) | 5.00 | 4.50 |
| Parameter | Starter (1–10 days) | Grower (11–21 days) |
|---|---|---|
| Ingredients (%): | ||
| Corn | 33.88 | 22.55 |
| Wheat (incl. xylanase) | 25.00 | 40.00 |
| Soybean meal | 26.00 | 24.00 |
| Rapeseed meal | 4.00 | 5.00 |
| Full fat soyabeans | 5.00 | - |
| Animal fat | - | 4.00 |
| Soyabean oil | 2.25 | 1.20 |
| Limestone | 1.75 | 1.40 |
| Monocalcium phosphate | 0.70 | 0.40 |
| Vitamins & trace minerals | 0.50 | 0.50 |
| Sodium bicarbonate | 0.24 | 0.21 |
| Sodium chloride | 0.22 | 0.21 |
| DL-Methionine | 0.22 | 0.20 |
| L-Lysine HCl | 0.17 | 0.22 |
| L-Threonine | 0.05 | 0.07 |
| L-Valine | 0.01 | 0.03 |
| Phytase | 0.01 | 0.01 |
| Calculated Nutritional Values: | ||
| ME (kcal/kg) | 2849 | 2963 |
| Crude protein (g/kg) | 215 | 201 |
| Lysine (g/kg) | 12.5 | 11.7 |
| Methionine (g/kg) | 5.5 | 5.1 |
| Met + Cys (g/kg) | 9.2 | 8.6 |
| Threonine (g/kg) | 8.4 | 8.0 |
| Tryptophan (g/kg) | 2.6 | 2.4 |
| Calcium (g/kg) | 9.0 | 7.2 |
| Phosphorus (g/kg) | 5.6 | 4.8 |
| Parameter | Control | Low | Medium | High | Pooled SEM | p | p Linear | p Square |
|---|---|---|---|---|---|---|---|---|
| Body Weight (g): | ||||||||
| Day 1 | 37.4 | 37.5 | 37.4 | 37.4 | 0.028 | 0.607 | - | - |
| Day 14 | 350.8 b | 361.3 ab | 368.3 a | 369.4 a | 2.479 | 0.024 | 0.042 | 0.210 |
| Day 35 | 1960.3 b | 2030.4 b | 2113.2 a | 2029.3 b | 14.722 | 0.001 | <0.001 | 0.002 |
| Average Daily Gain (g): | ||||||||
| Days 1–14 | 22.4 b | 23.1 ab | 23.6 a | 23.7 a | 0.177 | 0.023 | 0.042 | 0.210 |
| Days 15–35 | 76.6 b | 79.6 ab | 83.0 a | 79.1 ab | 0.626 | 0.002 | <0.001 | 0.001 |
| Days 1–35 | 54.9 c | 57.5 ab | 59.3 a | 56.9 bc | 0.431 | 0.002 | <0.001 | 0.001 |
| Average Daily Feed Intake (g): | ||||||||
| Days 1–14 | 33.0 | 32.7 | 32.9 | 32.7 | 0.230 | 0.942 | 0.876 | 0.957 |
| Days 15–35 | 121.6 | 125.1 | 125.9 | 121.4 | 0.828 | 0.114 | 0.022 | 0.015 |
| Days 1–35 | 86.2 | 88.1 | 88.7 | 85.5 | 0.517 | 0.090 | 0.023 | 0.013 |
| Feed Conversion Ratio: | ||||||||
| Days 1–14 | 1.460 | 1.401 | 1.394 | 1.391 | 0.012 | 0.132 | 0.075 | 0.202 |
| Days 15–35 | 1.579 b | 1.551 ab | 1.534 a | 1.518 a | 0.003 | <0.001 | 0.024 | 0.339 |
| Days 1–35 | 1.559 c | 1.525 b | 1.511 ab | 1.496 a | 0.006 | <0.001 | 0.006 | 0.170 |
| Livability (%) | 97.8 | 100.0 | 100.0 | 96.7 | 0.588 | 0.106 | - | - |
| Parameter (ng/mg Protein) | Day 10 | Day 21 | Pooled SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Control | Protected Butyrate | Control | Protected Butyrate | Age | Treatment | Interaction | ||
| Gizzard: | ||||||||
| Claudin-1 | 0.307 | 0.316 | 0.111 | 0.154 | 0.010 | <0.001 | 0.010 | 0.160 |
