
Chromosome Diversity and Evolution in Helicoide a (Gastropoda: Stylommatophora): A Synthesis from Original and Literature Data


 ,
,  ,
,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Molecular Analysis
2.3. Cytogenetic Analysis
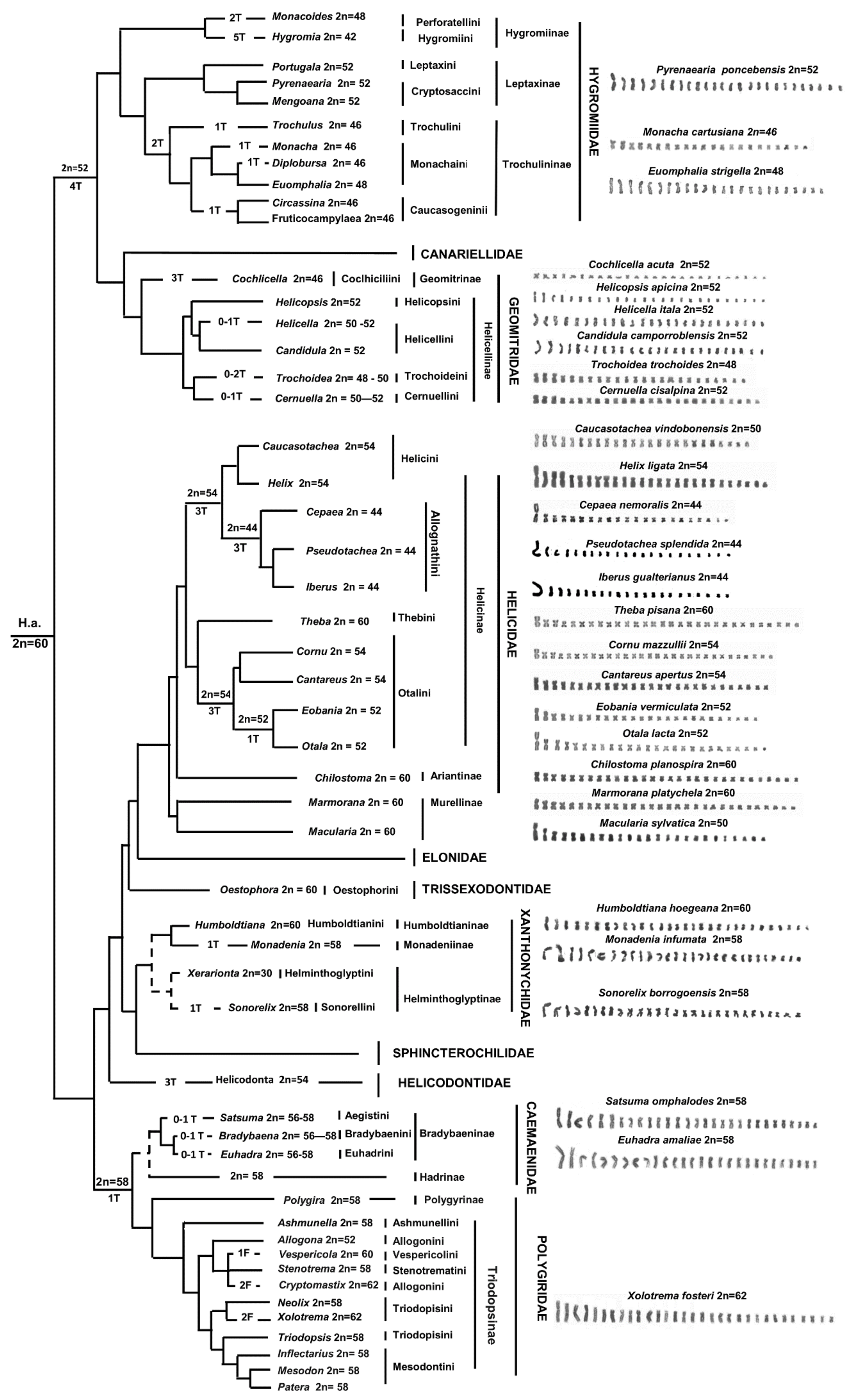
3. Results
3.1. Molecular Analysis and Taxonomic Attribution
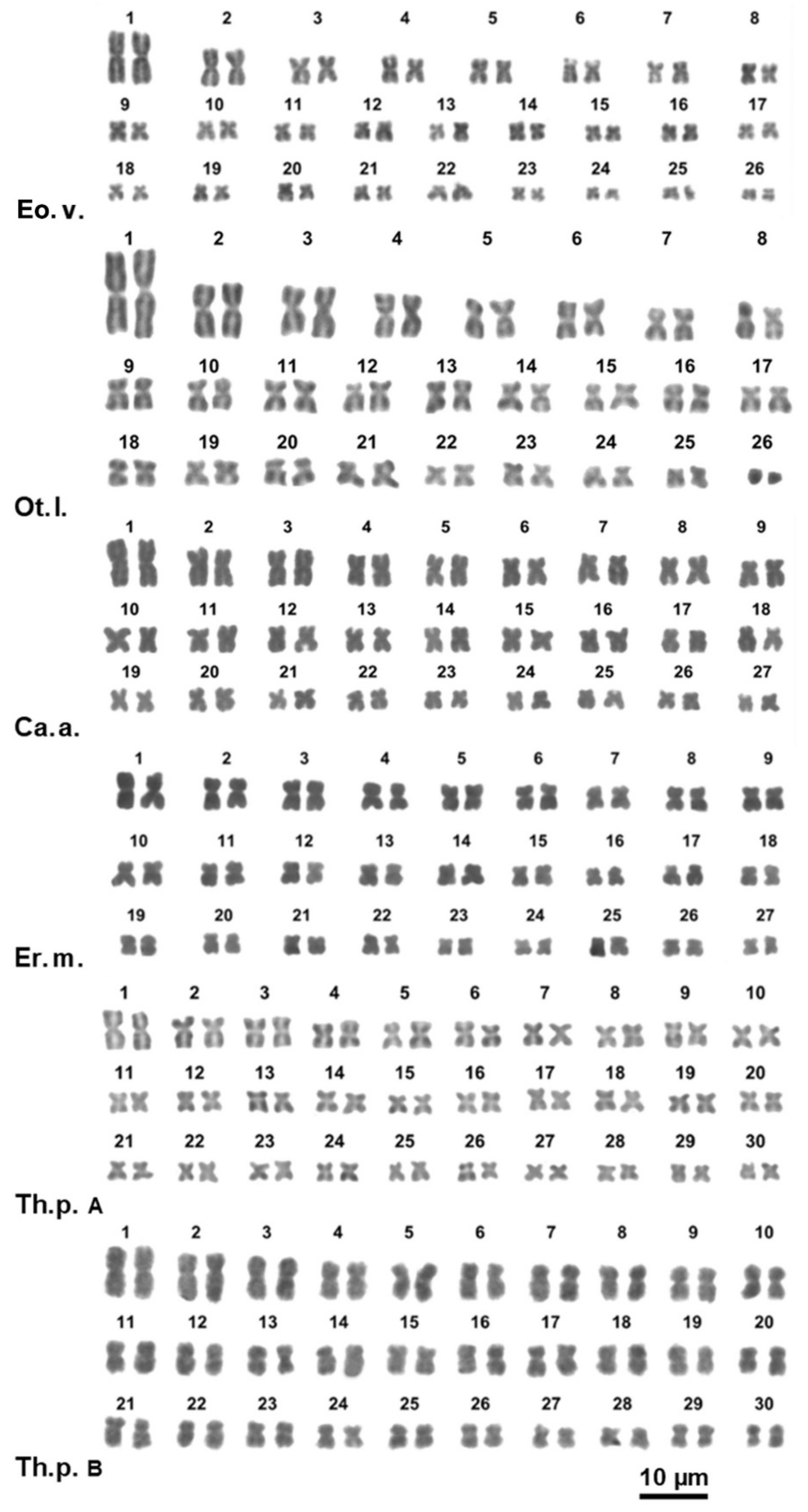
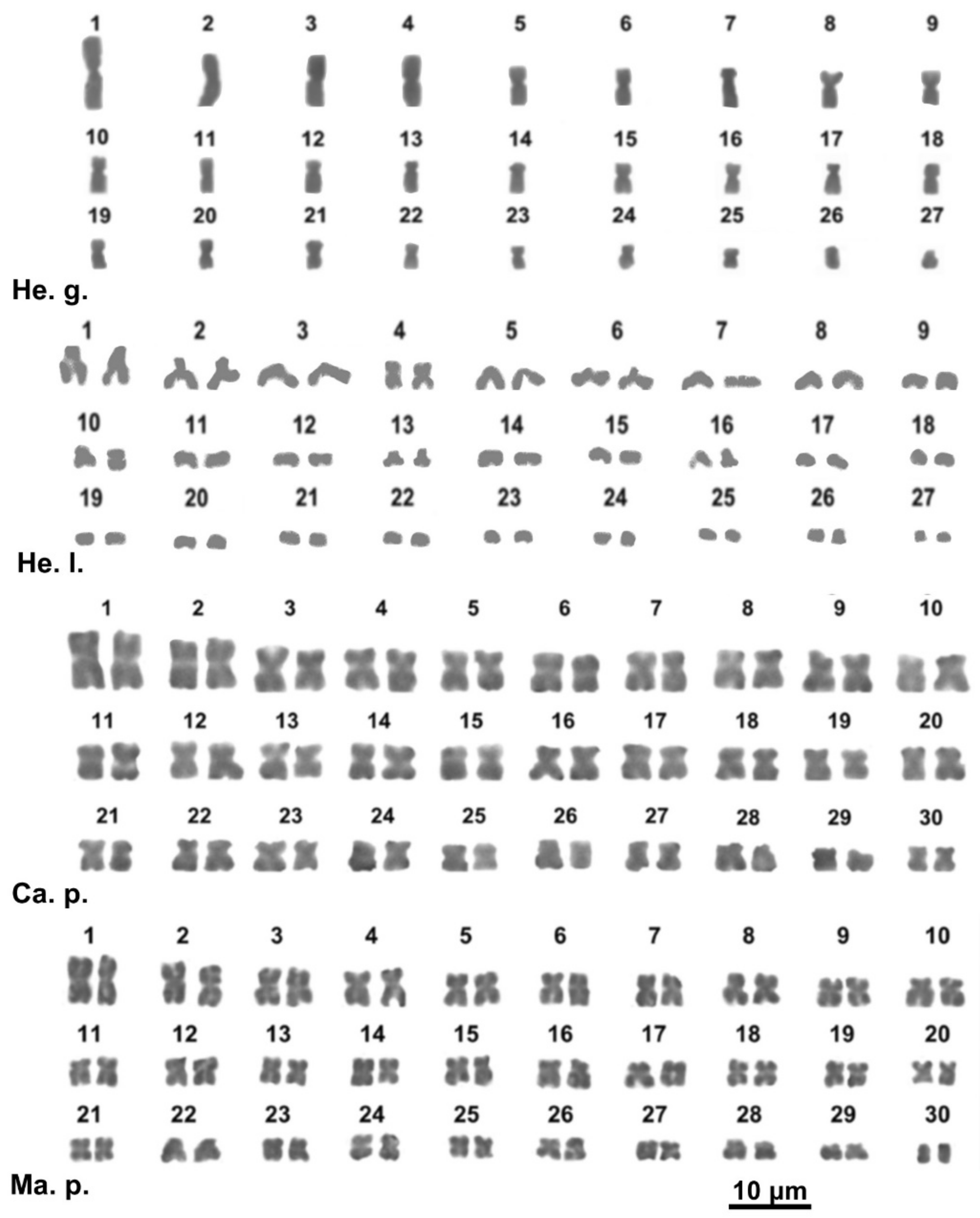
3.2. Karyotype Description
3.2.1. Family Higromiidae
3.2.2. Family Geomitridae
3.2.3. Family Helicidae
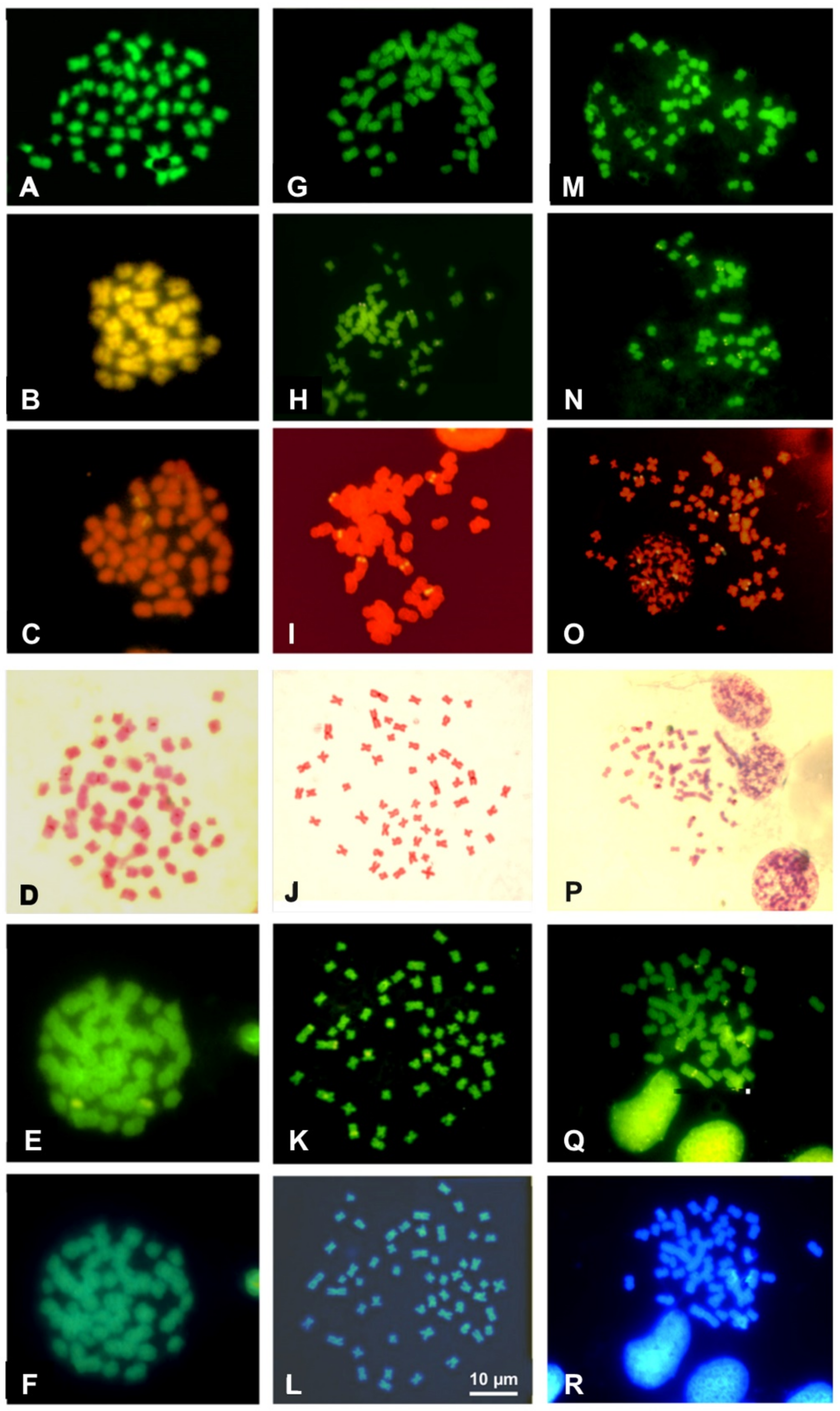
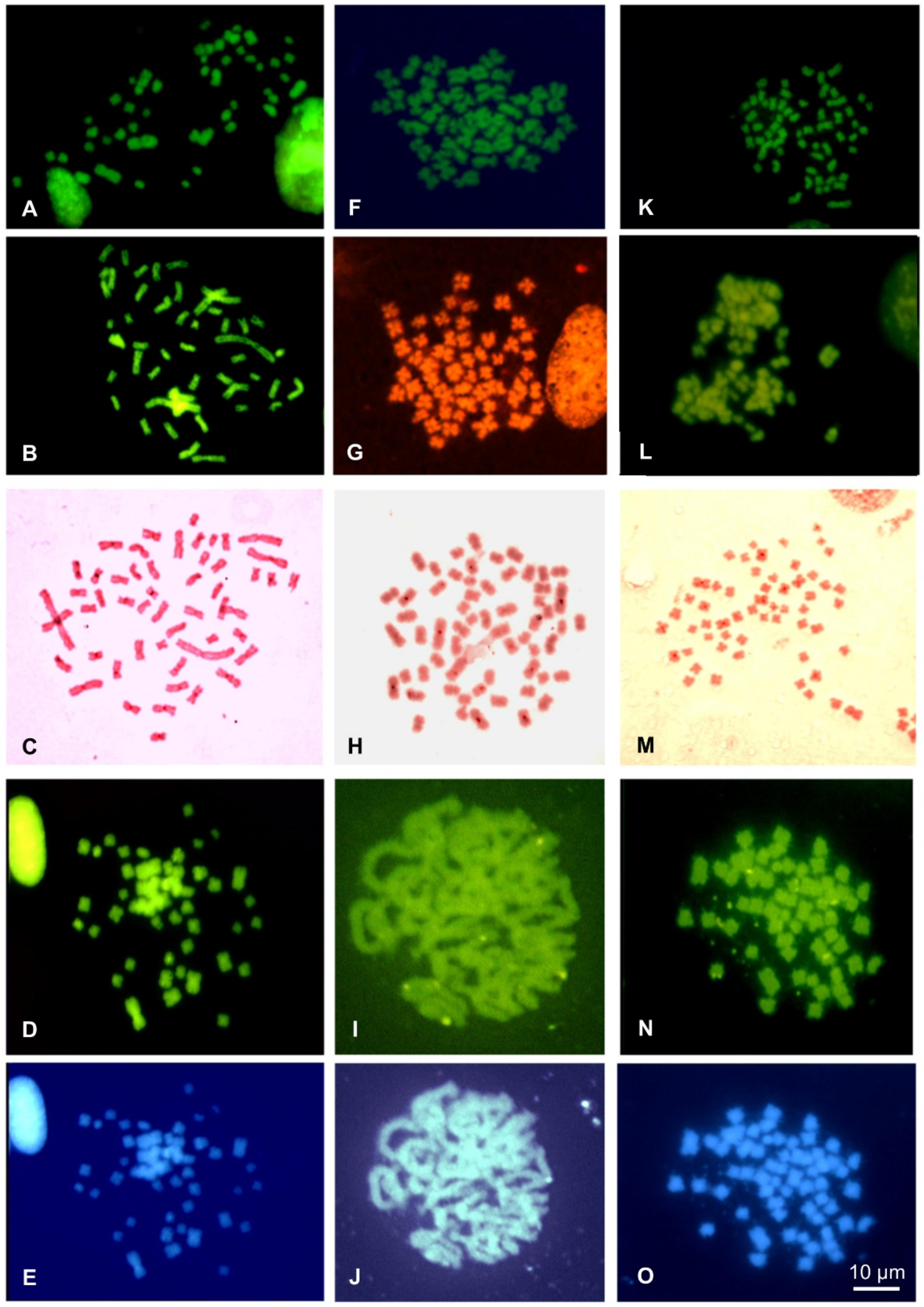
3.3. Chromosome Banding and NOR-FISH
4. Discussion
4.1. Molecular Analysis and Taxonomic Attribution
4.2. Chromosome Analysis and Karyotype Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WoRMS Editorial Board. World Register of Marine Species. 2021. Available online: http://www.marinespecies.org (accessed on 30 June 2021).
- Mollusca Base. 2021. Available online: http://www.molluscabase.org (accessed on 30 June 2021).
- Scott, B. Phylogenetic relationships of the Camaenidae (Pulmonata, Stylommatophora, Helicoidea). J. Molluscan Stud. 1996, 62, 65–74. [Google Scholar] [CrossRef]
- Scott, B. Biogeography of the Helicoidea (Mollusca, Gastropoda, Pulmonata), land snails with a Pangean distribution. J. Biogeogr. 1997, 24, 399–407. [Google Scholar] [CrossRef]
- Schileyko, A.A. Treatise on recent terrestrial pulmonate molluscs. Part 12. Bradybaenidae, Monadeniidae, Xanthonychidae, Epiphragmophoridae, Helminthoglyptidae, Elonidae, Humboldtianidae, Sphincterochilidae, Cochlicellidae. Ruthenica 2004, 2, 1627–1763. [Google Scholar]
- Schileyko, A.A. Treatise on recent terrestrial pulmonate molluscs. Part 13. Helicidae, Pleurodontidae, Polygyridae, Ammonitellidae, Oreohelicidae, Thysanophoridae. Ruthenica 2006, 2, 1765–1906. [Google Scholar]
- Schileyko, A.A. Treatise on recent terrestrial pulmonate molluscs. Part 14. Helicodontidae, Ciliellidae, Hygromiidae. Ruthenica 2006, 2, 1907–2047. [Google Scholar]
- Bouchet, P.; Rocroi, J.P. Classification and nomenclature of gastropod families. Malacologia 2005, 47, 1–397. [Google Scholar]
- Razkin, O.; Gómez-Moliner, B.J.; Prieto, C.E.; Martínez-Ortí, A.; Arrébola, J.R.; Muñoz, B.; Chueca, L.J.; Madeira, M.J. Molecular phylogeny of the western Palaearctic Helicoidea (Gastropoda, Stylommatophora). Mol. Phylogenet. Evol. 2015, 83, 99–117. [Google Scholar] [CrossRef]
- Perez, K.E.; Defreitas, N.; Slapcinsky, J.; Minton, R.L.; Anderson, F.E.; Pearcet, A. Molecular phylogeny, evolution of shell shape, and DNA barcoding in Polygyridae (Gastropoda, Pulmonata), an endemic North American clade of land snails. Am. Malacol. Bull. 2014, 32, 1–31. [Google Scholar] [CrossRef]
