1. Introduction
Avian coccidiosis is a worldwide disease that is caused by protozoan parasites of
Eimeria spp. Seven species of
Eimeria including
E. acervuline,
E. brunetti,
E. maxima,
E. mitis,
E necatrix,
E. praecox, and
E. tenella have been identified in the broiler chicken industry [
1]. The prevalence of coccidiosis usually occurs with a mixed infection instead of a single species with the mixed infection by
E. acervulina,
E. maxima, and
E. tenella being more prevalent [
2]. These three species of
Eimeria spp. preferentially invade and multiply in the regions of duodenum, jejunum plus ileum, and ceca, respectively; however, intersectional infection also can be observed [
3].
Damage to gastrointestinal epithelial cells resulting in depressed nutrient utilization and growth performance in birds leads to USD 14.5 billion losses annually in poultry production due to coccidiosis [
4,
5].
Eimeria-induced gastrointestinal damage is also considered to influence the regulation of intestinal tight junctions and nutrient transporters. The changes to intestinal nutrient transporters including sugars, amino acids, and mineral transporters may partly contribute to the
Eimeria-induced growth depression [
6,
7]. In addition, it has been demonstrated that
Eimeria infection impacted the microbial population and composition in the hindgut of broiler chickens, resulting in cecal dysbacteriosis. The infection decreased the abundance of beneficial bacteria such as carbohydrate-fermenting
Ruminococcaceae, butyrate-producing
Faecalibacterium, and probiotic bacterium
Lactobacillus, whereas enriched opportunistic pathogenic bacteria of the genera
Enterococcus and
Streptococcus [
8]. The dysbacteriosis in the hindgut indicates a shift of fermentation patterns in ceca.
Anticoccidial drugs and live vaccines have been applied in the commercial poultry industry as conventional methods to prevent coccidiosis. However, after exposure to antibiotics for decades,
Eimeria spp. have developed resistance to these synthetic anticoccidial drugs whereas vaccines are costly for production and storage [
9]. In view of the need for alternatives to antibiotics, dietary antibiotic alternatives are possible options to help treat or prevent coccidiosis. Probiotics such as
Lactobacillus-based probiotic,
Pediococcus acidilactici-based probiotic, and
B. subtilis-based probiotics have shown positive results in enhancing mucosal immunity, lessening
Eimeria-induced growth depression, and improving resistance to
Eimeria infection [
10,
11,
12]. Prebiotics with the ability to maintain intestinal flora balance has also shown promise by enriching beneficial bacteria [
12].
It has been reported that the effects of prebiotic and carbohydrase supplementation on the performance of broilers challenged with coccidia are similar, supporting the hypothesis that oligosaccharides are released from carbohydrase-degraded non-starch polysaccharides and play prebiotic roles in birds’ gastrointestinal tract [
13,
14,
15]. In a previous study with supplementation of protease alone or in combination with carbohydrases, the effects of enzymes on the disappearance of non-starch polysaccharides in digesta and accumulation of lower molecular weight carbohydrates were similar [
16]. In view of the potential effects of enzyme supplementation and dietary fiber profile affecting the generation of prebiotic substances in the digestive tract of birds [
17], there is a possibility of these factors having positive effects on the intestinal milieu of broiler chickens challenged with coccidiosis. Consequently, two experiments were conducted to investigate the potential and mechanism of enzymes on mitigating the negative effects of
Eimeria challenge on growth performance, gene expression of nutrient transporters, and cecal fermentation patterns. A two-diet model, high fiber-adequate protein, and low fiber-low protein were chosen from previous work [
17], where xylanase and protease showed effects on digesta concentration of oligosaccharides and cecal short-chain fatty acids (SCFA). Consequently, these diets were used separately in the current experiment because their interactions in a factorial treatment arrangement setting have been demonstrated previously [
17].
4. Discussion
The objective of the two experiments reported herein was to investigate the potential and mechanisms of exogenous enzymes action on mitigating the negative effects of
Eimeria challenge on growth performance, gene expression of tight junctions and nutrient transporters, and cecal fermentation pattern.
Eimeria-induced reduction in broiler growth performance is positively correlated with infective dose [
21,
22]. Bodyweight gain of birds at 1 to 6 DPI (day 14 to day 20) can be linearly reduced from 27% to 49% when challenged by increasing oocysts doses of mixed
Eimeria spp. (6250
E. maxima; 6250
E.tenella; 31,250
E. acervuline to 50,000
E. maxima; 50,000
E. tenella; 250,000
E. acervulina) [
21]. A medium-low dose of
Eimeria was used in the current study in order to develop a mild infection resulting in a 63% reduction in body weight gain at 6 DPI, which was more severe than expected but similar to what others have reported [
23,
24]. There are species differences in the extent to which
Eimeria impacts growth performance. The less pathogenic species such as
E. praecox and
E. mitis, which do not cause lesions, generally have less influence on growth performance whereas
E. maxima, which mainly invades jejunum and ileum, showed a more serious impairment on feed conversion ratio (FCR) [
1,
25]. Exogenous enzymes do not have anticoccidial activities. However, dietary inclusion of such enzymes may reduce the negative effect of coccidia infection via the independent effect of such enzymes on promoting growth performance, nutrient utilization, the integrity of intestinal epithelial cells, or bacterial balance. For example, enzyme complexes including xylanase or protease have been shown to significantly decrease FCR in broiler chickens challenged with coccidia [
14,
26,
27]. In Expt. 2 of the current study, enzyme supplementation increased feed intake without any effect on WG or the gain–feed ratio. The enzyme’s effect during coccidia challenge is not universally observed. Enzyme supplementation had no significant mitigation effect on the impairment of the growth performance associated with coccidiosis in Parker et al. [
28] or in Expt. 1 of the current study. Dietary and other factors may play a role in differences in response observed from different studies.
