
Intestinal Microbiota, Anti-Inflammatory, and Anti-Oxidative Status of Broiler Chickens Fed Diets Containing Mushroom Waste Compost By-Products

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Characteristics of Pennisetum Purpureum Schum No.2 (PP), PP Waste Mushroom Compost (PWMC), and Saccharomyces Cerevisiae Fermented PWMC (FPW)
2.1.1. Crude Polysaccharide Measurement
2.1.2. Phenolic Components
2.1.3. Free Radical Scavenging Ability of 2,2-. Diphenyl-1-Picrylhydrazyl (DPPH)
2.1.4. Reducing Powder
2.1.5. Malondialdehyde Production
2.1.6. Ferrous Chelating Capacity
2.2. Animal Experiment Design
2.3. Serum Characteristics
2.4. DNA Extraction and 16S rRNA Gene Sequencing
2.5. Total RNA Isolation, qPCR, and Sequencing
2.6. Statistical Analysis
3. Results
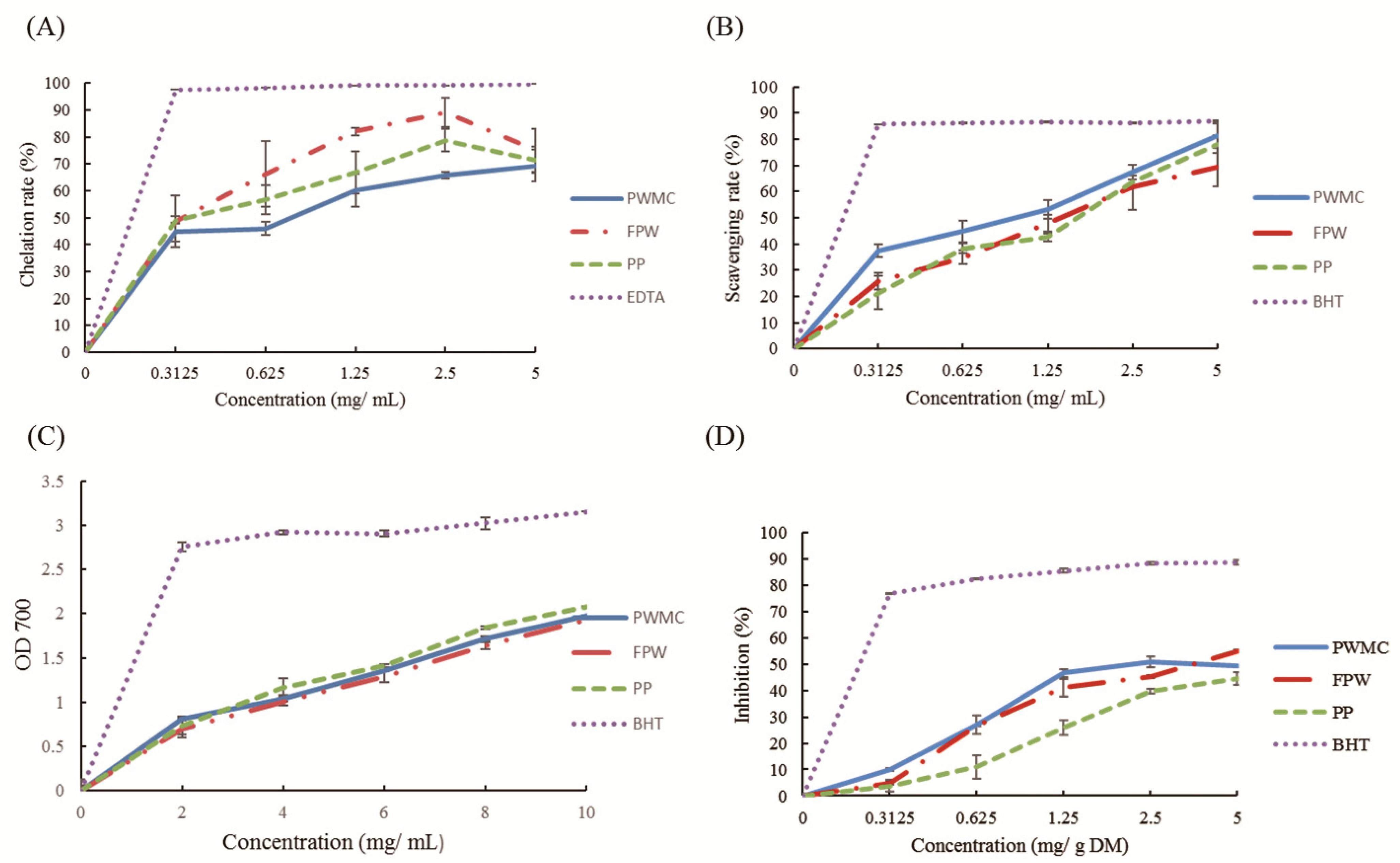
3.1. The Characteristics of PWMC, FPW, and PP
3.2. Serum Characteristic of Broilers
3.3. Gut Microbiota Composition and Function
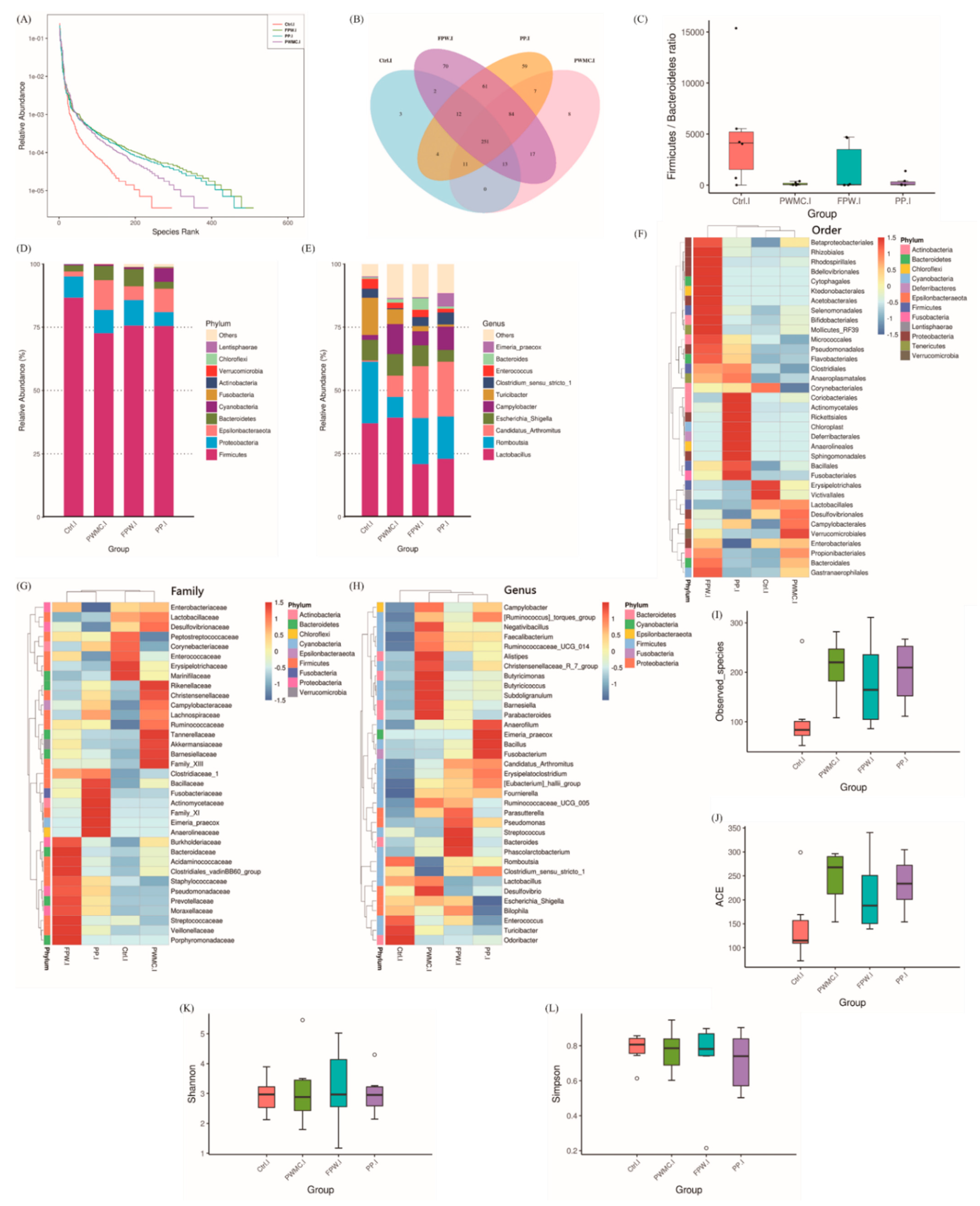
3.3.1. Microbiota Composition in the Ileum of Broilers
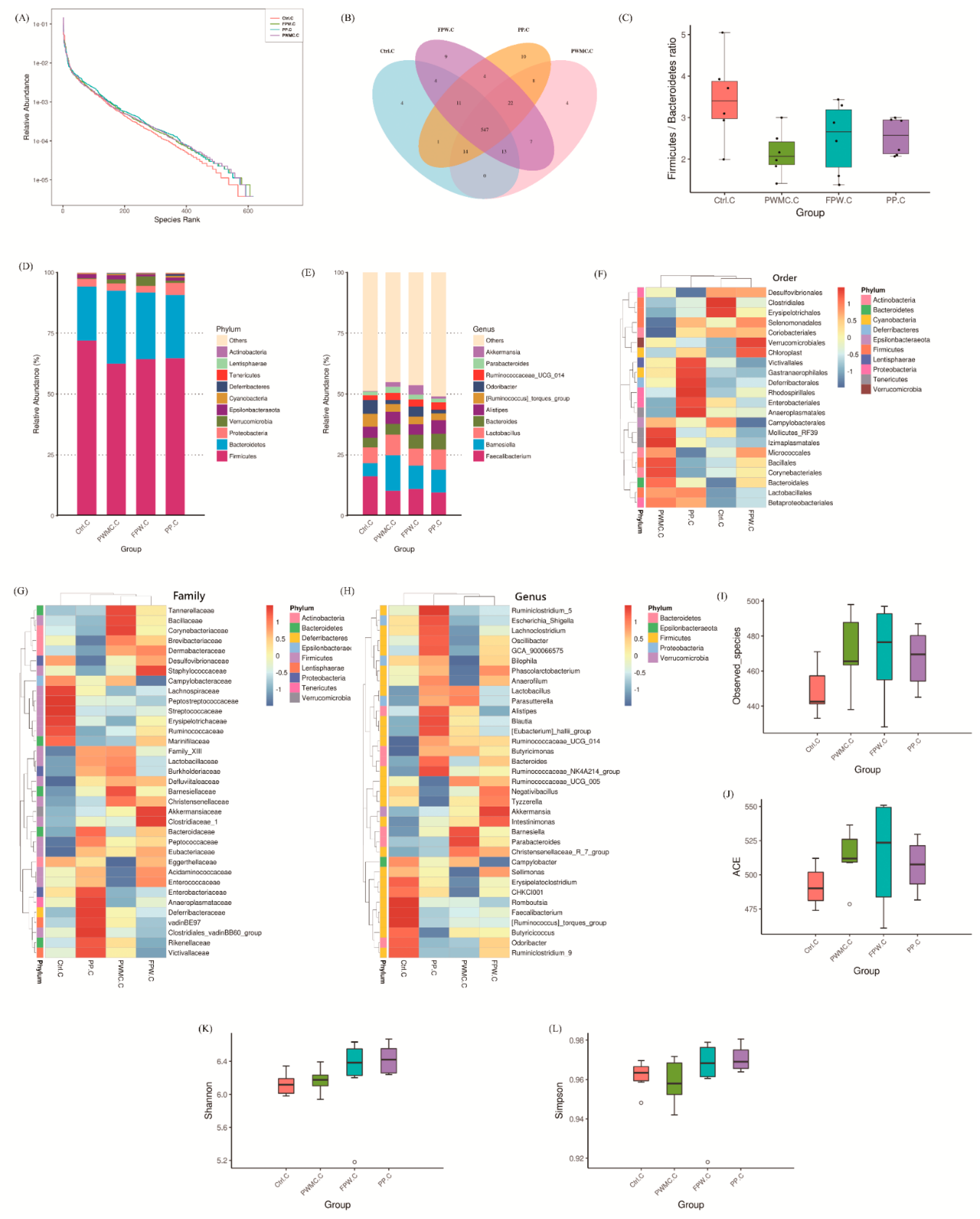
3.3.2. Microbiota Composition in the Cecum of Broilers
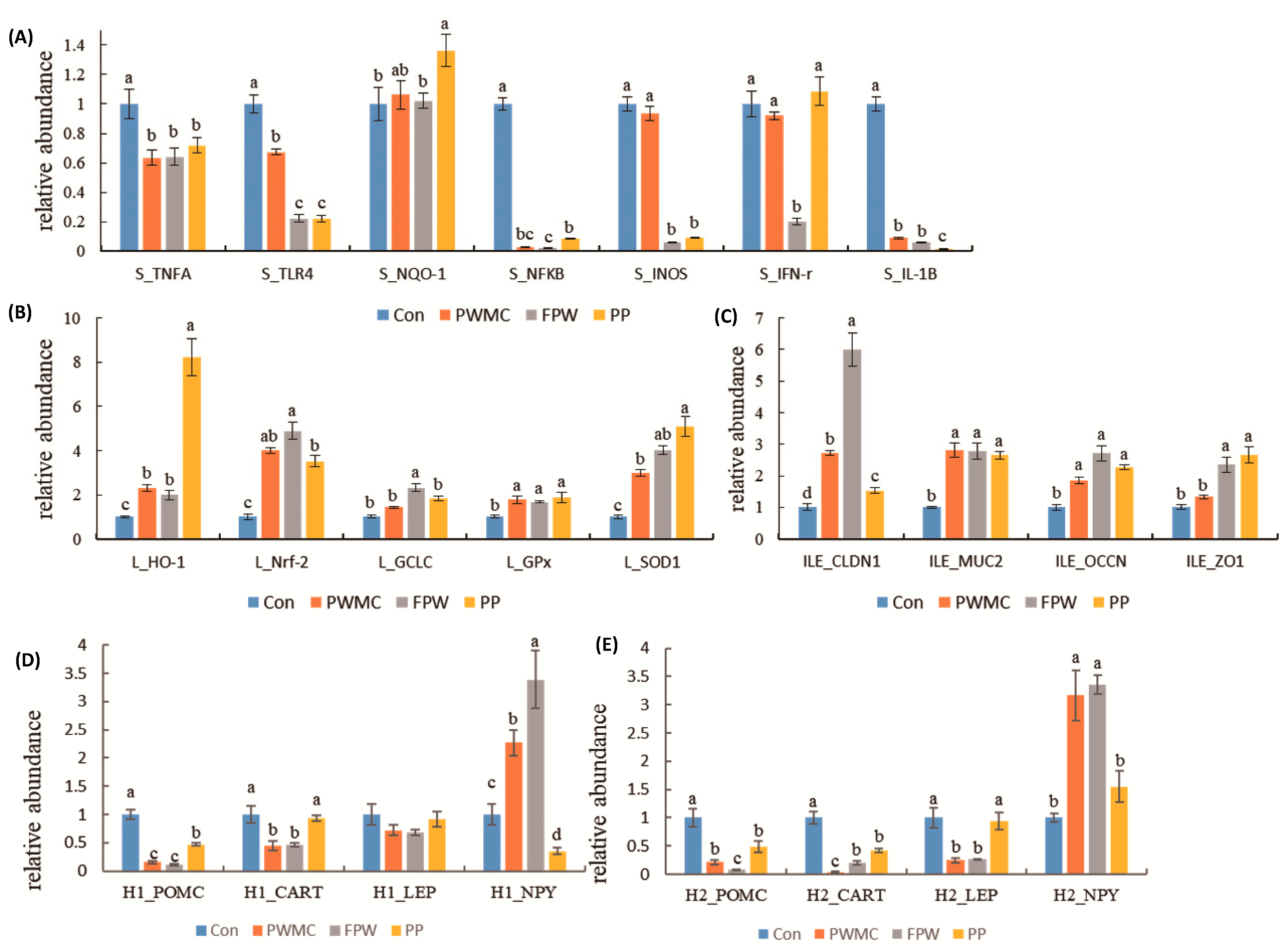
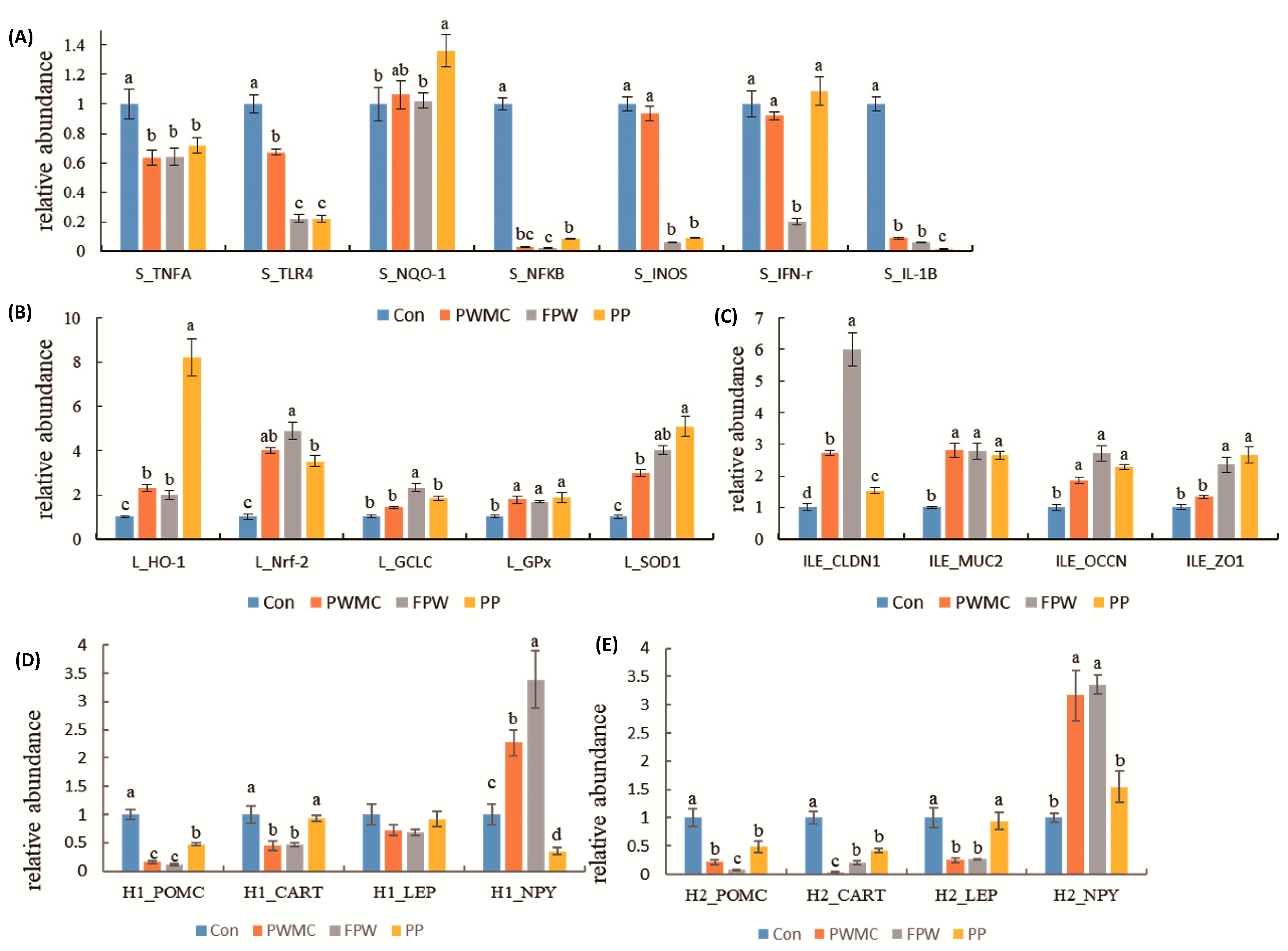
3.4. mRNA Expression in Liver, Spleen, and Ileum of Broilers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE | Abundance based coverage estimator |
| ADF | Acid detergent fiber |
| ARE | Antioxidant responsive element |
| BHT | Butylated hydroxytoluene |
| CC | Catechin |
| CG | Catechin gallate |
| CP | Crude protein |
| DM | Dry matter |
| DPPH | Di (phenyl)-(2,4,6-trinitrophenyl) iminoazanium |
| EC | Epicatechin |
| ECG | Epicatechin gallate |
| EE | Ether extra |
| EGC | Epigallocatechin |
| EGCG | Epigallocatechin gallate |
| FCR | Feed conversion rate |
| FPW | Saccharomyces cerevisiae fermented Pennisetum purpureum schum No.2 waste mushroom compost |
| GA | Gallic acid; GAE: Gallic acid equivalent |
| GC | Gallocatechin |
| GCLC | Glutamate-cysteine ligase catalytic |
| GPx | Glutathione peroxidase |
| GSH | Glutathione |
| HO-1 | Heme oxygenase-1 |
| IFN-γ | Interferon gamma |
| IL-1β | Interleukin 1 beta |
| LEP | Leptin |
| MDA | Malondialdehyde |
| MUC2 | Mucin2 |
| NDF | Neuteral detergent fiber |
| NFκB | Nuclear factor kappa B |
| Nrf2 | Nuclear factor erythroid 2–related factor 2 |
| NPY | Neuropeptide Y |
| OTUs | Operational taxonomic units |
| PP | Pennisetum purpureum schum No.2 |
