
Seasonal and Nutritional Fluctuations in the mRNA Levels of the Short Form of the Leptin Receptor (LRa) in the Hypothalamus and Anterior Pituitary in Resistin-Treated Sheep


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Animal Treatments
2.3.1. Experiment 1. The Effects of Photoperiod and RbresistinTreatment on the Expression of LRa in Selected Brain Areas and Hormone Concentrations
2.3.2. Experiment 2. The Effect of BW and Rbresistin Treatment on the Expression of LRa in Selected Brain Areas and Hormone Concentrations
2.4. Hormone Assays
2.5. Statistics
2.5.1. Statistical Analysis
2.5.2. Molecular Analysis
2.5.3. Molecular Data Analysis
3. Results
3.1. The Effects of Photoperiod and Rbresistin Treatment on the Expression of LRa in Selected Brain Areas and Hormone Concentrations
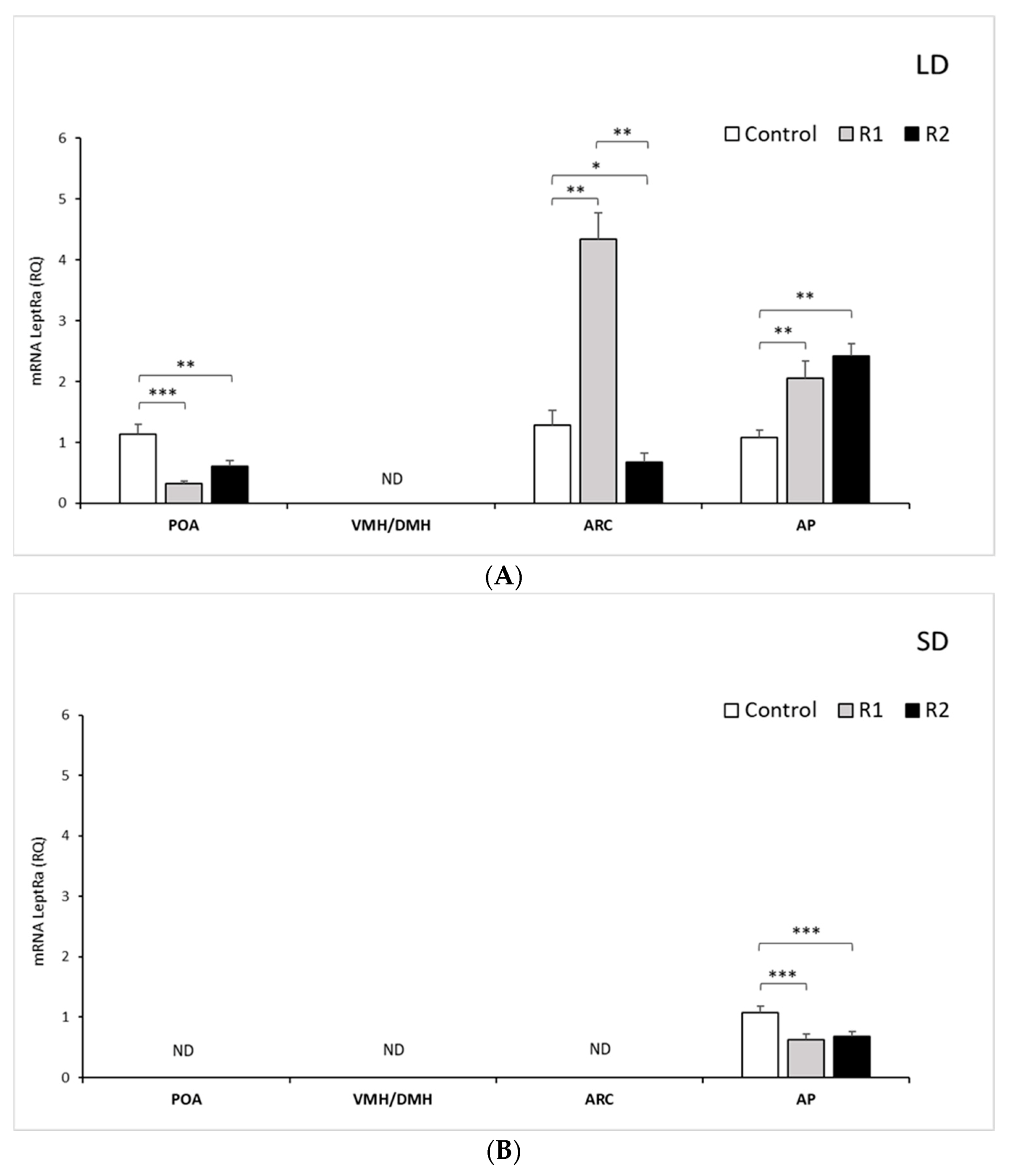
3.1.1. Expression of mRNA LRa in Selected Brain Areas
3.1.2. Estradiol Concentration
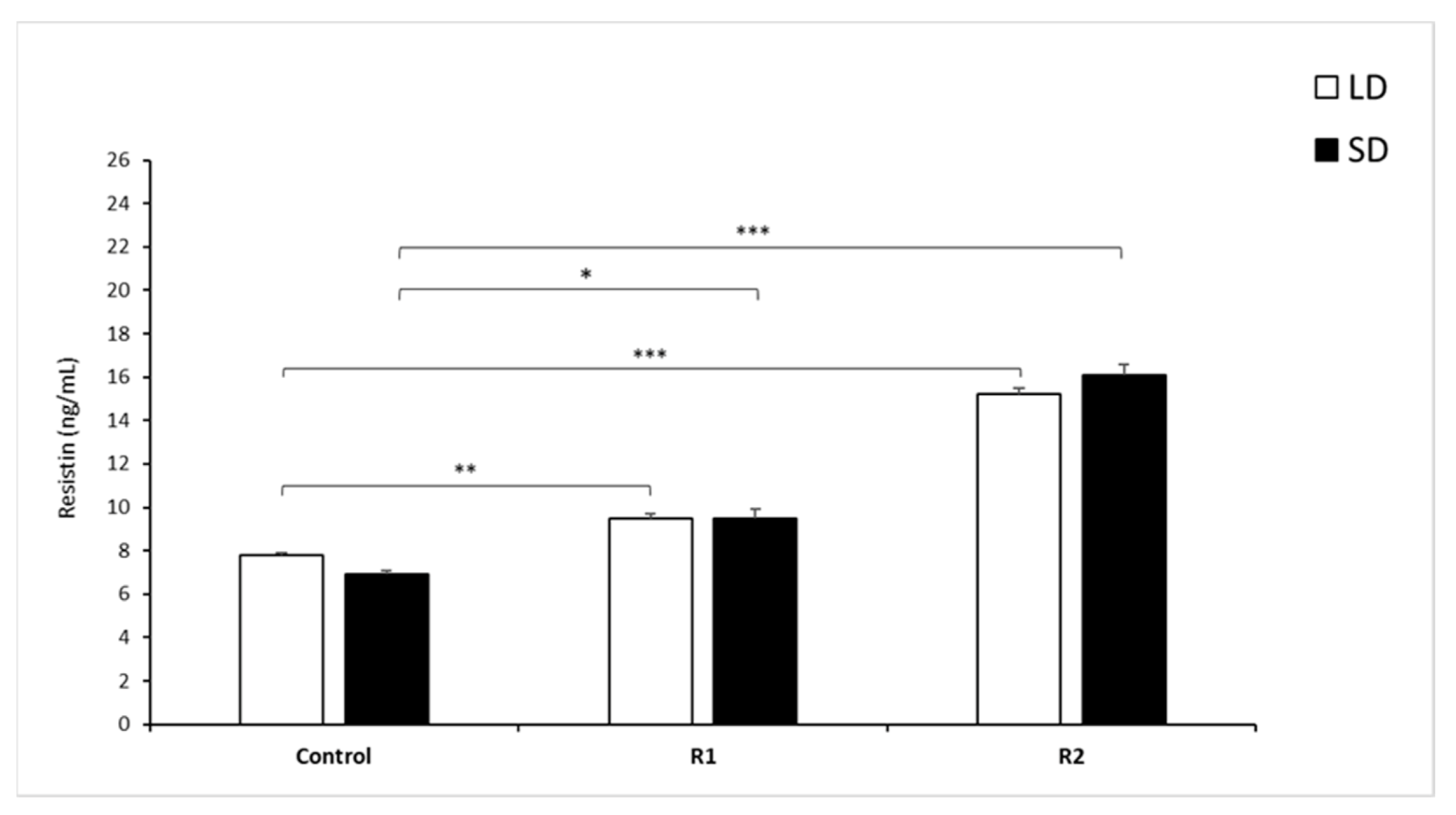
3.1.3. Resistin Concentrations
3.1.4. Leptin Concentrations
3.2. The Effect of BW and Rbresistin Treatment on the Expression of LRa in Selected Brain Areas and Hormone Concentrations
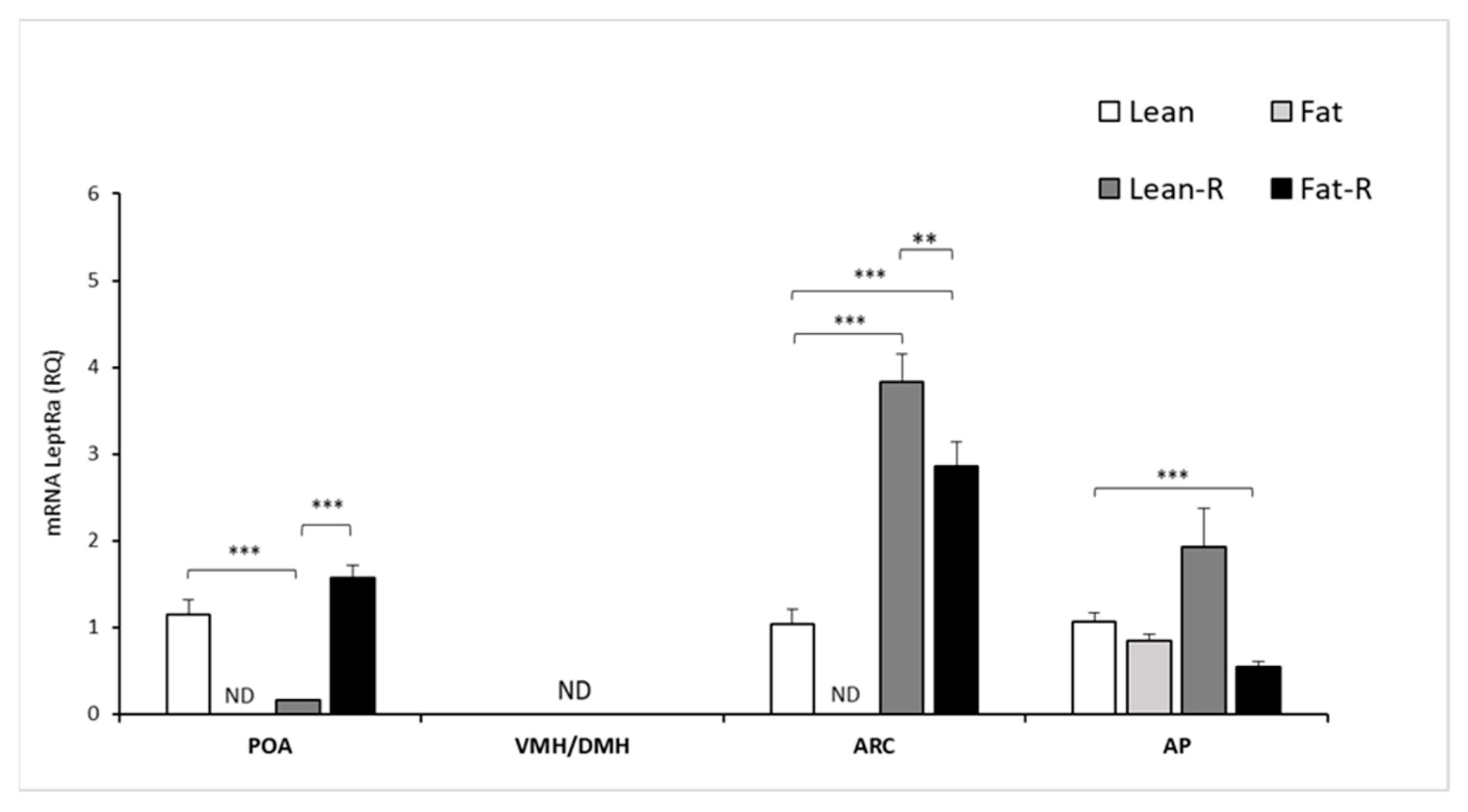
3.2.1. Expression of mRNA LRa in Selected Brain Areas
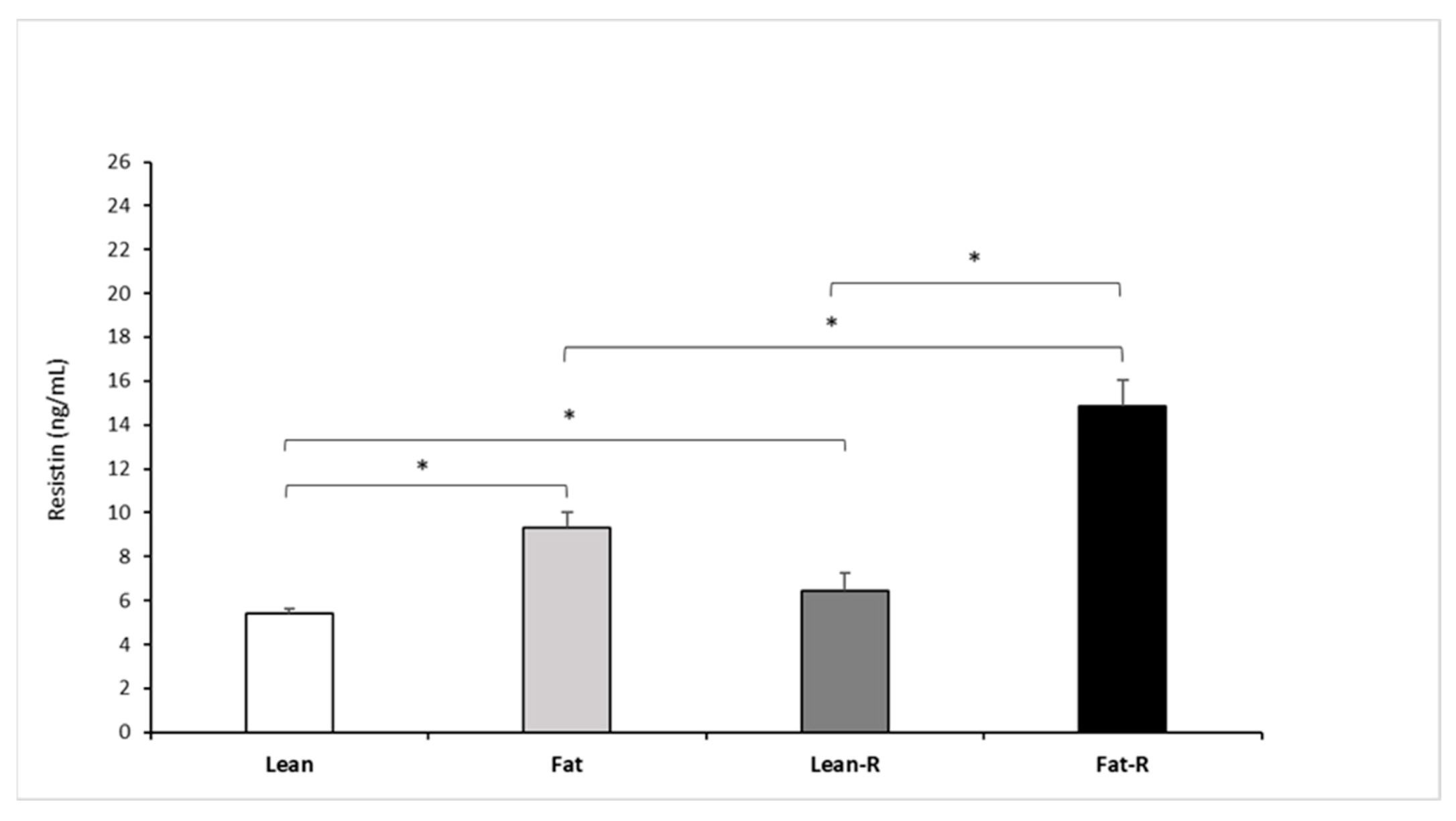
3.2.2. Resistin Concentrations
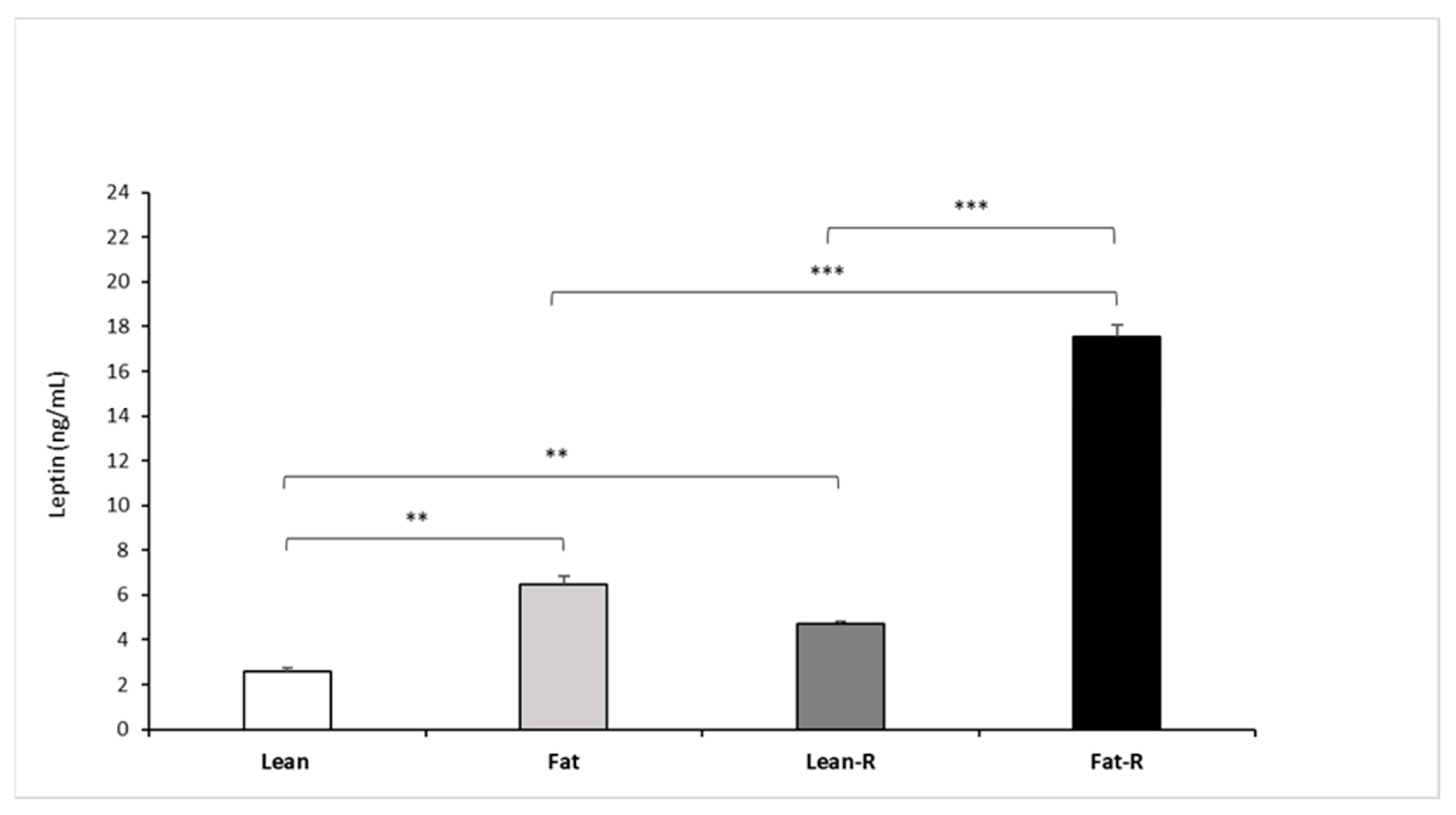
3.2.3. Leptin Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wuliji, T.; Litherland, A.; Goetsch, A.L.; Sahlu, T.; Puchala, R.; Dawson, L.J.; Gipson, T. Evaluation of melatonin and bromocryptine administration in Spanish goats. Small Rumin. Res. 2003, 49, 31–40. [Google Scholar] [CrossRef]
- Spicer, L.J.; Schreiber, N.B.; Lagaly, D.V.; Aad, P.Y.; Douthit, L.B.; Grado-Ahuir, J.A. Effect of resistin on granulosa and theca cell function in cattle. Anim. Reprod. Sci. 2011, 124, 19–27. [Google Scholar] [CrossRef]