- Köhler, F.; Criscione, F. A molecular phylogeny of camaenid land snails from north-western Australia unravels widespread homoplasy in morphological characters (Gastropoda, Helicoidea). Mol. Phylogenet. Evol. 2015, 83, 44–55. [Google Scholar] [CrossRef]
- Wade, C.M.; Mordan, P.B.; Naggs, F. Evolutionary relationships among the Pulmonate land snails and slugs (Pulmonata, Stylommatophora). Biol. J. Linn. Soc. 2016, 87, 593–610. [Google Scholar] [CrossRef]
- Caro, A.; Neiber, M.T.; Gomez-Moliner, B.J.; Madeira, M.J. Molecular phylogeny and biogeography of the land snail subfamily Leptaxinae (Gastropoda, Hygromiidae). Mol. Phylogenet. Evol. 2019, 139, 106570. [Google Scholar] [CrossRef]
- Groenenberg, D.S.J.; Subai, P.; Gittenberger, E. Systematics of Ariantinae (Gastropoda, Pulmonata, Helicidae), a new approach to an old problem. Contrib. Zool. 2016, 85, 37–65. [Google Scholar] [CrossRef]
- Tatewaki, R.; Kitada, J. Karyological studies of five species of land snails (Helicoidea, Mollusca). Genetica 2004, 74, 73–80. [Google Scholar] [CrossRef]
- Thiriot-Quiévreux, C. Advances in chromosomal studies of gastropod molluscs. J. Molluscan Stud. 2003, 69, 187–201. [Google Scholar] [CrossRef]
- Patterson, C.M. Chromosomes of molluscs. In Proceedings of the 2nd Symposium of Mollusca, Cochin, India, 12–16 January 1968; Series 3, Part 2. Marine Biological Association of India: Ernakulam, India, 1968; pp. 635–686. [Google Scholar]
- Husted, L.; Burch, P.R. The Chromosomes of Polygyrid Snails. Am. Nat. 1946, 80, 410–429. Available online: http://www.jstor.org/stable/2458185 (accessed on 18 August 2021). [CrossRef] [PubMed]
- Laws, H.M. Chromosomes of snails introduced into South Australia and the Northern Territory. Rec. Aust. Mus. 1965, 15, 79–87. [Google Scholar]
- Bantock, C.R. Localisation of chiasmata in Cepaea nemoralis L. Heredity 1972, 29, 213–221. [Google Scholar] [CrossRef][Green Version]
- Laws, H.M. The chromosomes of some Australian camaenid land snails. Cytologia 1973, 38, 229–235. [Google Scholar] [CrossRef][Green Version]
- Stern, E.M.; Metcalf, A. L Chromosome Numbers in Ashmunella (Gastropoda Eupulmonata Polygyridae). Veliger 1974, 17, 19–22. [Google Scholar]
- Babrakzai, N. Chromosomes of Helminthoglyptidae (Pulmonata: Gastropoda). Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 1975. [Google Scholar]
- Page, C. The Karyotype of the Land Snail Cepaea nemoralis (L.). Heredity 1978, 41, 321–325. [Google Scholar] [CrossRef]
- Aparicio, M.T. Cytotaxonomic studies of the family Helicidae (Gastropoda, Pulmonata). Genét. Ibér. 1981, 33, 211–224. [Google Scholar]