| POMC | Pro-opiomelanocortin |
| PWMC | Pennisetum purpureum schum No.2 waste mushroom compost |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| TBARS | Thiobarbituric acid reactive substances |
| TNF-α | Tumor necrosis factor alpha |
| ZO-1 | Zonula occludens 1 |
| iNOS | Inducible nitric oxide synthases |
References
- Eshel, G.; Shepon, A.; Makov, T.; Milo, R. Land, irrigation water, greenhouse gas, and reactive nitrogen burdens of meat, eggs, and dairy production in the United States. Proc. Natl. Acad. Sci. USA 2014, 111, 11996–12001. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.P.; Lawrie, A.C.; Huynh, T.T.; Morrison, P.D.; Mautner, A.; Bismarck, A.; John, S. Agricultural by-product suitability for the production of chitinous composites and nanofibers utilising Trametes versicolor and Polyporus brumalis mycelial growth. Process Biochem. 2019, 80, 95–102. [Google Scholar] [CrossRef]
- Teng, P.Y.; Chang, C.L.; Huang, C.M.; Chang, S.C.; Lee, T.T. Effects of solid-state fermented wheat bran by Bacillus amyloliquefaciens and Saccharomyces cerevisiae on growth performance and intestinal microbiota in broiler chickens. Ital. J. Anim. Sci. 2017, 16, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.C.; Lin, L.J.; Chao, Y.P.; Chiang, C.J.; Lee, M.T.; Chang, S.C.; Yu, B.; Lee, T.T. Antioxidant molecular targets of wheat bran fermented by white rot fungi and its potential modulation of antioxidative status in broiler chickens. Br. Poult. Sci. 2017, 58, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.G.; Lin, W.C.; Lo, C.T.; Chang, S.C.; Yu, B.; Lee, T.T. Effects of substitution of Bermuda grass hay with Trichoderma fermented rice straw on growth, blood, and rumen fluid parameters in Barbados sheep. J. Appl. Anim. Res. 2018, 46, 1162–1168. [Google Scholar] [CrossRef] [Green Version]
- Buxton, D.R.; Redfearn, D.D. Plant limitations to fiber digestion and utilization. J. Nutr. 1997, 127, 8145–8185. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Stappenbeck, T.S.; Martens, E.C. A Dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Chassaing, B.; Singh, V.; Pellizzon, M.; Ricci, M.; Fythe, M.D.; Kumar, M.V.; Gewirtz, A.T. Fiber-mediated nourishment of gut microbiota protects against diet-induced obesity by restoring IL-22-mediated colonic health. Cell Host Microbe 2018, 23, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.T.; Lin, W.C.; Lee, T.T. Potential crosstalk of oxidative stress and immune response in poultry through phytochemicals—A review. Asian Aust. J. Anim. Sci. 2019, 32, 309–319. [Google Scholar] [CrossRef]