- Sarmento-Cabral, A.; Peinado, J.R.; Halliday, L.C.; Malagon, M.M.; Castaño, J.P.; Kineman, R.D.; Luque, R.M. Adipokines (leptin, adiponectin, resistin) differentially regulate all hormonal cell types in primary anterior pituitary cell cultures from two primate species. Sci. Rep. 2017, 7, 43537. [Google Scholar] [CrossRef]
- Biernat, W.; Kirsz, K.; Szczesna, M.; Zieba, D.A. Resistin regulates reproductive hormone secretion from the ovine adenohypophysis depending on season. Domest. Anim. Endocrinol. 2018, 65, 95–100. [Google Scholar] [CrossRef]
- Zieba, D.A.; Biernat, W.; Szczesna, M.; Kirsz, K.; Barć, J.; Misztal, T. Changes in expression of the genes for the leptin signaling in hypothalamic-pituitary selected areas and endocrine responses to long-term manipulation in body weight and resistin in ewes. Int. J. Mol. Sci. 2020, 21, 4238. [Google Scholar] [CrossRef]
- Zieba, D.; Bilińska, B.; Schmalz-Fraczek, B.; Murawski, M. Immunohistochemical localization of estrogen receptors in the ovine corpus luteum throughout the estrous cycle. Folia Histochem. Cytobiol. 2000, 38, 111–117. [Google Scholar]
- Gao, Q.; Wolfgang, M.J.; Neschen, S.; Morino, K.; Horvath, T.L.; Shulman, G.I.; Fu, X.Y. Disruption of neural signal transducer and activator of transcription 3 causes obesity, diabetes, infertility, and thermal dysregulation. Proc. Natl. Acad. Sci. USA 2004, 101, 4661–4666. [Google Scholar] [CrossRef]
- Dalamaga, M.; Chou, S.H.; Shields, K.; Papageorgiou, P.; Polyzos, S.A.; Mantzoros, C.S. Leptin at the intersection of neuroendocrinology and metabolism: Current evidence and therapeutic perspectives. Cell Metab. 2013, 18, 29–42. [Google Scholar] [CrossRef]