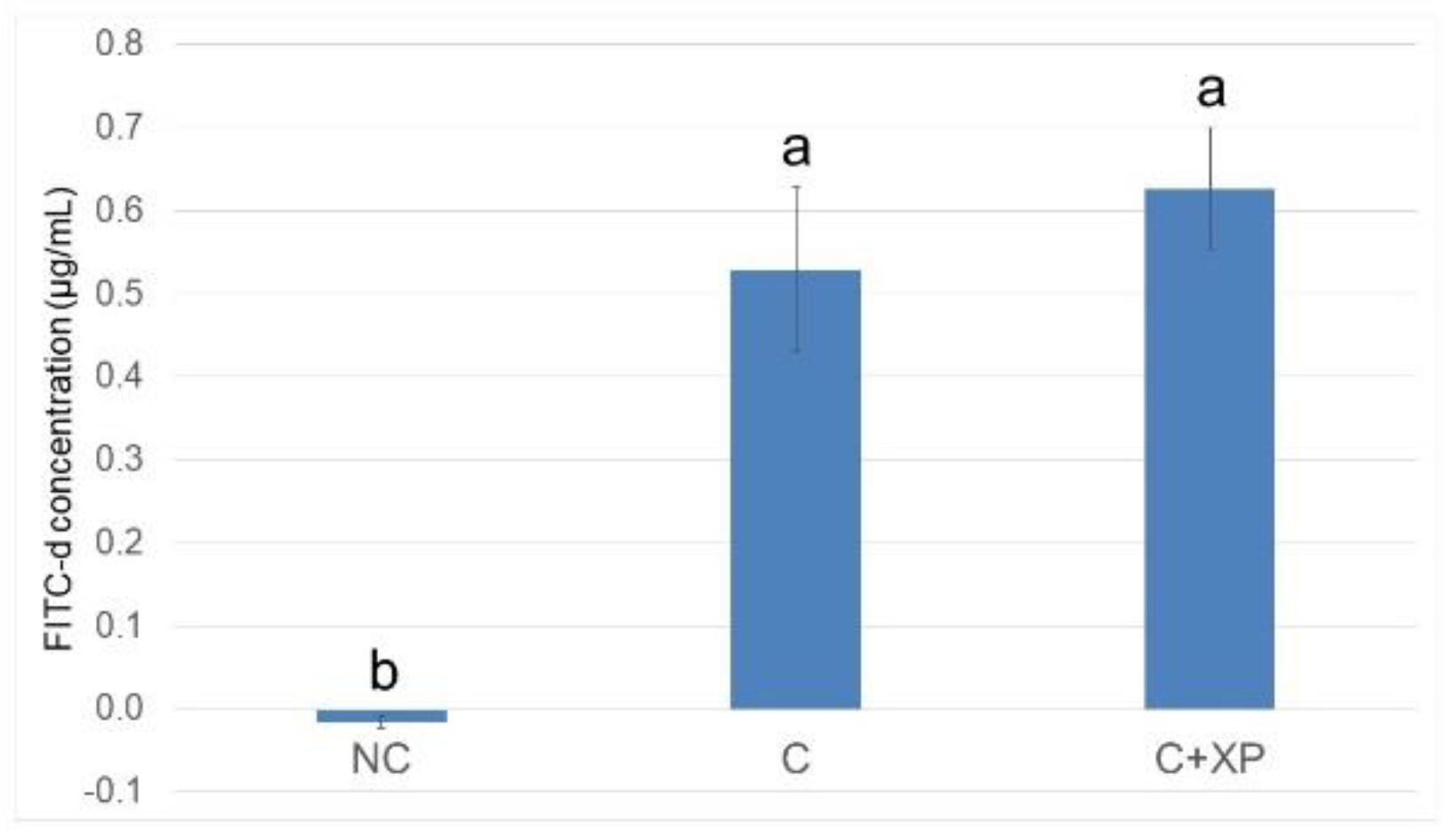
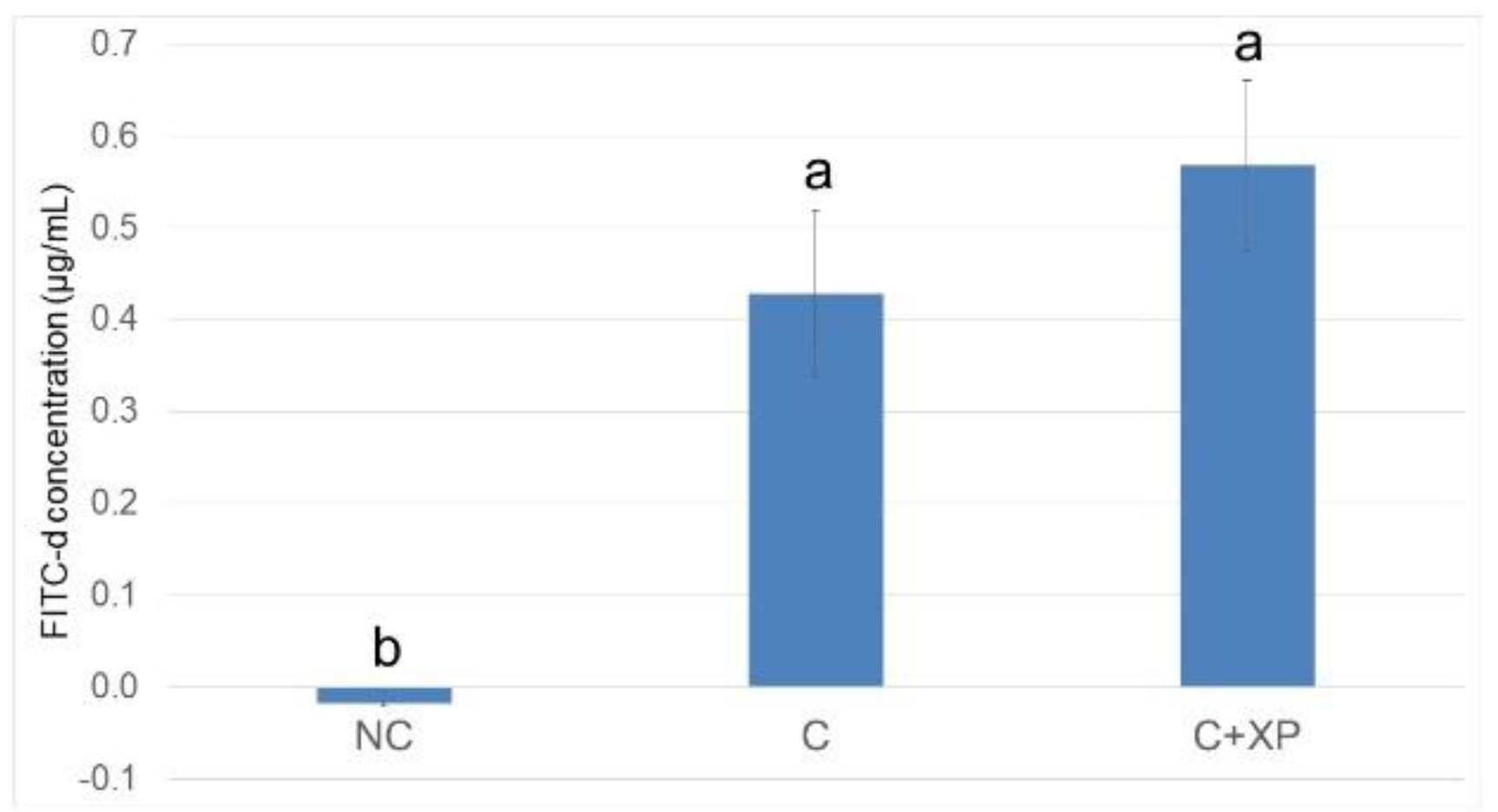
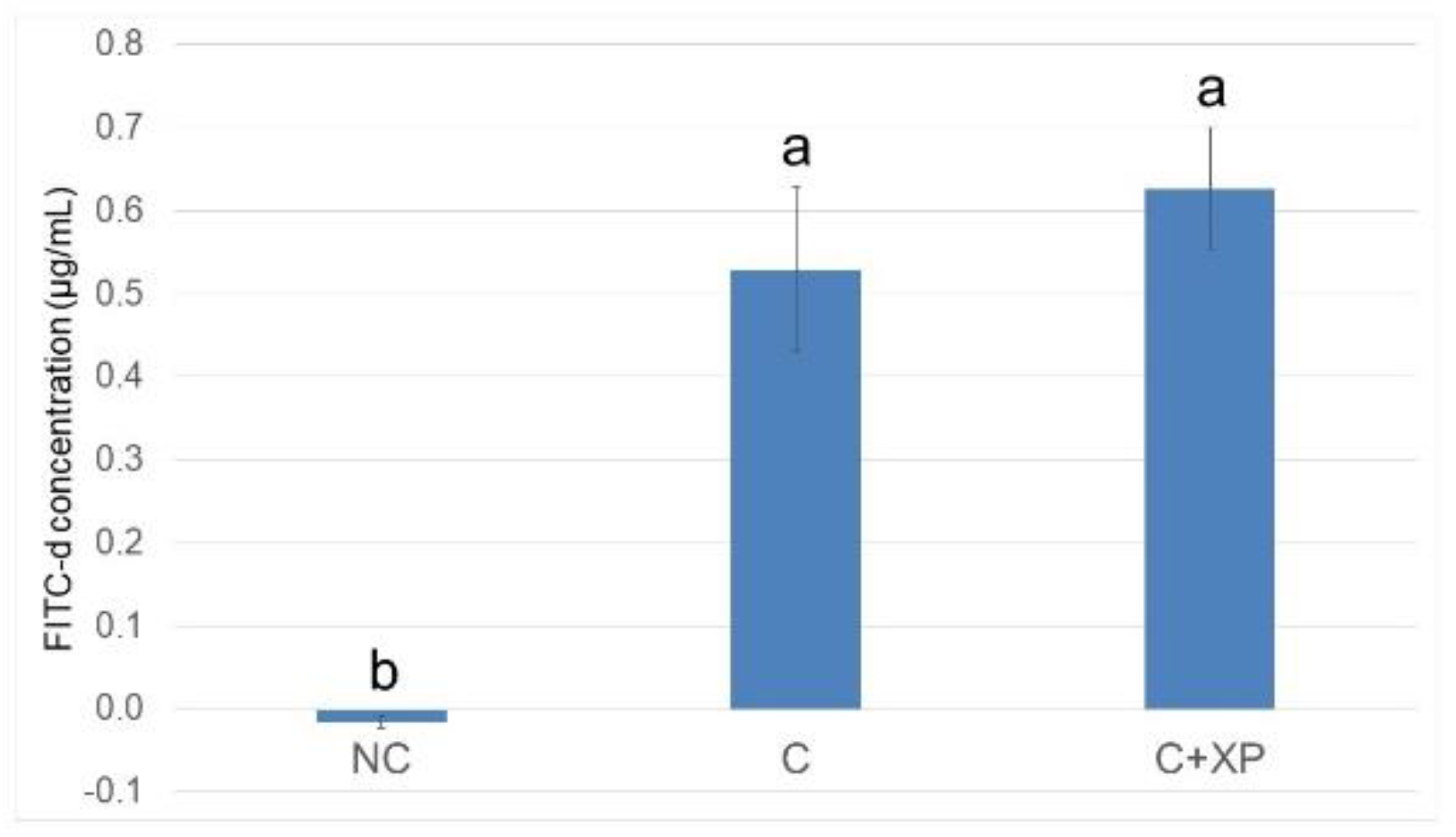
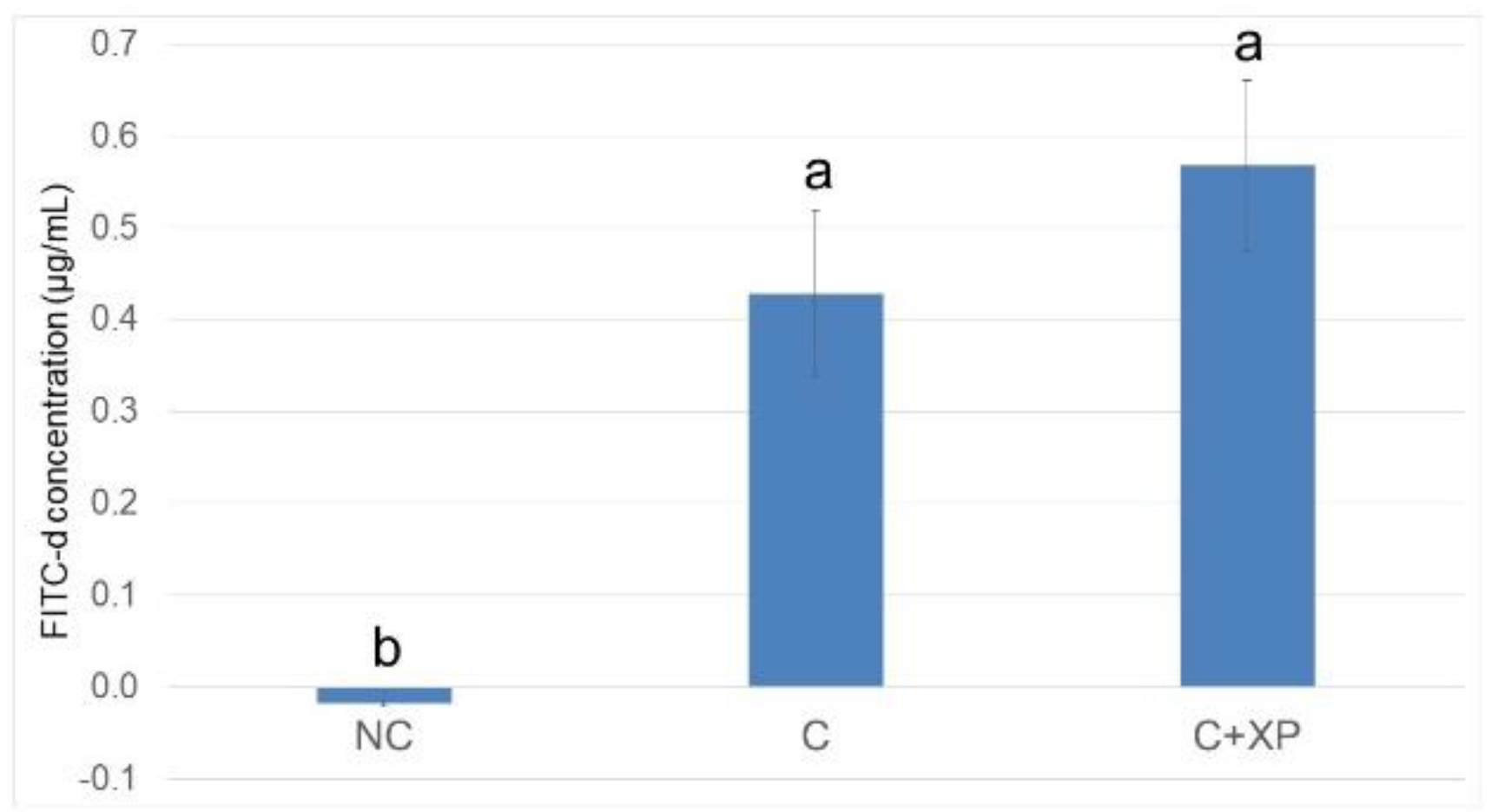
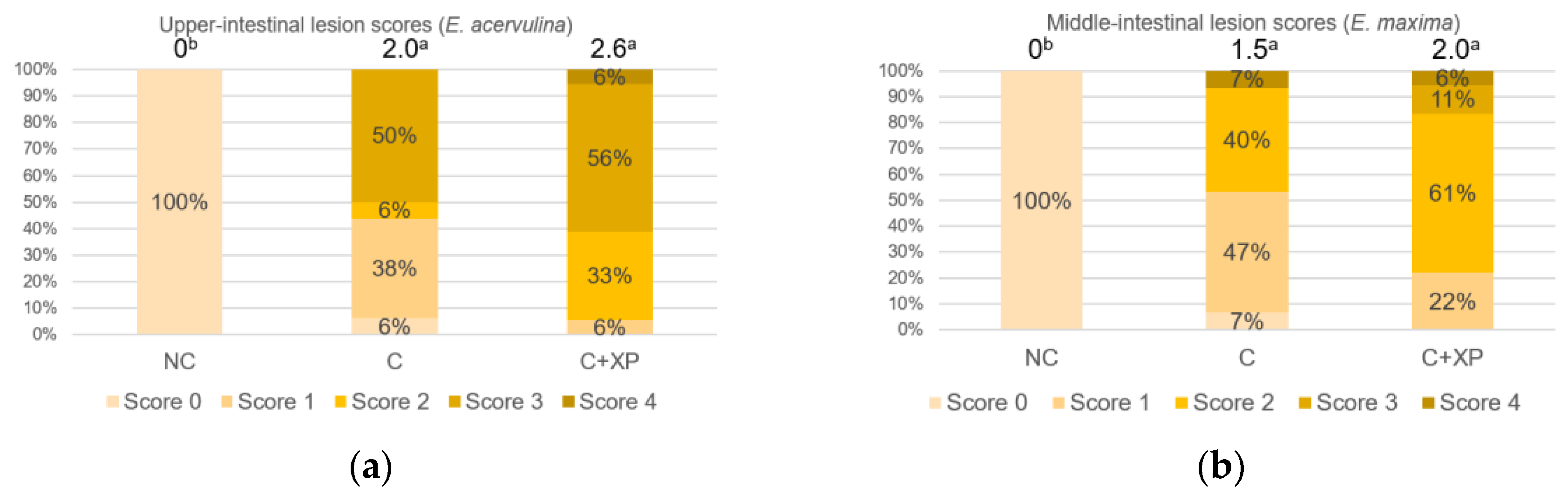
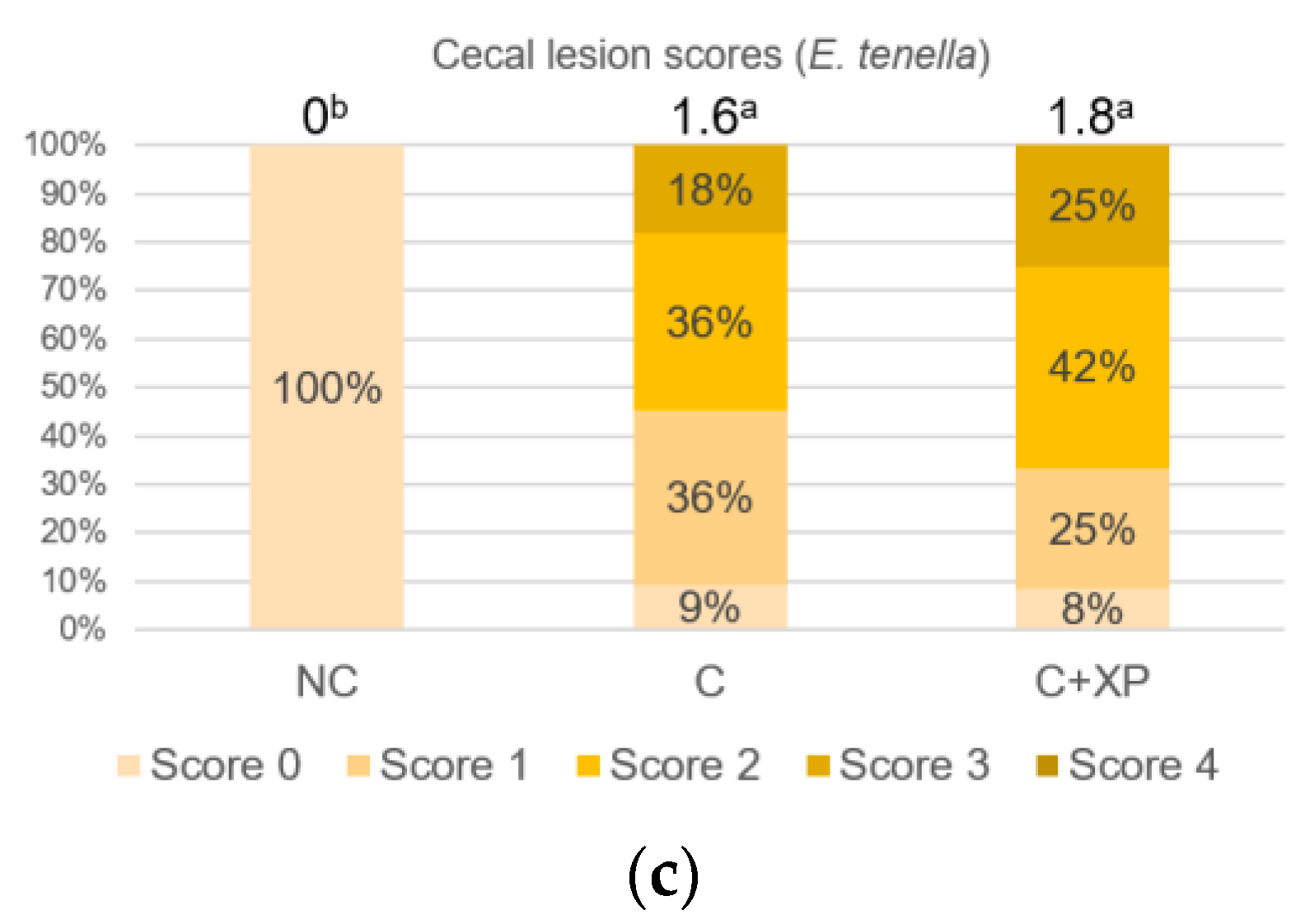
Eimeria challenge increases the permeability of the intestinal epithelium, and the extent of the damage can be assessed using fluorescein isothiocyanate-dextran (FITC-d). Higher levels of FITC-d in serum indicate greater FITC-d leakage from the intestine, revealing the severity of impairment of tight junction barriers [