- Mezzasalma, M.; Guarino, F.M.; Aprea, G.; Petraccioli, A.; Crottini, A.; Odierna, G. Karyological evidence for diversification of Italian slow worm populations. Comp. Cytogenet. 2013, 7, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Mezzasalma, M.; Andreone, F.; Branch, W.R.; Glaw, F.; Guarino, F.M.; Nagy, Z.T.; Odierna, G.; Aprea, G. Chromosome evolution in pseudoxyrhophiine snakes from Madagascar, a wide range of karyotypic variability. Biol. J. Linn. Soc. 2014, 112, 450–460. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Andreone, F.; Aprea, G.; Glaw, F.; Odierna, G.; Guarino, F.M. Molecular phylogeny, biogeography and chromosome evolution of Malagasy dwarf geckos of the genus Lygodactylus (Squamata, Gekkonidae). Zool. Scr. 2017, 46, 42–54. [Google Scholar] [CrossRef]
- Odierna, G.; Aprea, G.; Barucca, M.; Biscotti, M.A.; Canapa, A.; Capriglione, T.; Olmo, E. Karyology of the Antarctic chiton Nuttalochiton mirandus (Thiele, 1906) (Mollusca, Polyplacophora) with some considerations on chromosome evolution in chitons. Chromosome Res. 2008, 16, 899–906. [Google Scholar] [CrossRef]
- Petraccioli, A.; Maio, N.; Odierna, G. Chromosomes of Lepidochitona caprearum (Scacchi, 1836) (Polyplacophora, Acanthochitonina, Tonicellidae) provide insights into Acanthochitonina karyological evolution. Comp. Cytogenet. 2012, 6, 397–407. [Google Scholar] [CrossRef]
- Petraccioli, A.; Capriglione, T.; Colomba, M.; Crovato, P.; Odierna, G.; Sparacio, I.; Maio, N. Comparative cytogenetic study in four alopiinae door snails (Gastropoda, Clausiliidae). Malacologia 2015, 58, 225–232. [Google Scholar] [CrossRef]
- Fiorentino, V.; Salomone, N.; Manganelli, G.; Giusti, F. Historical biogeography of Tyrrhenian land snails, the Marmorana-Tyrrheniberus radiation (Eupulmonata, Helicidae). Mol. Phylogenet. Evol. 2010, 55, 26–37. [Google Scholar] [CrossRef]
- Fiorentino, V.; Manganelli, G.; Giusti, F.; Ketmaier, V. Recent expansion and relic survival, phylogeography of the land snail genus Helix (Mollusca, Gastropoda) from south to north Europe. Mol. Phylogenet. Evol. 2016, 98, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Dayrat, B.; Conrad, M.; Balayan, S.; White, T.R.; Albrecht, C.; Golding, R.; Gomes, S.R.; Harasewych, M.G.; Martins, A.M.D.F. Phylogenetic relationships and evolution of pulmonate gastropods (Mollusca), new insights from increased taxon sampling. Mol. Phylogenet. Evol. 2011, 59, 425–437. [Google Scholar] [CrossRef]
- Giusti, F. Notulae malacologicae XVI. I molluschi terrestri e di acqua dolce viventi sul massiccio dei Monti Reatini (Appennino Centrale). Lav. Soc. Ital. Biogeog. 1971, 2, 423–576. [Google Scholar] [CrossRef]