| Claudin-3 | 0.871 | 0.578 | 0.278 | 0.359 | 0.047 | <0.001 | 0.106 | 0.002 |
| MCT-1 | 0.625 | 0.723 | 0.280 | 0.409 | 0.019 | <0.001 | <0.001 | 0.780 |
| Duodenum: | ||||||||
| Claudin-1 | 0.248 | 0.223 | 0.117 | 0.091 | 0.012 | <0.001 | 0.041 | 0.968 |
| Claudin-3 | 0.428 | 0.347 | 0.189 | 0.189 | 0.022 | <0.001 | 0.078 | 0.079 |
| MCT-1 | 0.516 | 0.377 | 0.128 | 0.119 | 0.026 | <0.001 | 0.006 | 0.015 |
| Jejunum + Ileum: | ||||||||
| Claudin-1 | 0.240 | 0.364 | 0.128 | 0.157 | 0.025 | <0.001 | 0.004 | 0.061 |
| Claudin-3 | 0.528 | 0.609 | 0.280 | 0.388 | 0.028 | <0.001 | 0.002 | 0.640 |
| MCT-1 | 0.697 | 0.881 | 0.172 | 0.374 | 0.041 | <0.001 | <0.001 | 0.823 |
| Caecum: | ||||||||
| Claudin-1 | 0.435 | 0.387 | 0.184 | 0.192 | 0.022 | <0.001 | 0.376 | 0.211 |
| Claudin-3 | 1.197 | 0.990 | 0.460 | 0.480 | 0.060 | <0.001 | 0.127 | 0.064 |
| MCT-1 | 0.876 | 0.661 | 0.339 | 0.349 | 0.052 | <0.001 | 0.053 | 0.041 |
| Gut Site | Parameter (μmol/g Digesta) | Day 10 | Day 21 | Pooled SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Control | Protected Butyrate | Control | Protected Butyrate | Age | Treatment | Interaction | |||
| Gizzard | Acetate | 6.72 | 5.24 | 5.48 | 4.79 | 0.387 | 0.053 | 0.012 | 0.348 |
| Propionate | 0.272 | 0.384 | 0.305 | 0.262 | 0.023 | 0.053 | 0.130 | 0.002 | |
| Butyrate | 0.036 | 0.196 | 0.029 | 0.184 | 0.020 | 0.678 | <0.001 | 0.918 | |
| Valerate | 0.012 | 0.009 | 0.009 | 0.008 | 0.002 | 0.158 | 0.116 | 0.435 | |
| Total SCFA | 7.04 | 5.82 | 5.83 | 5.24 | 0.402 | 0.049 | 0.044 | 0.472 | |
| Duodenum | Acetate | 3.57 | 3.02 | 6.04 | 3.14 | 0.364 | 0.001 | <0.001 | 0.004 |
| Propionate | 0.206 | 0.314 | 0.425 | 0.204 | 0.023 | 0.026 | 0.023 | <0.001 | |
| Butyrate | 0.023 | 0.039 | 0.021 | 0.037 | 0.004 | 0.688 | 0.001 | 0.955 | |
| Valerate | 0.012 | 0.009 | 0.010 | 0.008 | 0.002 | 0.493 | 0.222 | 0.980 | |
| Total SCFA | 3.81 | 3.38 | 6.50 | 3.39 | 0.376 | 0.001 | <0.001 | 0.001 | |
| Jejunum + Ileum | Acetate | 6.51 | 5.85 | 3.88 | 4.60 | 0.390 | <0.001 | 0.980 | 0.089 |
| Propionate | 0.218 | 0.248 | 0.181 | 0.156 | 0.013 | <0.001 | 0.809 | 0.043 | |
| Butyrate | 0.011 | 0.016 | 0.009 | 0.009 | 0.001 | <0.001 | 0.005 | 0.002 | |
| Valerate | 0.010 | 0.008 | 0.005 | 0.005 | 0.001 | 0.158 | 0.048 | 0.434 | |
| Total SCFA | 6.75 | 6.12 | 4.08 | 4.77 | 0.396 | <0.001 | 0.974 | 0.107 | |
| Caecum | Acetate | 79.37 | 90.51 | 62.66 | 75.37 | 4.877 | 0.003 | 0.023 | 0.877 |
| Propionate | 2.18 | 2.85 | 3.43 | 4.53 | 0.421 | 0.002 | 0.047 | 0.623 | |
| Butyrate | 5.83 | 8.36 | 7.44 | 10.64 | 1.044 | 0.079 | 0.012 | 0.760 | |