- Okrathok, S.; Khempaka, S. Modified-dietary fiber from cassava pulp reduces abdominal fat and meat cholesterol contents without affecting growth performance of broiler chickens. J. Appl. Poult. Res. 2020, 29, 229–239. [Google Scholar] [CrossRef]
- Chuang, W.Y.; Lin, W.C.; Hsieh, Y.C.; Huang, C.M.; Chang, S.C.; Lee, T.T. Evaluation of the combined use of saccharomyces cerevisiae and aspergillus oryzae with phytase fermentation products on growth, inflammatory, and intestinal morphology in broilers. Animals 2019, 9, 1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.M.; Lee, T.T. Immunomodulatory effects of phytogenics in chickens and pigs—A review. Asian Aust. J. Anim. Sci. 2018, 31, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Sanchez, S.; D’Souza, D.; Biswas, D.; Hanning, I. Botanical alternatives to antibiotics for use in organic poultry production. Poult. Sci. 2015, 94, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Lee, T.T. Effects of Laetiporus sulphureus-fermented wheat bran on growth performance, intestinal microbiota and digesta characteristics in broiler chickens. Animals 2020, 10, 1457. [Google Scholar] [CrossRef] [PubMed]
- Kermanshahi, H.; Shakouri, M.D.; Daneshmand, A. Effects of non-starch polysaccharides in semi-purified diets on performance, serum metabolites, gastrointestinal morphology, and microbial population of male broiler chickens. Livest. Sci. 2018, 214, 93–97. [Google Scholar] [CrossRef]
- Lee, M.T.; Lin, W.C.; Lee, T.T. Antioxidant capacity of phytochemicals and their potential effects on oxidative status in animals—A review. Asian Aust. J. Anim. Sci. 2017, 30, 299–308. [Google Scholar] [CrossRef]
- Lin, W.C.; Lee, T.T. Laetiporus sulphureus fermented wheat bran enhanced the antioxidant status, intestinal tight junction, and morphology of broiler chickens. Animals 2021, 11, 149. [Google Scholar] [CrossRef]
- Chen, L.W.; Chuang, W.Y.; Hsieh, Y.C.; Lin, H.H.; Lin, W.C.; Lin, L.J.; Chang, S.C.; Lee, T.T. Effects of dietary supplementation with Taiwanese tea byproducts and probiotics on growth performance, lipid metabolism, and the immune response in red feather native chickens. Anim. Biosci. 