- Fields, H.L.; Hjelmstad, G.O.; Margolis, E.B.; Nicola, S.M. Ventral tegmental area neurons in learned appetitive behavior and positive reinforcement. Annu. Rev. Neurosci. 2007, 30, 289–316. [Google Scholar] [CrossRef]
- Zhou, Y.; Rui, L. Leptin signaling and leptin resistance. Front. Med. 2013, 7, 207–222. [Google Scholar] [CrossRef]
- Liu, J.; Yang, X.; Yu, S.; Zheng, R. The leptin signaling. Adv. Exp. Med. Biol. 2018, 1090, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Król, E.; Tups, A.; Archer, Z.A.; Ross, A.W.; Moar, K.M.; Bell, L.M.; Duncan, J.S.; Mayer, C.; Morgan, P.J.; Mercer, J.G.; et al. Altered expression of SOCS3 in the hypothalamic arcuate nucleus during seasonal body mass changes in the field vole, microtusagrestis. J. Neuroendocrinol. 2007, 19, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Ladyman, S.R.; Grattan, D.R. Suppression of leptin receptor messenger ribonucleic acid and leptin responsiveness in the ventromedial nucleus of the hypothalamus during pregnancy in the rat. Endocrinology 2005, 146, 3868–3874. [Google Scholar] [CrossRef] [PubMed]
- Klocek-Gorka, B.; Szczesna, M.; Molik, E.; Zieba, D.A. The interactions of season, leptin and melatonin levels with thyroid hormone secretion, using an in vitro approach. Small Rumin. Res. 2010, 91, 231–235. [Google Scholar] [CrossRef]
- Gogga, P.; Karbowska, J.; Meissner, W.; Kochan, Z. Physiological leptin resistance. Med. Weter. 2016, 72, 604–610. [Google Scholar] [CrossRef][Green Version]
- Gao, Q.; Horvath, T.L. Neurobiology of feeding and energy expenditure. Annu. Rev. Neurosci. 2007, 30, 367–398. [Google Scholar] [CrossRef]
- Williams, K.W.; Elmquist, J.K. From neuroanatomy to behavior: Central integration of peripheral signals regulating feeding behavior. Nat. Neurosci. 2012, 15, 1350–1355. [Google Scholar] [CrossRef]