- Giusti, F.; Castagnolo, L. I molluschi terrestri delle dune italiane, brevi cenni di ecologia, elenco delle specie e chiavi per il loro riconoscimento. In Quaderni Sulla “Struttura delle Zoocenosi Terrestri”, 3. Ambienti Mediterranei, 1. Le Coste Sabbiose; Pubblicazioni AQ/1/174; C.N.R.: Roma, Italy, 1982; pp. 51–102. [Google Scholar]
- Giusti, F.; Castagnolo, L.; Manganelli, G. La fauna malacologica delle faggete Italiane, brevi cenni di ecologia, elenco delle specie e chiavi per il riconoscimento dei generi e delle entità più comuni. Boll. Malacol. 1985, 21, 69–144. [Google Scholar]
- Kerney, M.P.; Cameron, R.A.D. Guide des Escargots et limaces d’Europe Identification et Biologie de Plus de 300 Espèces; Delachaux et Niestlé: Paris, France, 1999. [Google Scholar]
- Gavetti, E.; Birindelli, S.; Bodon, M. Molluschi terrestri e d’acqua dolce della valle di Susa. Monogr. Mus. Reg. Sci. Nat. Torino 2008, 44, 1–273. [Google Scholar]
- Welter-Schultes, F.W. European Non-Marine Molluscs, a Guide for Species Identification; Planet Poster Editions: Göttingen, Germany, 2012. [Google Scholar]
- Maio, N.; Petraccioli, A.; Crovato, P.; Niero, I.; Pignataro, C.; Odierna, G. Guida naturalistica di campo ai Molluschi terrestri dei Monti Alburni. Biologia, ecologia, distribuzione e conservazione. PNCVDA Quad. Della Biodivers. 2016, 4, 1–354. [Google Scholar]
- Sokolov, E.P. An improved method for DNA isolation from mucopolysaccharide-rich molluscan tissues. J. Molluscan Stud. 2000, 66, 573–575. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Martin, A.; Romano, S.; Mcmillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Version 2.0; University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Hall, T.A. BioEdit, a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Sidhom, M.; Said, K.; Chatti, N.; Guarino, F.M.; Odierna, G.; Petraccioli, A.; Picariello, O.; Mezzasalma, M. Karyological characterization of the common chameleon (Chamaeleo chamaeleon) provides insights on the evolution and diversification of sex chromosomes in Chamaeleonidae. Zoology 2020, 141, 125738. [Google Scholar] [CrossRef]
- Schmid, M. Chromosome banding in Amphibia. Constitutive heterochromatin and nucleolus organizer regions in Bufo and Hyla. Chromosoma 1978, 66, 361–388. [Google Scholar] [CrossRef]
- Sahar, E.; Latt, S.A. Energy transfer and binding competition between dyes used to enhance staining differentiation in metaphase chromosomes. Chromosoma 1980, 79, 1–28. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Odierna, G.; Aprea, G.; Capriglione, T. Chromosomal and molecular analysis of some repeated families in Discoglossus Otth, 1837 (Anura, Discoglossidae), taxonomic and phylogenetic implications. Ital. J. Zool. 1999, 66, 275–283. [Google Scholar] [CrossRef][Green Version]