2021, 34, 393–404. [Google Scholar] [CrossRef]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of diurnal variation of gut microbes and high-fat feeding on host circadian clock function and metabolism. Cell Host Microbe 2015, 17, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Hsu, B.B.; Gibson, T.E.; Yeliseyev, V.; Liu, Q.; Lyon, L.; Bry, L.; Silver, P.A.; Gerber, G.K. Dynamic modulation of the gut microbiota and metabolome by bacteriophages in a mouse model. Cell Host Microbe 2019, 25, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Simpson, H.L.; Campbell, B.J. Review article: Dietary fiber–microbiota interactions. Aliment. Pharmacol. Ther. 2015, 42, 158–179. [Google Scholar] [CrossRef] [Green Version]
- AOAC International. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2012. [Google Scholar]
- Nielsen, S.S. (Ed.) Phenol-sulfuric acid method for total carbohydrates. In Food Analysis Laboratory Manual; Food Science Texts Series; Springer: Boston, MA, USA, 2010. [Google Scholar]
- Tabart, J.; Kevers, C.; Pincemail, J.; Defraigne, J.O.; Dommes, J. Comparative antioxidant capacities of phenolic compounds measured by various tests. Food Chem. 2009, 113, 1226–1233. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on product of browning reaction prepared from glucose amine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Zeb, A.; Ullah, F. A simple spectrophotometric method for the determination of thiobarbituric acid reactive substances in fried fast foods. J. Anal. Methods Chem. 2016, 2016, 9412767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuris, A.; Siow, L.F. A Comparative study of the antioxidant properties of three pineapple (Ananas Comosus L.) varieties. J. Food Stud. 2014, 3, 40–56. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protocols 2001, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoz, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Tech. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Kamboh, A.A.; Zhu, W.Y. Effect of increasing levels of bioflavonoids in broiler feed on plasma anti-oxidative potential, lipid metabolites, and fatty acid composition of meat. Poult. Sci. 2013, 92, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2007, 88, 1243–1276. [Google Scholar] [CrossRef] [Green Version]
- Bognar, E.; Sarszegi, Z.; Szabo, A.; Debreceni, B.; Kalman, N.; Tucsek, Z.; Sumegi, B.; Gallyas, F., Jr. Antioxidant and anti-inflammatory effects in RAW264.7 macrophages of malvidin, a major red wine polyphenol. PLoS ONE 2013, 8, e65355. [Google Scholar] [CrossRef]
