
The Invasive Round Goby Neogobius melanostomus as a Potential Threat to Native Crayfish Populations


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Predator and Prey Acquisition and Acclimatization
2.2. Experiment Design
2.3. Data Analysis
3. Results
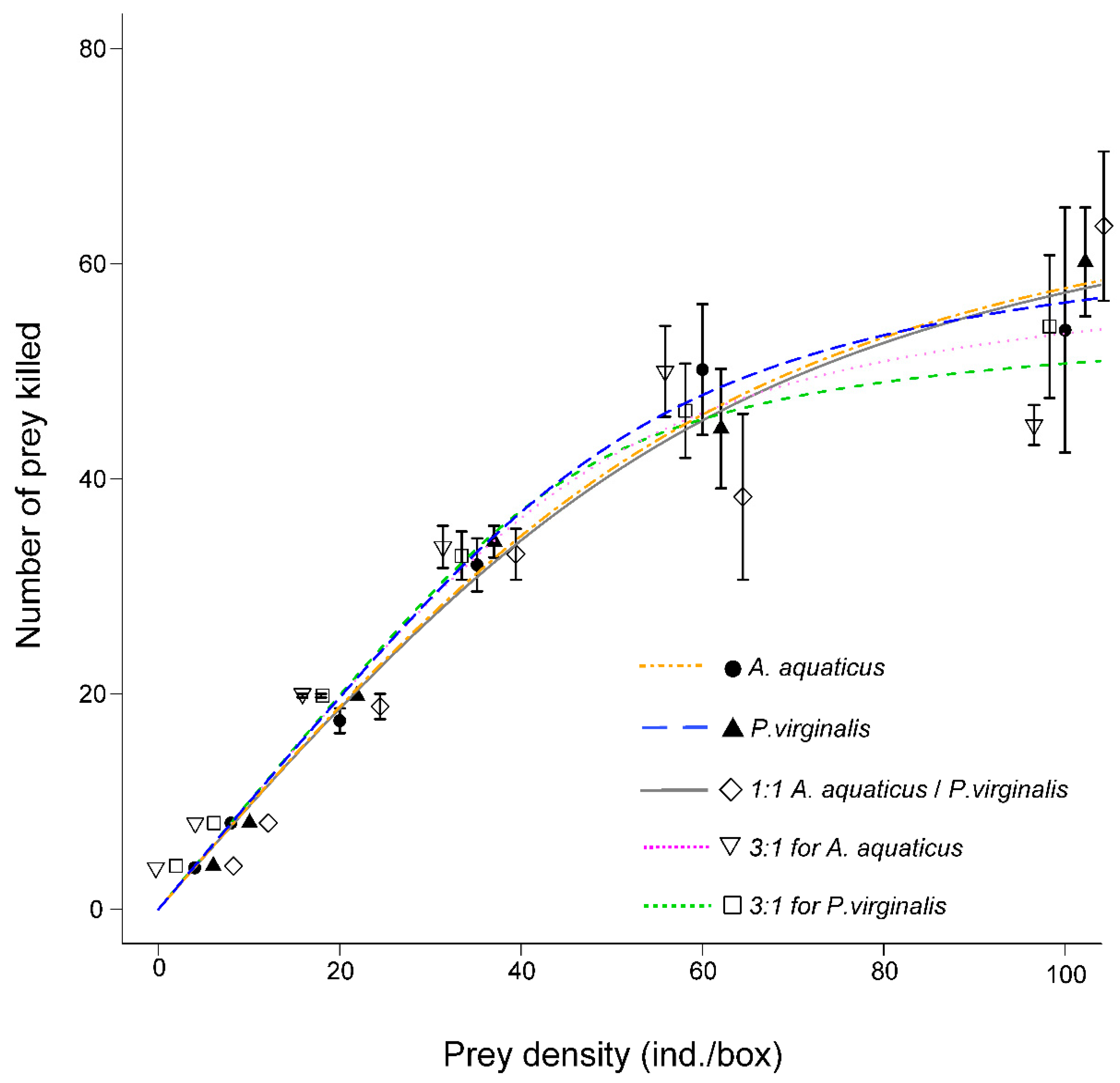
3.1. Functional Response Type
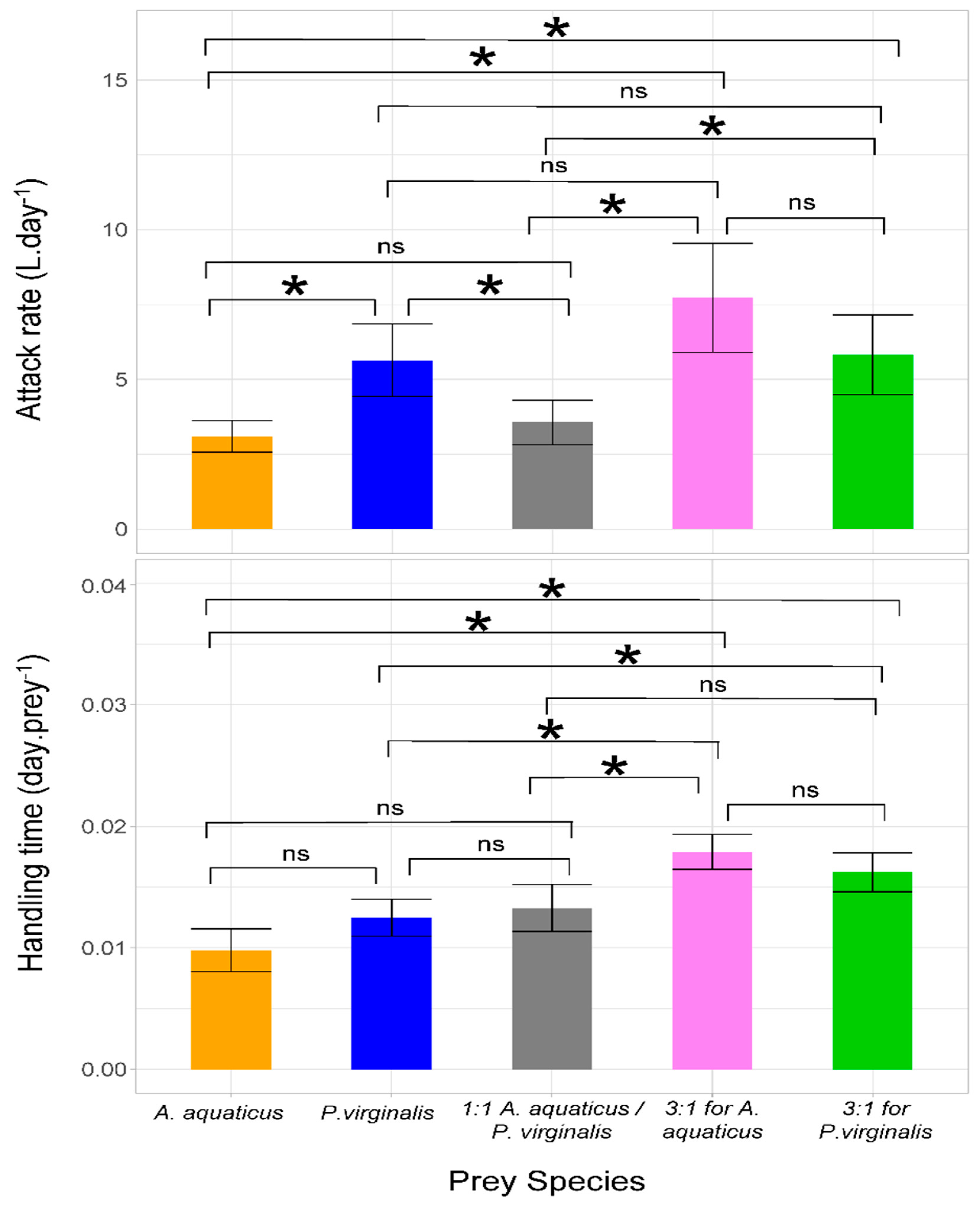
3.2. Attack Rate and Handling Time
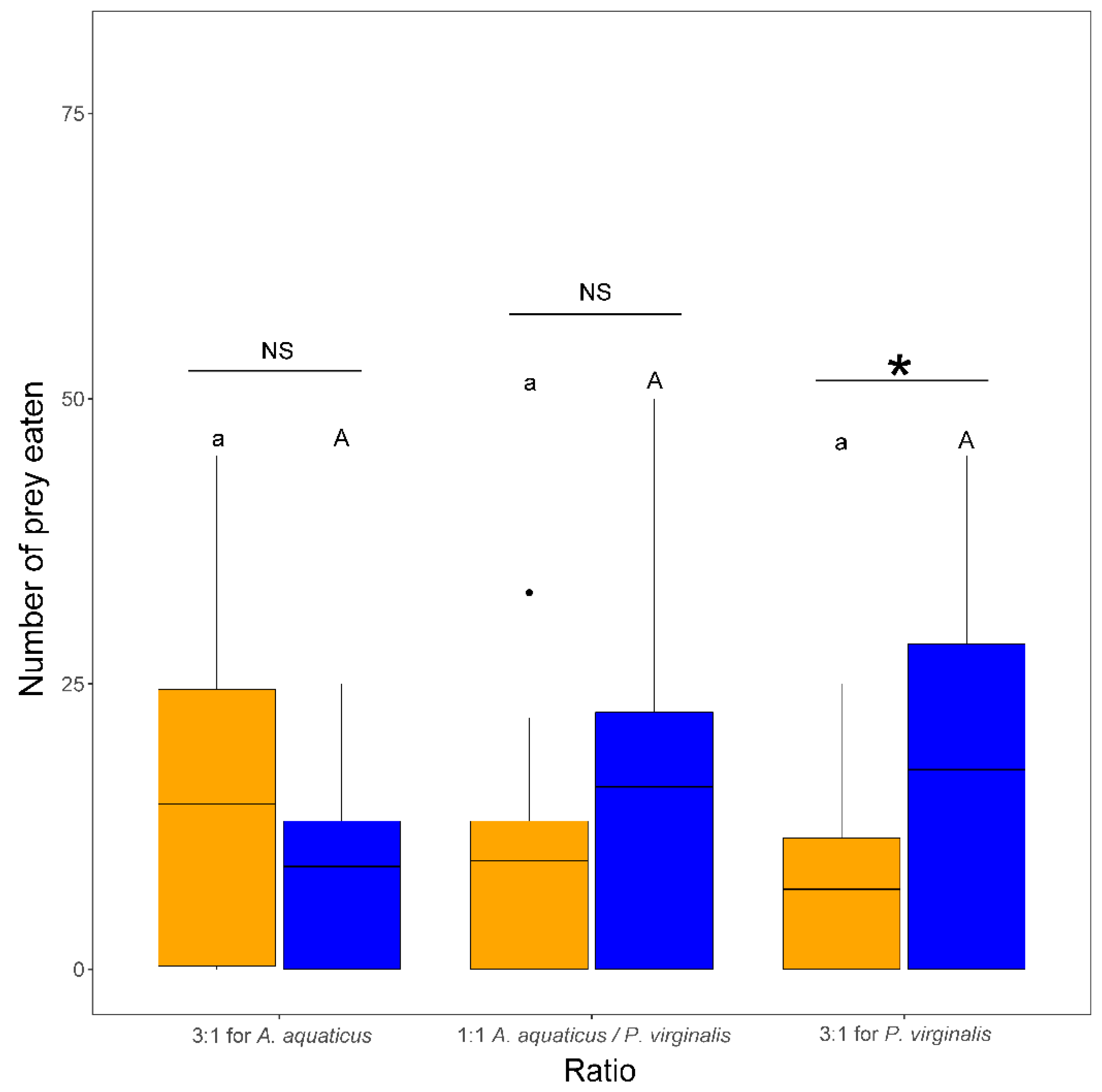
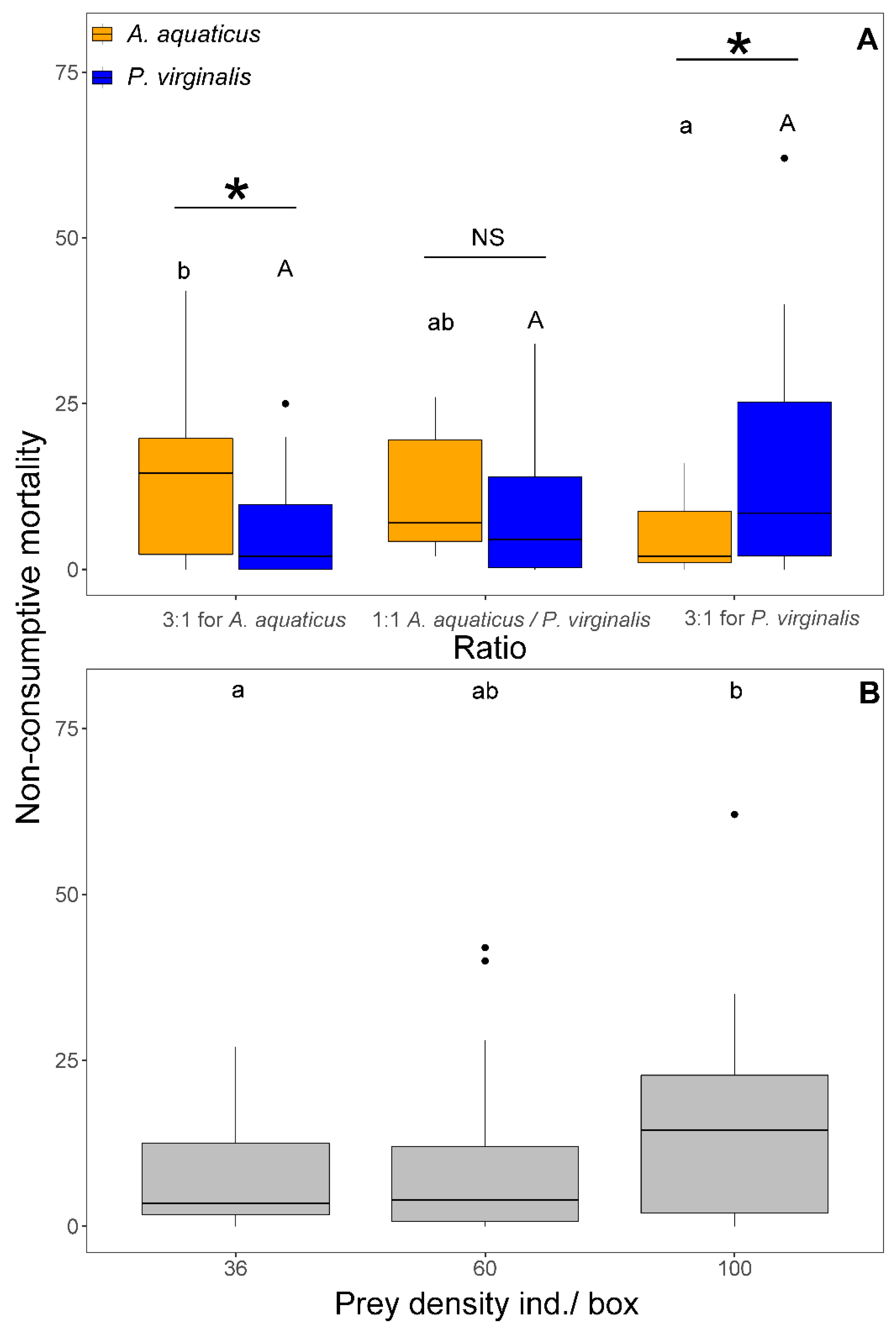
3.3. Number of Killed and Eaten Prey and Non-Consumptive Mortality