| Valerate | 0.055 | 0.063 | 0.514 | 0.817 | 0.097 | <0.001 | 0.105 | 0.148 | |
| Total SCFA | 87.44 | 102.10 | 74.05 | 91.36 | 5.604 | 0.046 | 0.009 | 0.820 | |
| Gut Site | Parameter | Age | Butyrate |
|---|---|---|---|
| Gizzard | Claudin-1 | ↓ | ↑ |
| Claudin-3 | ↓ | - | |
| MCT-1 | ↓ | ↑ | |
| Acetate | ↓ | - | |
| Propionate | - | - | |
| Butyrate | - | ↑ | |
| Valerate | - | - | |
| Total SCFA | ↓ | ↓ | |
| Duodenum | Claudin-1 | ↓ | ↓ |
| Claudin-3 | ↓ | - | |
| MCT-1 | ↓ | ↓ | |
| Acetate | ↑ | ↓ | |
| Propionate | ↑ | ↓ | |
| Butyrate | - | ↑ | |
| Valerate | - | - | |
| Total SCFA | ↑ | ↓ | |
| Jejunum + Ileum | Claudin-1 | ↓ | ↑ |
| Claudin-3 | ↓ | ↑ | |
| MCT-1 | ↓ | ↑ | |
| Acetate | ↓ | - | |
| Propionate | ↓ | - | |
| Butyrate | ↓ | ↑ | |
| Valerate | - | ↓ | |
| Total SCFA | ↓ | - | |
| Caecum | Claudin-1 | ↓ | - |
| Claudin-3 | ↓ | - | |
| MCT-1 | ↓ | - | |
| Acetate | ↓ | ↑ | |
| Propionate | ↑ | ↑ | |
| Butyrate | ↑ | ↑ | |
| Valerate | ↑ | ↑ | |
| Total SCFA | ↓ | ↑ | |
| Bacterial 16S RNA | - | - | |
| BCACT | - | - | |
| Butyrate kinase | ↑ | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mátis, G.; Mackei, M.; Boomsma, B.; Fébel, H.; Nadolna, K.; Szymański, Ł.; Edwards, J.E.; Neogrády, Z.; Kozłowski, K. Dietary Protected Butyrate Supplementation of Broilers Modulates Intestinal Tight Junction Proteins and Stimulates Endogenous Production of Short Chain Fatty Acids in the Caecum. Animals 2022, 12, 1940. https://doi.org/10.3390/ani12151940
Mátis G, Mackei M, Boomsma B, Fébel H, Nadolna K, Szymański Ł, Edwards JE, Neogrády Z, Kozłowski K. Dietary Protected Butyrate Supplementation of Broilers Modulates Intestinal Tight Junction Proteins and Stimulates Endogenous Production of Short Chain Fatty Acids in the Caecum. Animals. 2022; 12(15):1940. https://doi.org/10.3390/ani12151940
Chicago/Turabian StyleMátis, Gábor, Máté Mackei, Bart Boomsma, Hedvig Fébel, Katarzyna Nadolna, Łukasz Szymański, Joan E. Edwards, Zsuzsanna Neogrády, and Krzysztof Kozłowski. 2022. "Dietary Protected Butyrate Supplementation of Broilers Modulates Intestinal Tight Junction Proteins and Stimulates Endogenous Production of Short Chain Fatty Acids in the Caecum" Animals 12, no. 15: 1940. https://doi.org/10.3390/ani12151940
APA StyleMátis, G., Mackei, M., Boomsma, B., Fébel, H., Nadolna, K., Szymański, Ł., Edwards, J. E., Neogrády, Z., & Kozłowski, K. (2022). Dietary Protected Butyrate Supplementation of Broilers Modulates Intestinal Tight Junction Proteins and Stimulates Endogenous Production of Short Chain Fatty Acids in the Caecum. Animals, 12(15), 1940. https://doi.org/10.3390/ani12151940