- Mezzasalma, M.; Visone, V.; Petraccioli, A.; Odierna, G.; Capriglione, T.; Guarino, F.M. Non-random accumulation of LINE1-like sequences on differentiated snake W chromosomes. J. Zool. 2016, 300, 67–75. [Google Scholar] [CrossRef]
- Levan, A.R.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ezzine, I.K.; Dimassi, N.; Pfarrer, B.; Said, K.; Neubert, E. New records of the endemic Sicilian land snail species Marmorana (Murella) muralis (O. F. Muller, 1774) from the north of Tunisia. (Eupulmonata, Gastropoda). Zookeys 2018, 775, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, G.; Salomone, N.; Giusti, F. A molecular approach to the phylogenetic relationships of the western Palaearctic Helicoidea (Gastropoda, Stylommatophora). Biol. J. Linn. Soc. 2005, 85, 501–512. [Google Scholar] [CrossRef]
- Boeckers, A.; Greve, C.; Hutterer, R.; Misof, B.; Haase, M. Testing heterogeneous base composition as potential cause for conflicting phylogenetic signal between mitochondrial and nuclear DNA in the land snail genus Theba Risso 1826 (Gastropoda, Stylommatophora, Helicoidea). Org. Divers. Evol. 2016, 16, 835–846. [Google Scholar] [CrossRef]
- Steinke, D.; Albrecht, C.; Pfenninger, M. Molecular phylogeny and character evolution in the Western Palaearctic Helicidae s.l. (Gastropoda, Stylommatophora). Mol. Phylogenet. Evol. 2004, 32, 724–734. [Google Scholar] [CrossRef]
- Neiber, M.T.; Hausdorf, B. Phylogeography of the land snail genus Circassina (Gastropoda, Hygromiidae) implies multiple Pleistocene refugia in the western Caucasus region. Mol. Phylogenet. Evol. 2015, 93, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Pienkowska, J.R.; Manganelli, G.; Giusti, F.; Lesicki, A. Monacha claustralis (Rossmässler, 1834) new to Polish and Czech malacofauna (Gastropoda, Eupulmonata, Hygromiidae). J. Conchol. 2015, 42, 79–93. [Google Scholar]
- Pienkowska, J.R.; Prockow, M.; Gorka, M.; Lesicki, A. Distribution of Monacha claustralis (Rossmässler, 1834) and M. cartusiana (O. F. Mueller, 1774) (Eupulmonata, Hygromiidae) in central European and Balkan countries, new data. Folia Malacol. 2018, 26, 103–120. [Google Scholar] [CrossRef]
- Patterson, C.M.; Burch, J.B. Chromosomes of Pulmonated Molluscs. In Pulmonates; Systematics, Evolution and Ecology; Fretter, V., Peake., J., Eds.; Academic Press: New York, NY, USA, 1978; Volume 2A, pp. 171–217. [Google Scholar]