- Zieba, D.A.; Biernat, W.; Szczesna, M.; Kirsz, K.; Misztal, T. Hypothalamic–pituitary and adipose tissue responses to the effect of resistin in sheep: The integration of leptin and resistin signaling involving a suppressor of cytokine signaling 3 and the long form of the leptin receptor. Nutrients 2019, 11, 2180. [Google Scholar] [CrossRef]
- Russel, A.J.F.; Doney, J.M.; Gunn, R.G. Subjective assessment of body fat in live sheep. J. Agric. Sci. 1969, 72, 451–454. [Google Scholar] [CrossRef]
- Zieba, D.A.; Szczesna, M.; Klocek-Gorka, B.; Molik, E.; Misztal, T.; Williams, G.L.; Romanowicz, K.; Stepien, E.; Keisler, D.H.; Murawski, M. Seasonal effects of central leptin infusion on secretion of melatonin and prolactin and on SOCS-3 gene expression in ewes. J. Endocrinol. 2008, 198, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Norms. Nutrient Requirements for Cattle and Sheep in the Traditional System; IZ Krakow: Krakow, Poland, 1993. (In Polish) [Google Scholar]
- Instytut National de la Recherche Agronomique (INRA). Alimentation des Bovines, Ovins et Caprins. Besoins des Animaux. Valeur des Aliments; INRA: Vesaukkes, France, 2007. [Google Scholar]
- Tartaglia, L.A.; Dembski, M.; Weng, X.; Deng, N.; Culpepper, J.; Devos, R.; Richards, G.J.; Campfield, L.A.; Clark, F.T.; Deeds, J.; et al. Identification and expression cloning of a leptin receptor, OB-R. Cell 1995, 83, 1263–1271. [Google Scholar] [CrossRef]
- Kastin, A.J.; Akerstrom, V. Fasting, but not adrenalectomy, reduces transport of leptin into the brain. Peptides 2000, 21, 679–682. [Google Scholar] [CrossRef]
- Hileman, S.M.; Tornøe, J.; Flier, J.S.; Bjørbæk, C. Transcellular transport of leptin by the short leptin receptor isoform ObRa in Madin-Darby canine Sidney cells. Endocrinology 2000, 141, 1955–1961. [Google Scholar] [CrossRef]