29]. Upon entering the host intestine, sporulated oocysts begin several cycles of asexual multiplication before sexual multiplication. During the asexual multiplication stages, a mass of merozoites is produced and penetrates the epithelial cells of the host, resulting in severe intestinal impairment and permeability defects [
3]. In the current study, the levels of FITC-d detected in bird serum were greater in challenged birds compared with non-challenged birds. However, enzyme supplementation had no effect on gut leakage. The observation was likely because enzymes cannot directly target
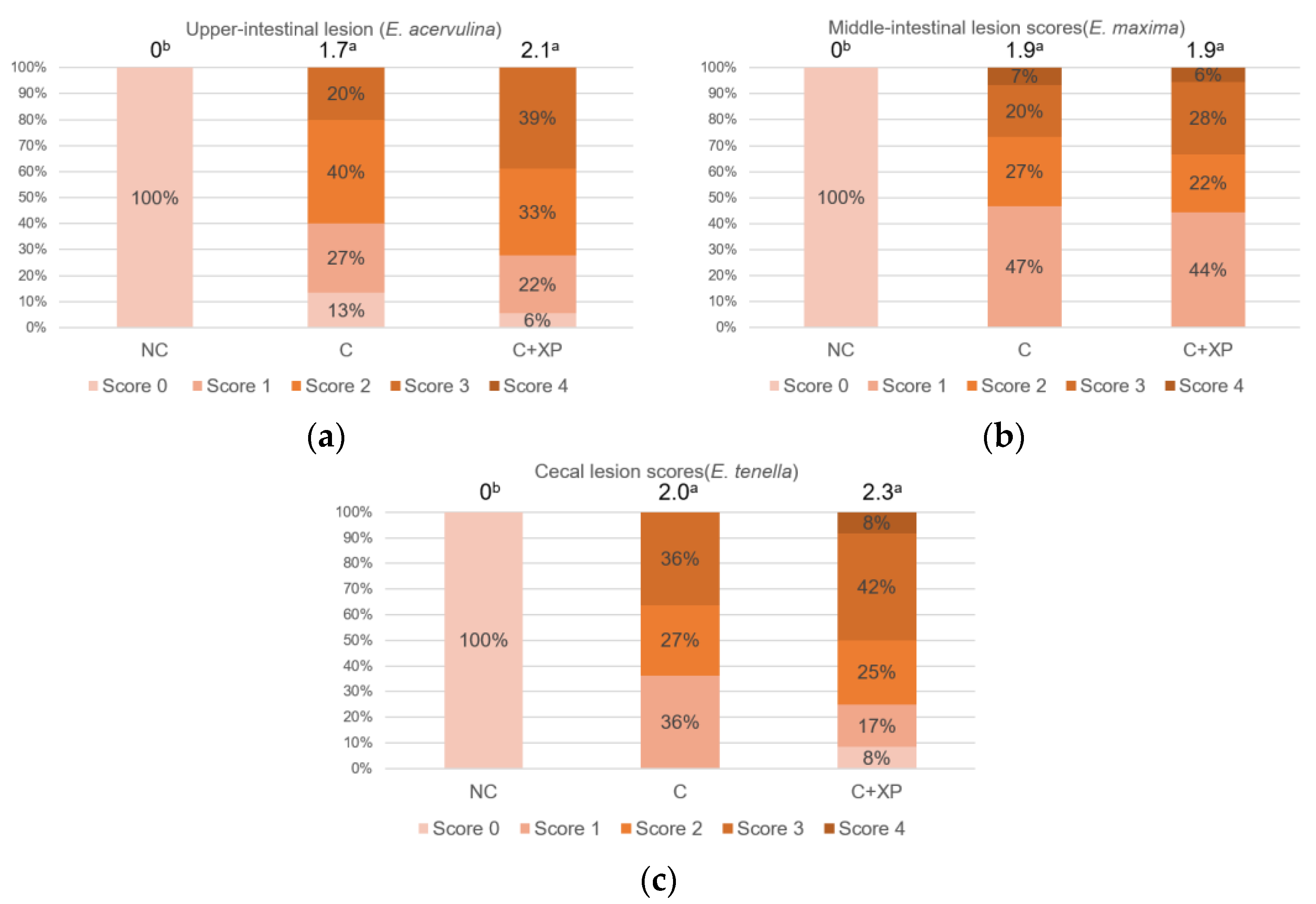
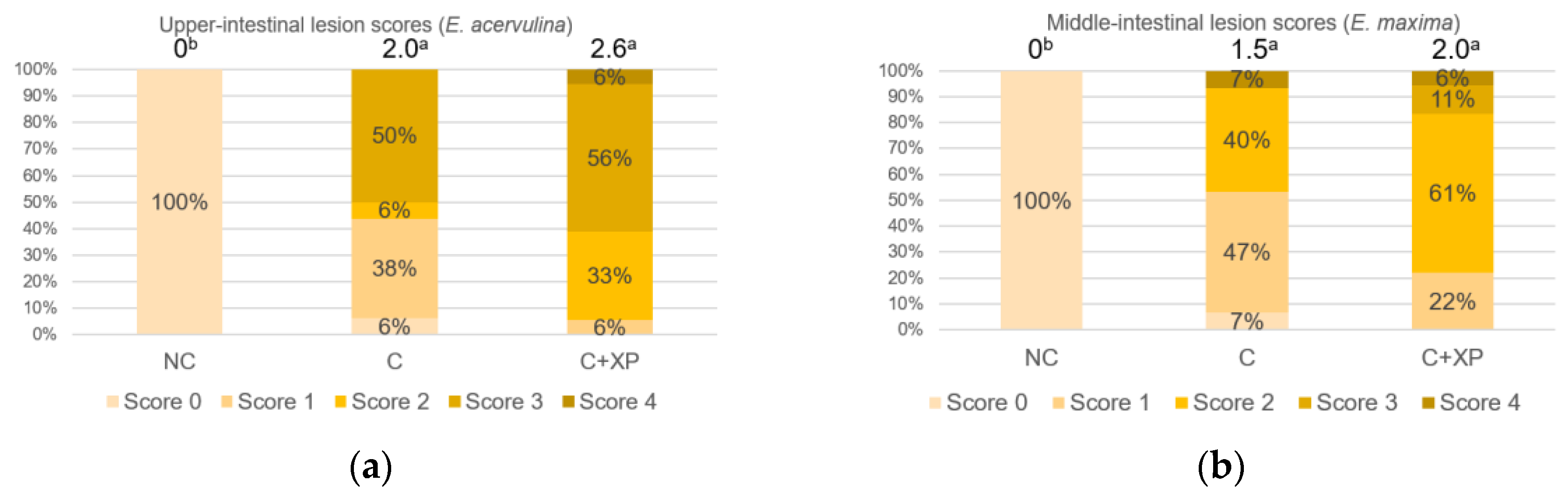
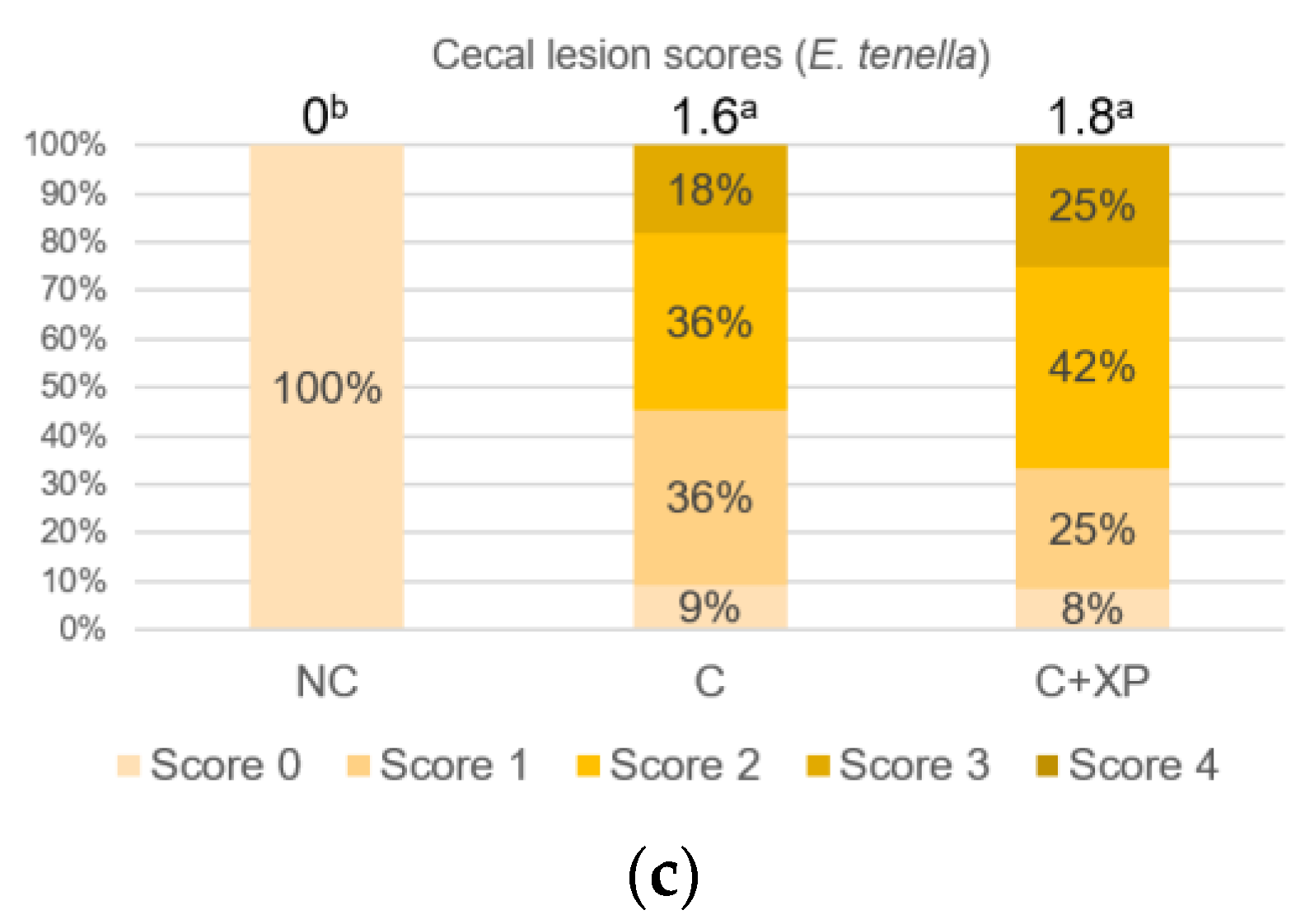
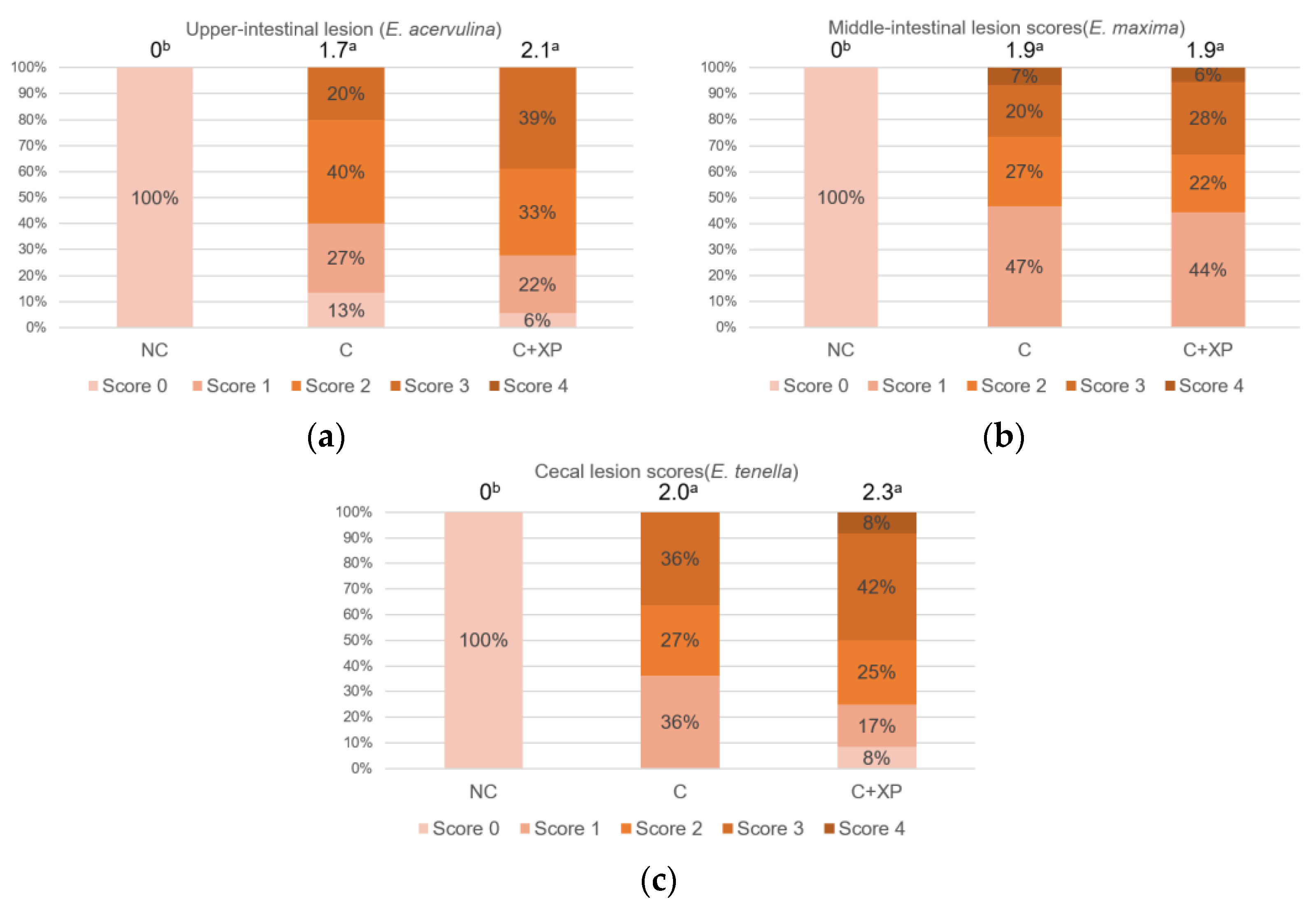
Eimeria oocyst nor reduce its multiplication, which is the primary cause of cell destruction and gut leakage. For the same reason, there was no lesion score reduction observed in enzyme-supplemented treatments.
On the other hand, observation in the current experiments on the expression of tight junction proteins indicates that the enzymes supplementation promoted intestinal integrity. Tight junctions are multi-functional protein complexes principally consisting of three families of transmembrane proteins: claudins, occludins, and junctional adhesion molecules. These complexes act as guards to seal the paracellular space between adjacent epithelial cells, regulating nutrient absorption and restricting pathogen entry. Claudin family proteins constitute the tight junction structural framework and each protein plays a specific function [
30]. In the current study, in response to the mixed
Eimeria challenge, the expression of claudin-1 at 6 DPI was significantly increased. The same change of claudin-1 was reported in previous coccidia studies [
21,
31]. The key inflammatory cytokine TNFα led to the increase in claudin-1 in IEC-18 cells, and increased expression of claudin-1 was observed in ulcerative colitis. It is speculated that the increased claudin-1 is related to inflammatory bowel disease [
32,
33,
34]. When attacked by inflammatory pathogens, the intestinal paracellular structure, as well as osmotic balance, are damaged, requiring a mass replacement of barrier-forming proteins including claudin-1. This intensive replacement is clinically observed as diarrhea. This replacement can not only renew intestinal barriers but also protectively exclude invasive pathogens [
34].