- Lau, J.; Herzog, H. CART in the regulation of appetite and energy homeostasis. Front. Neurosci. 2014, 8, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delp, M.S.; Cline, M.A.; Gilbert, E.R. The central effects of alpha-melanocyte stimulating hormone (-MSH) in chicks involve changes in gene expression of neuropeptide Y and other factors in distinct hypothalamic nuclei. Neurosci. Lett. 2017, 651, 52–56. [Google Scholar] [CrossRef]
- Seroussi, E.; Cinnamon, Y.; Yosefi, S.; Genin, O.; Smith, J.G.; Rafati, N.; Bornelov, S.; Andersson, L.; Friedman-Einat, M. Identification of the long-sought leptin in chicken and duck: Expression pattern of the highly GC-Rich avian leptin fitsan autocrine/paracrine rather than endocrine function. Endocrinology 2016, 157, 737–751. [Google Scholar] [CrossRef] [Green Version]
- Geary, N. Control-theory models of body-weight regulation and body-weight regulatory appetite. Appetite 2020, 144, 104440. [Google Scholar] [CrossRef]
- Shang, H.; Zhao, J.; Dong, X.; Guo, Y.; Zhang, H.; Cheng, J.; Zhou, H. Inulin improves the egg production performance and affects the cecum microbiota of laying hens. Int. J. Biol. Macromol. 2020, 155, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Munyaka, P.M.; Rabbi, M.F.; Khafipour, E.; Ghia, J.E. Acute dextran sulfate sodium (DSS)-induced colitis promotes gut microbial dysbiosis in mice. J. Basic Microbiol. 2016, 56, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Froebel, L.K.; Jalukar, S.; Lavergne, T.A.; Lee, J.T.; Duong, T. Administration of dietary prebiotics improves growth performance and reduces pathogen colonization in broiler chickens. Poult. Sci. 2019, 98, 6668–6676. [Google Scholar] [CrossRef]
- Li, H.; Zhang, T.; Li, C.; Zheng, S.; Li, H.; Yu, J. Development of a microencapsulated synbiotic product and its application in yoghurt. Food Sci. Tech. 2020, 122, 109033. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Starter (1–21 Days) | Finisher (22–35 Days) | ||||||
|---|---|---|---|---|---|---|---|---|
| Con | PWMC | FPW | PP | Con | PWMC | FPW | PP | |
| ----------------------------------------------g/kg--------------------------------------------- | ||||||||
| Yellow corn | 519.9 | 452.0 | 457.5 | 456.4 | 571.6 | 535.9 | 537.3 | 537.1 |
| Soybean meal (CP 44.0%) | 369.4 | 307.1 | 302.5 | 304.9 | 246.9 | 236.6 | 234.2 | 236.1 |