- Aparicio, M.T. The chromosomes of eight species of the subfamily Helicinae (Gastropoda, Pulmonata, Helicidae) from Spain. Malacol. Rev. 1983, 16, 71–78. [Google Scholar]
- Bakhtadze, N.G.; Chakvetadze, N.L.; Mumladze, L.J.; Bakhtadze, G.I.; Tskhadaia, E.A. Karyological data of terrestrial mollusks (Mollusca: Gastropoda: Pulmonata) of Georgia. Proc. Inst. Zool. 2014, 25, 23–27. [Google Scholar] [CrossRef]
- Vitturi, R.; Rasotto, M.B.; Farinella-Ferruzza, N. The chromosomes of 16 molluscan species. Boll. Zool. 1982, 49, 61–71. [Google Scholar] [CrossRef]
- Vitturi, R.; Libertini, A.; Sineo, L.; Sparacio, I.; Lannino, A.; Gregorini, A.; Colomba, M. Cytogenetics of the land snails Cantareus aspersus and C. mazzullii (Mollusca, Gastropoda, Eupulmonata). Micron 2005, 36, 351–357. [Google Scholar] [CrossRef]
- Sun, T. Chromosomal studies in three land snails. Sinozoologia 1995, 12, 154–162. [Google Scholar]
- Babrakzai, N.; Samsam, S.; Miller, W.B. Cytological Relationships of Helminthoglyptidae and Bradybaenidae (Mollusca, Pulmonata). In Proceedings of the Twenty-Fourth Annual Meeting of the Arizona-Nevada Academy of Science and the Fifty-Sixth Annual Meeting Southwestern and Rocky Mountain Division American Association for the Advancement of Science, Las Vegas, NV, USA, 9–12 April 1980; p. 31. [Google Scholar]
- Rainer, M. Chromosomenuntersuchungen an Gastropoden (Stylommatophora). Malacologia 1967, 5, 341–373. [Google Scholar]
- Petraccioli, A.; Crovato, P.; De Vico, G.; Odierna, G.; Picariello, O.L.A.; Tardy, E.; Viglietti, S.; Guarino, F.M.; Maio, N. Helix straminea Briganti, 1825 in Italy (Gastropoda, Pulmonata), taxonomic history, morphology, biology, distribution and phylogeny. Eur. Zool. J. 2021, 88, 390–416. [Google Scholar] [CrossRef]
- Odierna, G.; Olmo, E.; Cobror, O. Taxonomic implications of NOR-localization in lacertid lizards. Amphib. Reptil. 1987, 87, 373–382. [Google Scholar]
- Carvalho, K.A.; Garcia, P.C.A.; Recco-Pimentel, S.M. NOR dispersion, telomeric sequence detection in centromeric regions and meiotic multivalent configurations in species of the Aplastodiscus albofrenatus group (Anura, Hylidae). Cytogenet. Genome Res. 2009, 126, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Britton-Davidian, J.; Cazaux, B.; Catalan, J. Chromosomal dynamics of nucleolar organizer regions (NORs) in the house mouse, micro-evolutionary insights. Heredity 2012, 108, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Thiriot-Quiévreux, C. Review of the literature on bivalve cytogenetics in the last ten years. Cah. Biol. Mar. C 2002, 43, 17–26. [Google Scholar]
- Wang, Y.; Guo, X. Chromosomal rearrangement in Pectinidae revealed by rRNA loci and implications for bivalve evolution. Biol. Bull. 2004, 207, 247–256. [Google Scholar] [CrossRef]