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dorn, N.; Mittelbach, G. More than predator and prey: A review of interactions between fish and crayfish. Vie Milieu 1999, 49, 229–237. [Google Scholar]
- Creed, R.P., Jr.; Reed, J.M. Ecosystem engineering by crayfish in a headwater stream community. J. N. Am. Benthol. Soc. 2004, 23, 224–236. [Google Scholar] [CrossRef]
- Dorn, N.J.; Wojdak, J.M. The role of omnivorous crayfish in littoral communities. Oecologia 2004, 140, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Stenroth, P.; Holmqvist, N.; Nyström, P.; Berglund, O.; Larsson, P.; Granéli, W. The influence of productivity and width of littoral zone on the trophic position of a large-bodied omnivore. Oecologia 2008, 156, 681–690. [Google Scholar] [CrossRef]
- Edwards, B.A.; Jackson, D.A.; Somers, K.M. Multispecies crayfish declines in lakes: Implications for species distributions and richness. J. N. Am. Benthol. Soc. 2009, 28, 719–732. [Google Scholar] [CrossRef]
- Reynolds, J.; Souty-Grosset, C.; Richardson, A. Ecological roles of crayfish in freshwater and terrestrial habitats. Freshw. Crayfish 2013, 19, 197–218. [Google Scholar]
- Richman, N.I.; Böhm, M.; Adams, S.B.; Alvarez, F.; Bergey, E.A.; Bunn, J.J.S.; Burnham, Q.; Cordeiro, J.; Coughran, J.; Crandall, K.A.; et al. Multiple drivers of decline in the global status of freshwater crayfish (Decapoda: Astacidea). Philos. Trans. R. Soc. B 2015, 370, 20140060. [Google Scholar] [CrossRef]
- European Parliament and of the Council on Minimum Requirements for Water Reuse. 337 Final 2018/0169 (COD) Brussels. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52018PC0337 (accessed on 28 May 2018).