| Full fat soybean meal | 0.00 | 80.00 | 80.00 | 80.00 | 50.00 | 48.90 | 49.00 | 48.99 |
| Fish meal (CP 65.0%) | 30.00 | 30.00 | 30.00 | 30.00 | 50.00 | 50.00 | 50.00 | 50.00 |
| Soybean oil | 40.58 | 40.83 | 39.86 | 38.54 | 41.39 | 38.49 | 39.33 | 37.70 |
| Calcium carbonate | 16.50 | 16.50 | 16.50 | 16.50 | 16.50 | 16.50 | 16.50 | 16.50 |
| Monocalcium phosphate | 11.20 | 11.20 | 11.20 | 11.20 | 11.20 | 11.20 | 11.20 | 11.20 |
| NaCl | 3.9 | 3.9 | 3.9 | 3.9 | 3.9 | 3.9 | 3.9 | 3.9 |
| L-Lysine-HCl | 3.7 | 3.7 | 3.7 | 3.7 | 3.7 | 3.7 | 3.7 | 3.7 |
| DL-Methionine | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 |
| Choline-Cl | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
| Vitamin premix 2 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Mineral premix 3 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| PWMC | 0 | 50 | 0 | 0 | 0 | 50 | 0 | 0 |
| FPW | 0 | 0 | 50 | 0 | 0 | 0 | 50 | 0 |
| PP | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 50 |
| Total | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 |
| Calculated nutrient value | ||||||||
| Dry matter,% | 88 | 89 | 89 | 88 | 88 | 91 | 90 | 90 |
| Crude protein,%DM | 23.0 | 23.0 | 23.0 | 23.0 | 21.0 | 20.9 | 21.0 | 21.0 |
| Calcium,%DM | 1.11 | 1.12 | 1.11 | 1.13 | 1.21 | 1.22 | 1.22 | 1.23 |
| Total Phosphorus,%DM | 0.68 | 0.72 | 0.72 | 0.69 | 0.72 | 0.76 | 0.76 | 0.72 |
| Available Phosphorus,%DM | 0.46 | 0.48 | 0.48 | 0.46 | 0.52 | 0.54 | 0.54 | 0.51 |
| Methionine + Cystein,%DM | 0.92 | 0.93 | 0.93 | 0.90 | 0.89 | 0.90 | 0.90 | 0.87 |
| ME, kcal/kg DM | 3050 | 3050 | 3050 | 3050 | 3175 | 3175 | 3175 | 3175 |
| Chemical analysis value | ||||||||
| Dry matter,% | 88.7 | 89.2 | 89.8 | 88.5 | 89.3 | 89.1 | 90.3 | 90.2 |
| Crude protein,%DM | 23.1 | 23.1 | 23.0 | 23.0 | 21.0 | 21.0 | 21.0 | 21.0 |
| Crude fat,%DM Crude fiber,%DM | 6.85 | 7.94 | 7.86 | 7.88 | 7.96 | 7.66 | 7.73 | 7.72 |
| 3.60 | 6.42 | 6.14 | 6.78 | 3.11 | 6.03 | 5.74 | 6.39 | |
| Gene Name 1 | Primer Sequence | Accession No. | Size (bp) |
|---|---|---|---|
| ß-actin | F: 5′-CTGGCACCTAGCACAATGAA-3′ R: 5′-ACATCTGCTGGAAGGTGGAC-3′ | X00182.1 | 109 |
| TNF | F: 5′-TGTGTATGTGCAGCAACCCGTAGT-3′ R: 5′-GGCATTGCAATTTGGACAGAAGT-3′ | NM 204267 | 229 |
| TLR4 | F: 5′-TGCACAGGACAGAACATCTCTGGA-3′ R: 5′-AGCTCCTGCAGGGTATTCAAGTGT-3′ | NM_001030693 | 347 |
| NQO-1 | F: 5′-AAGAAGATTGAAGCGGCTGA-3′ R: 5′-GCATGGCTTTCTTCTTCTGG-3′ | NM_001277619.1 | 166 |