- Hileman, S.M.; Pierroz, D.D.; Masuzaki, H.; Bjørbæk, C.; El-Haschimi, K.; Banks, W.A.; Flier, J.S. Characterization of short isoforms of the leptin receptor in rat cerebral microvessels and of brain uptake of leptin in mouse models of obesity. Endocrinology 2002, 143, 775–783. [Google Scholar] [CrossRef]
- Adam, C.L.; Mercer, J.G. Appetite regulation and seasonality: Implications for obesity. Proc. Nutr. Soc. 2004, 63, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Halaas, J.L.; Boozer, C.; Blair-West, J.; Fidahusein, N.; Denton, D.A.; Friedman, J.M. Physiological response to long-term peripheral and central leptin infusion in lean and obese mice. Proc. Natl. Acad. Sci. USA 1997, 94, 8878–8883. [Google Scholar] [CrossRef] [PubMed]
- Van Heek, M.; Compton, D.S.; France, C.F.; Tedesco, R.P.; Fawzi, A.B.; Graziano, M.P.; Sybertz, E.J.; Strader, C.D.; Davis, H.R., Jr. Diet-induced obese mice develop peripheral, but not central, resistance to leptin. J. Clin. Investig. 1997, 99, 385–390. [Google Scholar] [CrossRef]
- Levin, B.E.; Dunn-Meynell, A.A.; Banks, W.A. Obesity-prone rats have normal blood-brain barrier transport but defective central leptin signaling before obesity onset. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2004, 286, R143–R150. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; DiPalma, C.R.; Farrell, C.L. Impaired transport of leptin across the blood-brain barrier in obesity. Peptides 1999, 20, 1341–1345. [Google Scholar] [CrossRef]
- Kastin, A.J.; Pan, W.; Maness, L.M.; Koletsky, R.J.; Ernsberger, P. Decreased transport of leptin across the blood–brain barrier in rats lacking the short form of the leptin receptor. Peptides 1999, 20, 1449–1453. [Google Scholar] [CrossRef]