- Ding, L.; Chen, L.; Ding, C.; Tao, J. Global trends in dam removal and related research: A systematic review based on associated datasets and bibliometric analysis. Chin. Geogr. Sci. 2019, 29, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wohl, E.; Lane, S.N.; Wilcox, A.C. The science and practice of river restoration. Water Resour. Res. 2015, 51, 5974–5997. [Google Scholar] [CrossRef] [Green Version]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Vilà, M.; Basnou, C.; Gollasch, S.; Josefsson, M.; Pergl, J.; Scalera, R. One hundred of the most invasive alien species in Europe. In Handbook of Alien Species in Europe; Hulme, P.E., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 265–268. [Google Scholar]
- Kornis, M.S.; Mercado-Silva, N.; Vander Zanden, M.J. Twenty years of invasion: A review of round goby Neogobius melanostomus biology, spread and ecological implications. J. Fish Biol. 2012, 80, 235–285. [Google Scholar] [CrossRef]
- French, J.R.; Jude, D.J. Diets and diet overlap of nonindigenous gobies and small benthic native fishes co-inhabiting the St. Clair River, Michigan. J. Great Lakes Res. 2001, 27, 300–311. [Google Scholar] [CrossRef]
- Almqvist, G.; Strandmark, A.K.; Appelberg, M. Has the invasive round goby caused new links in Baltic food webs? Environ. Biol. Fishes 2010, 89, 79–93. [Google Scholar] [CrossRef]
- Emde, S.; Rueckert, S.; Kochmann, J.; Knopf, K.; Sures, B.; Klimpel, S. Nematode eel parasite found inside acanthocephalan cysts-a Trojan horse strategy? Parasite Vector 2014, 7, 1–5. [Google Scholar]
- Pagnucco, K.S.; Remmal, Y.; Ricciardi, A. An invasive benthic fish magnifies trophic cascades and alters pelagic communities in an experimental freshwater system. Freshw. Sci. 2016, 35, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Janáč, M.; Šlapanský, L.; Valová, Z.; Jurajda, P. Downstream drift of round goby (Neogobius melanostomus) and tubenose goby (Proterorhinus semilunaris) in their non-native area. Ecol. Freshw. Fish 2013, 22, 430–438. [Google Scholar] [CrossRef]
- Šlapanský, L.; Janáč, M.; Roche, K.; Mikl, L.; Jurajda, P. Expansion of round gobies in a non-navigable river system. Limnologica 2017, 67, 27–36. [Google Scholar] [CrossRef]
- Kornis, M.S.; Vander Zanden, M.J. Forecasting the distribution of the invasive round goby (Neogobius melanostomus) in Wisconsin tributaries to Lake Michigan. Can. J. Fish. Aquat. Sci. 2010, 67, 553–562. [Google Scholar] [CrossRef] [Green Version]
- Poos, M.; Dextrase, A.J.; Schwalb, A.N.; Ackerman, J.D. Secondary invasion of the round goby into high diversity Great Lakes tributaries and species at risk hotspots: Potential new concerns for endangered freshwater species. Biol. Invasions 2010, 12, 1269–1284. [Google Scholar] [CrossRef]
- Verliin, A.; Kesler, M.; Svirgsden, R.; Taal, I.; Saks, L.; Rohtla, M.; Hubel, K.; Eschbaum, R.; Vetemaa, M.; Saat, T. Invasion of round goby to the temperate salmonid streams in the Baltic Sea. Ichthyol. Res. 2017, 64, 155–158. [Google Scholar] [CrossRef]
- Meyer, J.L.; Strayer, D.L.; Wallace, J.B.; Eggert, S.L.; Helfman, G.S.; Leonard, N.E. The contribution of headwater streams to biodiversity in river networks. JAWRA J. Am. Water Resour. 2017, 43, 86–103. [Google Scholar] [CrossRef] [Green Version]
- Bottcher, J.L.; Walsworth, T.E.; Thiede, G.P.; Budy, P.; Speas, D.W. Frequent usage of tributaries by the endangered fishes of the upper Colorado River basin: Observations from the San Rafael River, Utah. N. Am. J. Fish. Manag. 2013, 33, 585–594. [Google Scholar] [CrossRef]