| NFκB | F: 5′- CCAGGTTGCCATCGTGTTCC-3′ R: 5′- GCGTGCGTTTGCGCTTCT-3′ | D13719.1 | 179 |
| iNOS | F: 5′-TACTGCGTGTCCTTTCAACG-3′ R: 5′-CCCATTCTTCTTCCAACCTC-3′ | U46504 | 108 |
| IFN-γ | F: 5′-CTCCCGATGAACGACTTGAG-3′ R: 5′-CTGAGACTGGCTCCTTTTCC-3′ | Y07922 | 111 |
| IL-1ß | F: 5′-GCTCTACATGTCGTGTGTGATGAG-3′ R: 5′-TGTCGATGTCCCGCATGA-3′ | NM_204524 | 80 |
| HO-1 | F: 5′-GGTCCCGAATGAATGCCCTTG-3′ R: 5′-ACCGTTCTCCTGGCTCTTGG-3′ | NM_205344.1 | 137 |
| Nrf-2 | F: 5′-GGAAGAAGGTGCGTTTCGGAGC-3′ R: 5′-GGGCAAGGCAGATCTCTTCCAA-3′ | NM_205117.1 | 171 |
| GCLC | F: 5′-CAGCACCCAGACTACAAGCA-3′ R: 5′-CTACCCCCAACAGTTCTGGA-3′ | XM_419910.3 | 118 |
| GPX7 | F: 5′- CAGCAAGAACCAGACACCAA-3′ R: 5′- CCAGGTTGGTTCTTCTCCAG-3′ | NM_001163245.1 | 156 |
| SOD-1 | F: 5′- ATTACCGGCTTGTCTGATGG-3′ R: 5′- CCTCCCTTTGCAGTCACATT-3′ | NM_205064.155 | 173 |
| Claudin-1 | F: 5′-GGAGGATGACCAGGTGAAGA-3′ R: 5′-TCTGGTGTTAACGGGTGTGA-3′ | NM_001013611.2 | 149 |
| MUC-2 | F: 5′-GCTACAGGATCTGCCTTTGC-3′ R: 5′-AATGGGCCCTCTGAGTTTTT-3′ | JX284122.1 | 152 |
| Occludin | F: 5′-GTCTGTGGGTTCCTCATCGT-3′ R: 5′-GTTCTTCACCCACTCCTCCA-3′ | NM_205128.1 | 178 |
| ZO-1 | F: 5′-AGGTGAAGTGTTTCGGGTTG-3′ R: 5′-CCTCCTGCTGTCTTTGGAAG-3′ | XM_015278975.1 | 145 |
| POMC | 5′-AAGGCGAGGAGGAAAAGAAG-3′ 5′-CCTTCTTGTAGGCGCTTTTG-3′ | XM_015285103.2 | 168 |
| CART | 5′- CCTCGTGCAACTCCTTTCTT-3′ 5′- TTTCCTGAACGGACGAAAAC-3′ | XM_003643097.4 | 173 |
| LEP | 5′-AGCAACGATTGAGGCGATT-3′ 5′-AGCAGCTCCTTCAACTCAGG-3′ | LN794246.1 | 208 |
| NPY | 5′-CCTCATCACCAGGCAGAGAT-3′ 5′-CACTGGGAATGACGCTATGA-3′ | NM_205473.1 | 212 |
| Items 1 | PWMC | FPW | PP |
|---|---|---|---|
| Functional component analysis | |||
| Crude polysaccharide (Glucose mg/g DM) | 85.0 ± 1.23 | 83.0 ± 1.10 | 87.9 ± 2.1 |
| Total VFA (μmole/g) | 30.8 ± 1.4 | 28.1 ± 0.6 | 12.2 ± 0.3 |
| Phenol-like chemical analysis (μg/g) | |||
| Gallic acid | 114 ± 2.7 | 159 ± 2.7 | 127 ± 2.4 |
| Gallocatechin | 1035 ± 8.0 | 1404 ± 23.0 | 1245 ± 15 |
| Epigallocatechin | 1493 ± 14.0 | 1817 ± 23.0 | 1687 ± 17 |
| Catechin | 7.90 ± 0.6 | 169 ± 0.9 | 13 ± 0.65 |
| Caffeic acid | 113 ± 0.5 | 235 ± 1.5 | 127 ± 1.2 |
| Epicatechin | 230 ± 2.0 | 213 ± 9.7 | 231 ± 5.8 |
| Epigallocatechin gallate | 210 ± 0.1 | 216 ± 3.5 | 202 ± 2.2 |
| Epicatechin gallate | 78.1 ± 2.8 | 199 ± 1.9 | 102 ± 2.2 |
| Catechin gallate | 401 ± 5.1 | 534 ± 7.3 | 430 ± 6.2 |
| Chemical analysis 2 | |||
| DM | 94.6 ± 0.4 | 94.2 ± 0.5 | 94.6 ± 0.6 |
| CP (% DM) | 6.8 ± 0.35 | 10.0 ± 0.53 | 12.6 ± 0.43 |