- Morton, G.J.; Cummings, D.E.; Baskin, D.G.; Barsh, G.S.; Schwartz, M.W. Central nervous system control of food intake and body weight. Nature 2006, 443, 289–295. [Google Scholar] [CrossRef]
- Marie, M.; Findlay, P.A.; Thomas, L.; Adam, C.L. Daily patterns of plasma leptin in sheep: Effects of photoperiod and food intake. J. Endocrinol. 2001, 170, 277–286. [Google Scholar] [CrossRef]
- Szczesna, M.; Zieba, D.A.; Klocek-Gorka, B.; Keisler, D.H. Interactive in vitro effect of prolactin, growth hormone and season on leptin secretion by ovine adipose tissue: An in vitro study. Small Rumin. Res. 2011, 100, 177–183. [Google Scholar] [CrossRef]
- Adam, C.L.; Findlay, P.A.; Miller, D.W. Blood-brain leptin transport and appetite and reproductive neuroendocrine responses to intracerebroventricular leptin injection in sheep: Influence of photoperiod. Endocrinology 2006, 147, 4589–4598. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Banks, W.A.; Clever, C.M.; Farrell, C.L. Partial saturation and regional variation in the blood-to-brain transport of leptin in normal weight mice. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E1158–E1165. [Google Scholar] [CrossRef] [PubMed]
- Kirsz, K.; Szczesna, M.; Dudek, K.; Bartlewski, P.M.; Zieba, D.A. Influence of season and nutritional status on the direct effects of leptin, orexin-A and ghrelin on luteinizing hormone and growth hormone secretion in the ovine pituitary explant model. Domest. Anim. Endocrinol. 2014, 48, 69–76. [Google Scholar] [CrossRef]
- Asterholm, I.W.; Rutkowski, J.M.; Fujikawa, T.; Cho, Y.R.; Fukuda, M.; Tao, C.; Wang, Z.V.; Gupta, R.K.; Elmquist, J.K.; Scherer, P.E. Elevated resistin levels induce central leptin resistance and increased atherosclerotic progression in mice. Diabetologia 2014, 57, 1209–1218. [Google Scholar] [CrossRef]