- Kuhns, L.A.; Berg, M.B. Benthic invertebrate community responses to round goby (Neogobius melanostomus) and zebra mussel (Dreissena polymorpha) invasion in southern Lake Michigan. J. Great Lakes Res. 1999, 25, 910–917. [Google Scholar] [CrossRef]
- Lederer, A.M.; Janssen, J.; Reed, T.; Wolf, A. Impacts of the introduced round goby (Apollonia melanostoma) on dreissenids (Dreissena polymorpha and Dreissena bugensis) and on macroinvertebrate community between 2003 and 2006 in the littoral zone of Green Bay, Lake Michigan. J. Great Lakes Res. 2008, 34, 690–697. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Miyasaka, H.; Genkai-Kato, M.; Taniguchi, Y.; Nakano, S. Seasonal change in the gastric evacuation rate of rainbow trout feeding on natural prey. J. Fish Biol. 2007, 71, 1873–1878. [Google Scholar] [CrossRef]
- Perello, M.M.; Simon, T.P.; Thompson, H.M.; Kane, D.D. Feeding ecology of the invasive round goby, Neogobius melanostomus (Pallas, 1814), based on laboratory size preference and field diet in different habitats in the western basin of Lake Erie. Aquat. Invasions 2015, 10, 463–474. [Google Scholar] [CrossRef]
- Polačik, M.; Janáč, M.; Jurajda, P.; Adámek, Z.; Ondračková, M.; Trichkova, T.; Vassilev, M. Invasive gobies in the Danube: Invasion success facilitated by availability and selection of superior food resources. Ecol. Freshw. Fish 2009, 18, 640–649. [Google Scholar] [CrossRef]
- Pennuto, C.; Krakowiak, P.; Janik, C. Seasonal abundance, diet, and energy consumption of round gobies (Neogobius melanostomus) in Lake Erie tributary streams. Ecol. Freshw. Fish 2010, 19, 206–215. [Google Scholar] [CrossRef]
- Townsend, C.R.; Winfield, I.J. The application of optimal foraging theory to feeding behaviour in fish. In Fish Energetics: New Perspectives, 1st ed.; Tytler, P., Calow, P., Eds.; Croom Helm: Sydney, Australia, 1985; pp. 67–98. [Google Scholar]
- Shemonaev, E.; Kirilenko, E. Features of biology of the round goby Neogobius melanostomus (Perciformes, Gobiidae) in waters of Kuibyshev Reservoir. J. Ichthyol. 2009, 49, 454–459. [Google Scholar] [CrossRef]
- Kirilenko, E.; Shemonaev, E. Feeding of the round goby Neogobius melanostomus (Perciformes, Gobiidae) in two Volga reservoirs. J. Ichthyol. 2012, 52, 291–295. [Google Scholar] [CrossRef]
- Herberholz, J.; Sen, M.M.; Edwards, D.H. Escape behavior and escape circuit activation in juvenile crayfish during prey–predator interactions. J. Exp. Biol. 2004, 207, 1855–1863. [Google Scholar] [CrossRef] [Green Version]
- Stein, R.A. Selective predation, optimal foraging, and the predator-prey interaction between fish and crayfish. Ecology 1977, 58, 1237–1253. [Google Scholar] [CrossRef]
- Sandeman, R.; Sandeman, D. Development, growth, and plasticity in the crayfish olfactory system. Microsc. Res. Tech. 2003, 60, 266–277. [Google Scholar] [CrossRef]
- Lang, F.; Govind, C.; Costello, W.J.; Greene, S.I. Developmental neuroethology: Changes in escape and defensive behavior during growth of the lobster. Science 1977, 197, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Kellie, S.; Greer, J.; Cooper, R.L. Alterations in habituation of the tail flip response in epigean and troglobitic crayfish. J. Exp. Zool. 2001, 290, 163–176. [Google Scholar] [CrossRef]
- Olden, J.D.; Poff, N.L.; Douglas, M.R.; Douglas, M.E.; Fausch, K.D. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol. Evol. 2004, 19, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Dick, J.T.; Platvoet, D.; Kelly, D.W. Predatory impact of the freshwater invader Dikerogammarus villosus (Crustacea: Amphipoda). Can. J. Fish. Aquat. Sci. 2002, 59, 1078–1084. [Google Scholar] [CrossRef] [Green Version]