Eimeria spp. multiplies by invading and breaking down intestinal cells intercellularly and intracellularly. All of these
Eimeria-induced activities stimulate the replacement of tight junctions, consequently increasing the expression of claudin-1, which was also observed in the current study. The data in the current study demonstrated the combined effect of xylanase and protease on decreasing the claudin-1 expression. It can be reasoned that the enzymes helped to partly alleviate
Eimeria-induced intestinal barrier impairment and thus helped reduce the metabolic costs associated with the disease challenge. Similar findings were reported in animals receiving xylanase supplemented diet when challenged with Clostridium perfringens [
35,
36].
Eimeria infection causes the downregulation of nutrient transporters, which consequently contributes to possible depressed nutrient utilization and ultimately diminished growth performance [
37]. In the current study,
Eimeria infection significantly downregulated the expression of sugar transporters GLUT2 and GLUT5 in broilers fed HFAP diets (Expt. 1), although the decrease was rather nominal for birds receiving LFLP diets in Expt. 2. GLUT5, located on the apical side, is a facilitated-diffusion fructose transporter that facilitates the uptake of fructose into enterocytes. GLUT2, located at the basolateral side, takes over the sugar absorption transport once inside the cell and mediates the passive transport of glucose, fructose, and galactose out of enterocytes into the bloodstream [
38,
39]. Others have similarly observed the downregulation of intestinal GLUT2, GLUT5, and SGLT1 in
Eimeria- and
Campylobacter jejuni-challenged birds [
7,
22,
37,
40]. In the current study, amino acid transporters CAT2 and y+LAT1 were also downregulated in
Eimeria-challenged groups regardless of the dietary fiber and protein levels. This observation is consistent with previous observations that
E. maxima caused ileal and
E. tenella caused jejunal CAT2/y+LAT1 downregulation [
37]. CAT2 and y+LAT1 are basolateral amino acid transporters mediating the transport of cationic and neutral amino acids [
41]. As important L-arginine transporters, the depression of CAT2 and y+LAT1 may contribute to the reported sharp drop of plasma arginine in
E. acervulina challenged chickens [
42].
The downregulation of nutrient transporter proteins caused by
Eimeria challenge affects a wide range of sugar and amino acid transporter proteins such as GLUT2, GLUT5, SGLT1, PepT1, b0AT, b0+AT, EAAT3, rBAT, y+LAT2 [
6,
7,
37,
43]. The causes for the downregulation effect are not clear, but it is speculated that this response is a cell-mediated protective reaction in response to pathogenic invasion. By downregulating nutrient transporters in epithelial cells, a depleted nutritional environment is created to limit the development of parasites. In addition, the malnourished cells may trigger apoptosis and consequent epithelial renewal [
6,
37]. The interaction between
Eimeria challenge and enzyme supplementation in HFAP diets (Expt. 1) showed that compared to the non-challenged treatments, enzyme supplementation upregulated the expression of GLUT2 and GLUT5 to possibly compensate for nutrient utilization deficit. The xylanase-mediated nutrient transporter upregulation was reported in broiler chickens challenged with
Clostridium perfringens [
35,
44]. These observations suggest the potential of exogenous enzymes to alleviate some negative effects of
Eimeria challenge in broiler chickens.
Exogenous enzymes or dietary fiber have been shown to influence the content and profile of oligosaccharides in the digestive tract [
15,
17,
45,
46]. Some of these oligosaccharides could serve prebiotic functions in birds. In a study by Lin and Olukosi [
17], broiler chickens receiving diets with higher fiber content had a greater content of jejunal pentose-oligosaccharides as well as cecal SCFA. In the current study, the concentration of jejunal hexose-oligosaccharides was reduced in the challenged birds receiving LFLP diets (Expt. 2), which is likely due to the depressed feed intake in challenged birds. On the other hand, no significant difference was observed in the jejunal oligosaccharides content in the different HFAP treatments in Expt. 1. Interestingly, the
Eimeria challenge increased the cecal concentration of BCFA which exclusively originates from protein fermentation, but decreased the concentration of acetate and butyrate which mainly originate from carbohydrates fermentation, regardless of the types of diet profile in the two experiments. The production of BCFA is considered a marker for estimating cecal protein fermentation [
47], thus it can be speculated that
Eimeria promoted cecal protein fermentation but inhibited the fermentation of carbohydrates. This is possibly due to a change in the proportion of protein and carbohydrates reaching the hindgut for fermentation or a shift in the microbial population inhabiting the hindgut of challenged, compared with non-challenged birds.
It is widely acknowledged that hindgut fermentation of carbohydrates is generally more beneficial than protein fermentation [
48]. For example, butyrate resulting largely from carbohydrate fermentation is regarded as an energy source for enterocytes. Butyrate also plays a role in reducing inflammation and oxidative stress and in enhancing the colonic defense barrier. The supplementation of butyrate was reported to control coccidiosis [
49]. The greater ceca BCFA content observed in
Eimeria-challenged chickens may be explained by the observed downregulation in amino acid transporters. This may lead to impaired amino acids absorption in the small intestine of the host, and consequently leading to disproportionally large amounts of protein in the hindgut. Secondly, large amounts of epithelial cell debris produced by coccidiosis-induced intestinal damage ultimately flow to the ceca as potentially fermentable protein substrates. In addition, an
Eimeria-induced cecal microflora shift has been indicated in previous studies [
49], leading to an increase in Firmicutes (mucins and amino acids fermentation) and Proteobacteria (amino acids fermentation) and a decrease in Bacteroidetes (carbohydrates fermentation) [
50,
51,
52]. Therefore, protein fermentation is promoted at the expense of carbohydrates fermentation. Instructively though, in the current study, enzyme supplementation mitigated the
Eimeria-induced enhancement of cecal BCFA in HFAP diets (Expt. 1). These observations demonstrated a potential positive effect of enzyme supplementation in alleviating the shift in microbial fermentation patterns caused by
Eimeria challenge.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}