- Kastin, A.J.; Hahn, K.; Zadina, J.E. Regional differences in peptide degradation by rat cerebral microvessels. Life Sci. 2001, 69, 1305–1312. [Google Scholar] [CrossRef]
- Zieba, D.A.; Kirsz, K.; Molik, E.; Romanowicz, K.; Wojtowicz, A.K. Effects of orexigenic peptides and leptin on melatonin secretion during different photoperiods in seasonal breeding ewes. An in vitro study. Domest. Anim. Endocrinol. 2011, 40, 139–146. [Google Scholar] [CrossRef]
- Szczesna, M.; Zieba, D.A. Phenomenon of leptin resistance in seasonal animals: The failure of leptin action in the brain. Domest. Anim. Endocrinol. 2015, 52, 60–70. [Google Scholar] [CrossRef]
- Honkisz, E.; Zieba-Przybylska, D.A.; Wojtowicz, A.K. The effect of triclosan on hormone secretion and viability of human choriocarcinoma JEG-3 cells. Reprod. Toxicol. 2012, 34, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Szóstek, A.Z.; Siemieniuch, M.J.; Galvão, A.M.; Lukasik, K.; Zieba, D.; Ferreira-Dias, G.M.; Skarzynski, D.J. Effects of cell storage and passage on basal andoxytocin-regulated prostaglandin secretion by equine endometrial epithelial and stromal cells. Theriogenology 2012, 77, 1698–1708. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V.; Jovanovic, S.; Miao, W.; Samara, S.; Verma, S.; Farrell, C.L. Differential regulation of leptin transport by the choroid plexus and blood-brain barrier and high affinity transport systems for entry into hypothalamus and across the blood-cerebrospinal fluid barrier. Endocrinology 2000, 141, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Cheunsuang, O.; Morris, R. Astrocytes in the arcuate nucleus and median eminence that take up a fluorescent dye from the circulation express leptin receptors and neuropeptide Y Y1 receptors. Glia 2005, 52, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Gertler, A.; Solomon, G. Leptin-activity blockers: Development and potential use in experimental biology and medicine. Can. J. Physiol. Pharmacol. 