- Dick, J.T.; Gallagher, K.; Avlijas, S.; Clarke, H.C.; Lewis, S.E.; Leung, S.; Minchin, D.; Caffrey, J.; Alexander, M.E.; Maguire, C. Ecological impacts of an invasive predator explained and predicted by comparative functional responses. Biol. Invasions 2013, 15, 837–846. [Google Scholar] [CrossRef]
- Moorhouse, T.P.; Macdonald, D.W. Are invasives worse in freshwater than terrestrial ecosystems? WIREs Water 2015, 2, 1–8. [Google Scholar] [CrossRef]
- Solomon, M. The natural control of animal populations. J. Anim. Ecol. 1949, 18, 1–35. [Google Scholar] [CrossRef]
- Holling, C.S. Some Characteristics of Simple Types of Predation and Parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Murdoch, W.W.; Oaten, A. Predation and population stability. Adv. Ecol. Res. 1975, 9, 1–131. [Google Scholar]
- Juliano, S.A. Nonlinear curve fitting: Predation and functional response curve. In Design and Analysis of Ecological Experiments, 2nd ed.; Scheiner, S.M., Gurevitch, J., Eds.; Chapman and Hall: New York, NY, USA, 2001; pp. 178–196. [Google Scholar]
- Bollache, L.; Dick, J.T.; Farnsworth, K.D.; Montgomery, W.I. Comparison of the functional responses of invasive and native amphipods. Biol. Lett. 2008, 4, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, M.E.; Dick, J.T.; Weyl, O.L.; Robinson, T.B.; Richardson, D.M. Existing and emerging high impact invasive species are characterized by higher functional responses than natives. Biol. Lett. 2014, 10, 20130946. [Google Scholar] [CrossRef] [PubMed]
- Laverty, C.; Green, K.D.; Dick, J.T.; Barrios-O’Neill, D.; Mensink, P.J.; Médoc, V.; Spataro, T.; Caffrey, J.M.; Lucy, F.E.; Boets, P. Assessing the ecological impacts of invasive species based on their functional responses and abundances. Biol. Invasions 2017, 19, 1653–1665. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Mu, X.; Dick, J.T.; Fang, M.; Gu, D.; Luo, D.; Zhang, J.; Luo, J.; Hu, Y. Comparative functional responses predict the invasiveness and ecological impacts of alien herbivorous snails. PLoS ONE 2016, 11, e0147017. [Google Scholar] [CrossRef] [Green Version]
- Dubs, D.O.; Corkum, L.D. Behavioral interactions between round gobies (Neogobius melanostomus) and mottled sculpins (Cottus bairdi). J. Great Lakes Res. 1996, 22, 838–844. [Google Scholar] [CrossRef]
- Gebauer, R.; Veselý, L.; Kouba, A.; Buřič, M.; Drozd, B. Forecasting impact of existing and emerging invasive gobiids under temperature change using comparative functional responses. Aquat. Invasions 2018, 13, 289–297. [Google Scholar] [CrossRef]
- Colton, T.F. Extending functional response models to include a second prey type: An experimental test. Ecology 1987, 68, 900–912. [Google Scholar] [CrossRef]
- Dodd, J.A.; Dick, J.T.A.; Alexander, M.E.; Macneil, C.; Dunn, A.M.; Aldridge, D.C. Predicting the ecological impacts of a new freshwater invader: Functional responses and prey selectivity of the ‘killer shrimp’, Dikerogammarus villosus, compared to the native Gammarus pulex. Freshw. Biol. 2014, 59, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, R.N.; Dickey, J.W.; McMorrow, C.; Laverty, C.; Dick, J.T. Resistance is futile: Lack of predator switching and a preference for native prey predict the success of an invasive prey species. R. Soc. Open Sci. 2018, 5, 180339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudová, P.; Boukal, D.S.; Klecka, J. Prey selectivity and the effect of diet on growth and development of a dragonfly, Sympetrum sanguineum. PeerJ. 2019, 7, e7881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- South, J.; McCard, M.; Khosa, D.; Mofu, M.; Madzivanzira, T.C.; Dick, J.T.; Weyl, O.L. The effect of prey identity and substrate type on the functional response of a globally invasive crayfish. NeoBiota 2019, 52, 9. [Google Scholar] [CrossRef] [Green Version]