| EE (% DM) | N.D 3 | N.D | N.D |
| Ash (% DM) | 10.5 ± 0.4 | 15.5 ± 0.7 | 12.4 ± 0.8 |
| NDF (% DM) | 59.7 ± 0.8 | 54.4 ± 1.6 | 55.2 ± 0.9 |
| ADF (% DM) | 48.8 ± 2.8 | 34.3 ± 0.5 | 38.2 ± 1.7 |
| Items | Treatments | SEM 2 | p Value | |||
|---|---|---|---|---|---|---|
| Con | PWMC | FPW | PP | |||
| GLU (mg/dL) | 276 a | 236 b | 240 b | 227 b | 5.2 | <0.0001 |
| Protein | ||||||
| SGOT (U/L) | 332 | 251 | 248 | 246 | 24 | 0.0555 |
| SGPT (U/L) | 4.17 | 3.67 | 4.33 | 5.33 | 0.72 | 0.4432 |
| T-P (g/dL) | 3.4 | 3.18 | 3.1 | 3.13 | 0.08 | 0.0534 |
| ALB (g/dL) | 1.55 a | 1.47 ab | 1.4 b | 1.45 ab | 0.03 | 0.0432 |
| GLO (g/dL) | 1.85 | 1.72 | 1.7 | 1.68 | 0.06 | 0.1662 |
| Alk-P (IU/L) | 1086 | 1280 | 1366 | 1357 | 234 | 0.8185 |
| Gpx (nmol/min/mL) | 114.7 | 143.3 | 124.2 | 219.8 | 41.9 | 0.4080 |
| SOD (mU/mL) | 523.9 b | 539.2 b | 672.1 a | 620.7 a | 23.42 | 0.0009 |
| TNF-α (pg/mL) | 284.7 a | 145.4 b | 155.9 b | 195.8 b | 19.56 | 0.0019 |
| Lipid | ||||||
| CHOL (mg/dL) | 125 | 121 | 112 | 120 | 4.1 | 0.1542 |
| TG (mg/dL) | 54.8 a | 40.5 b | 38.2 b | 40.2 b | 2.7 | 0.0008 |
| HDL-C (mg/dL) | 78 | 74.7 | 69.3 | 72.2 | 2.7 | 0.1562 |
| LDL-C (mg/dL) | 42.2 | 39.5 | 39.7 | 44.2 | 2.0 | 0.3415 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, W.Y.; Lin, L.J.; Shih, H.D.; Shy, Y.M.; Chang, S.C.; Lee, T.T. Intestinal Microbiota, Anti-Inflammatory, and Anti-Oxidative Status of Broiler Chickens Fed Diets Containing Mushroom Waste Compost By-Products. Animals 2021, 11, 2550. https://doi.org/10.3390/ani11092550
Chuang WY, Lin LJ, Shih HD, Shy YM, Chang SC, Lee TT. Intestinal Microbiota, Anti-Inflammatory, and Anti-Oxidative Status of Broiler Chickens Fed Diets Containing Mushroom Waste Compost By-Products. Animals. 2021; 11(9):2550. https://doi.org/10.3390/ani11092550
Chicago/Turabian StyleChuang, Wen Yang, Li Jen Lin, Hsin Der Shih, Yih Min Shy, Shang Chang Chang, and Tzu Tai Lee. 2021. "Intestinal Microbiota, Anti-Inflammatory, and Anti-Oxidative Status of Broiler Chickens Fed Diets Containing Mushroom Waste Compost By-Products" Animals 11, no. 9: 2550. https://doi.org/10.3390/ani11092550
APA StyleChuang, W. Y., Lin, L. J., Shih, H. D., Shy, Y. M., Chang, S. C., & Lee, T. T. (2021). Intestinal Microbiota, Anti-Inflammatory, and Anti-Oxidative Status of Broiler Chickens Fed Diets Containing Mushroom Waste Compost By-Products. Animals, 11(9), 2550. https://doi.org/10.3390/ani11092550