2013, 91, 873–882. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | PrimerSequence (5′–3′) | Probe Sequence (5′–3′) | Amplicon Size | Gen Bank Accession Number |
|---|---|---|---|---|
| CPH | CGGCTCCCAGTTCTTCATCA | FAM-CGTTCCGACTCCGC-MGB | 64 bp | D14074 |
| ACTACGTGCTTCCCATCCAAA | ||||
| LRa | TCAAAGTATGTCCGTTCTCTTCTG | FAM-TGTTTTGGGAAGATGTTC-MGB | 131 bp | NM_001009763.1 |
| TCTTATTGCTTGGAACATTGTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biernat, W.; Szczęsna, M.; Kirsz, K.; Zieba, D.A. Seasonal and Nutritional Fluctuations in the mRNA Levels of the Short Form of the Leptin Receptor (LRa) in the Hypothalamus and Anterior Pituitary in Resistin-Treated Sheep. Animals 2021, 11, 2451. https://doi.org/10.3390/ani11082451
Biernat W, Szczęsna M, Kirsz K, Zieba DA. Seasonal and Nutritional Fluctuations in the mRNA Levels of the Short Form of the Leptin Receptor (LRa) in the Hypothalamus and Anterior Pituitary in Resistin-Treated Sheep. Animals. 2021; 11(8):2451. https://doi.org/10.3390/ani11082451
Chicago/Turabian StyleBiernat, Weronika, Malgorzata Szczęsna, Katarzyna Kirsz, and Dorota Anna Zieba. 2021. "Seasonal and Nutritional Fluctuations in the mRNA Levels of the Short Form of the Leptin Receptor (LRa) in the Hypothalamus and Anterior Pituitary in Resistin-Treated Sheep" Animals 11, no. 8: 2451. https://doi.org/10.3390/ani11082451
APA StyleBiernat, W., Szczęsna, M., Kirsz, K., & Zieba, D. A. (2021). Seasonal and Nutritional Fluctuations in the mRNA Levels of the Short Form of the Leptin Receptor (LRa) in the Hypothalamus and Anterior Pituitary in Resistin-Treated Sheep. Animals, 11(8), 2451. https://doi.org/10.3390/ani11082451