- Colomba, M.S.; Gregorini, A.; Liberto, F.; Reitano, A.; Giglio, S.; Sparacio, I. Monographic revision of the endemic Helix mazzullii De Cristofori & Jan, 1832 complex from Sicily and re-introduction of the genus Erctella Monterosato, 1894 (Pulmonata, Stylommatophora, Helicidae). Zootaxa 2011, 3134, 1–42. [Google Scholar] [CrossRef]
- Colomba, M.S.; Vitturi, R.; Rampin, M.; Lannino, A.; Taravella, A.; Libertini, A. Comparative cytogenetic analysis of three Stylommatophoran slugs (Mollusca, Pulmonata). Malacologia 2009, 51, 173–179. [Google Scholar] [CrossRef]
- Vitturi, R.; Colombera, D.; Catalano, E.; Amico, F.P. Spermatocyte chromosome analysis of Helicella virgata (Eupulmonata, Helicidae). J. Hered. 1991, 82, 339–343. [Google Scholar] [CrossRef]
- Diopotex-Chong, M.E.; Babrazkai, N. Preliminary cytological studies in Triodopsis fosteri (Baker, 1920) Mollusca-Pulmonata Polygiridae. An. Inst. Cienc. Mar Limnol. 1993, 19, 13–18. [Google Scholar]
- Schweizer, D. Counterstain-enhanced chromosome banding. Hum. Genet. 1981, 57, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Odierna, G.; Aprea, G.; Barucca, M.; Canapa, A.; Capriglione, T.; Olmo, E. Karyology of the Antarctic scallop Adamussium colbecki, with some comments on the karyological evolution of pectinids. Genetica 2006, 127, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Odierna, G.; Aprea, G.; Barucca, M.; Canapa, A.; Capriglione, T.; Olmo, E. The karyology of the Antarctic whelk, Neobuccinum eatoni (Mollusca, Neogastropoda). Ital. J. Zool. 2006, 73, 303–308. [Google Scholar] [CrossRef]
- Petraccioli, A.; Guarino, F.M.; Maio, N.; Odierna, G. Molecular cytogenetic study of three common Mediterranean limpets, Patella caerulea, P. rustica and P. ulyssiponensis (Archaeogastropoda, Mollusca). Genetica 2010, 138, 219–225. [Google Scholar] [CrossRef] [PubMed][Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family/Subfam/Tribe | Species | Nr. and Origin of Samples | Voucher | Chr. Nr. |
|---|---|---|---|---|
| Hygromiidae | ||||
| Hygromiinae | ||||
| Monachaini | Monacha sp. | 1, Portici (Naples, Italy) | Gast 32 85.9% vs. KX495397 | 2n = 44 * |
| Trochoideini | Trochoidea elegans (Gmelin, 1791) | 3, Santa Severa (Rome, Italy) | Gast 193–195 99.3% vs. MG585435 | 2n = 48 * |
| Trochoidea pyramidata (Draparnaud, 1805) | 3, Capri (Naples, Italy) | Gast 184–186 93.9% vs. AY741444 | 2n = 48 * | |
| Trochoidea trochoides (Poiret, 1789) | 2, Fusaro (Naples, Italy) | Gast 91, 170 98.3% vs. AY546379 | 2n = 48 * | |
| Geomitridae | ||||
| Helicellinae | ||||
| Cernuellini | Cernuella virgata (Da Costa, 1778) | 2, Seiano (Naples, Italy) | Gast 354, 355 100% vs. KF250441 | 2n = 52 |
| Cochlicellini | Cochlicella acuta (O. F. Müller, 1774) | 2, Monte S.’Angelo (Naples, Italy) | Gast 342–343 100% vs. AY741443 | 2n = 52 |
| Helicidae | ||||
| Ariantinae | Campylaea planospira (Lamarck, 1822) | 2, Amalfi (Salerno, Italy) | Gast 202–203 | 2n = 60 * |
| Helicinae | ||||
| Helicini | Helix gussoneana L. Pfeiffer, 1848 | 1, Petina (Salerno, Italy) | Gast 149 99.7% vs. KU869969 | 2n = 54 * |
| Helix lucorum Linnaeus, 1758 | 2, Montellago (Venice, Italy) | Gast 352–353 99.3% vs. MG709101 | 2n = 54 | |
| Otalini | Cornu apertus (Born, 1778) | 2, Frignano (Caserta, Italy) | Gast 357–358 97.7% vs. KU870010 | 2n = 54 |
| Eobania vermiculata (O. F. Müller, 1774) | 1, Capri (Naples, Italy) | Gast 356 | 2n = 52 | |
| Erctella mazzullii (De Cristofori & Jan, 1832) | 1, Palermo (Italy) | Gast 67 99.5% vs. GQ402415 | 2n = 54 | |
| Otala lactea (O. F. Müller, 1774) | 2, Morocco | Gast 23–24 100% vs. MK603015 | 2n = 52 | |
| Thebini | Theba pisana (O. F. Müller, 1774) | 2, Fusaro (Naples, Italy) | Gast 87, 172 98.3% vs. AY741415 | 2n = 60 |
| Theba pisana (O. F. Müller, 1774) | 1, Messina (Italy) | Gast 77 99.2% vs. KU521652 | 2n = 60 | |
| Murellinae | Marmorana platychela (Menke, 1830) | 2, Palermo (Italy) | Gast 66, 108 100% vs. MG774447 | 2n = 60 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petraccioli, A.; Crovato, P.; Guarino, F.M.; Mezzasalma, M.; Odierna, G.; Picariello, O.; Maio, N. Chromosome Diversity and Evolution in Helicoide a (Gastropoda: Stylommatophora): A Synthesis from Original and Literature Data. Animals 2021, 11, 2551. https://doi.org/10.3390/ani11092551
Petraccioli A, Crovato P, Guarino FM, Mezzasalma M, Odierna G, Picariello O, Maio N. Chromosome Diversity and Evolution in Helicoide a (Gastropoda: Stylommatophora): A Synthesis from Original and Literature Data. Animals. 2021; 11(9):2551. https://doi.org/10.3390/ani11092551
Chicago/Turabian StylePetraccioli, Agnese, Paolo Crovato, Fabio Maria Guarino, Marcello Mezzasalma, Gaetano Odierna, Orfeo Picariello, and Nicola Maio. 2021. "Chromosome Diversity and Evolution in Helicoide a (Gastropoda: Stylommatophora): A Synthesis from Original and Literature Data" Animals 11, no. 9: 2551. https://doi.org/10.3390/ani11092551
APA StylePetraccioli, A., Crovato, P., Guarino, F. M., Mezzasalma, M., Odierna, G., Picariello, O., & Maio, N. (2021). Chromosome Diversity and Evolution in Helicoide a (Gastropoda: Stylommatophora): A Synthesis from Original and Literature Data. Animals, 11(9), 2551. https://doi.org/10.3390/ani11092551