- Vašek, M.; Všetičková, L.; Roche, K.; Jurajda, P. Diet of two invading gobiid species (Proterorhinus semilunaris and Neogobius melanostomus) during the breeding and hatching season: No field evidence of extensive predation on fish eggs and fry. Limnologica 2014, 46, 31–36. [Google Scholar] [CrossRef]
- Hempel, M.; Magath, V.; Neukamm, R.; Thiel, R. Feeding ecology, growth and reproductive biology of round goby Neogobius melanostomus (Pallas, 1814) in the brackish Kiel Canal. Mar. Biodivers. 2018, 49, 795–807. [Google Scholar] [CrossRef]
- Hay, A.M. Foraging Behaviour of the Ruffe (Gymnocephalus cernuus) and Predator Avoidance by the Freshwater Isopod Asellus aquaticus: Implication for Predator-Prey Interaction. Ph.D. Thesis, University of Glasgow, Glasgow, UK, September 1999. [Google Scholar]
- Hossain, M.S.; Patoka, J.; Kouba, A.; Buřič, M. Clonal crayfish as biological model: A review on marbled crayfish. Biologia 2018, 73, 841–855. [Google Scholar] [CrossRef]
- Kouba, A.; Petrusek, A.; Kozák, P. Continental-wide distribution of crayfish species in Europe: Update and maps. Knowl. Manag. Aquat. Ecosyst. 2017, 413, 05. [Google Scholar] [CrossRef]
- Fantinou, A.; Perdikis, D.C.; Maselou, D.; Lambropoulos, P. Prey killing without consumption: Does Macrolophus pygmaeus show adaptive foraging behaviour? Biol. Control 2008, 47, 187–193. [Google Scholar] [CrossRef]
- Rogers, D. Random search and insect population models. J. Anim. Ecol. 1972, 369–383. [Google Scholar] [CrossRef]
- Holling, C.S. The Functional Response of Predators to Prey Density and its Role in Mimicry and Population Regulation. Mem. Entomol. Soc. Can. 1965, 97, 5–60. [Google Scholar] [CrossRef] [Green Version]
- Bolker, B. Ecological Models and Data in R; Princeton University Press: Princeton, NJ, USA, 2008; pp. 127–135. [Google Scholar]
- Sentis, A.; Hemptinne, J.; Brodeur, J. How functional response and productivity modulate intraguild predation. Ecosphere 2013, 4, 1–14. [Google Scholar] [CrossRef]
- David, P.; Thebault, E.; Anneville, O.; Duyck, P.F.; Chapuis, E.; Loeuille, N. Impacts of invasive species on food webs: A review of empirical data. Adv. Ecol. Res. 2017, 56, 1–60. [Google Scholar]
- Mikl, L.; Adámek, Z.; Všetičková, L.; Janáč, M.; Roche, K.; Šlapanský, L.; Jurajda, P. Response of benthic macroinvertebrate assemblages to round (Neogobius melanostomus, Pallas 1814) and tubenose (Proterorhinus semilunaris, Heckel 1837) goby predation pressure. Hydrobiologia 2017, 785, 219–232. [Google Scholar] [CrossRef]
- Gebauer, R.; Divíšek, J.; Buřič, M.; Večeřa, M.; Kouba, A.; Drozd, B. Distribution of alien animal species richness in the Czech Republic. Ecol. Evol. 2018, 8, 4455–4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeuwen, E.V.; Jansen, V.; Bright, P. How population dynamics shape the functional response in a one-predator–two-prey system. Ecology 2007, 88, 1571–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dick, J.T.; Alexander, M.E.; Jeschke, J.M.; Ricciardi, A.; MacIsaac, H.J.; Robinson, T.B.; Kumschick, S.; Weyl, O.L.; Dunn, A.M.; Hatcher, M.J. Advancing impact prediction and hypothesis testing in invasion ecology using a comparative functional response approach. Biol. Invasions 2014, 16, 735–753. [Google Scholar] [CrossRef] [Green Version]
- Paton, R.A.; Gobin, J.; Rooke, A.C.; Fox, M.G. Population density contributes to the higher functional response of an invasive fish. Biol. Invasions 2019, 21, 1737–1749. [Google Scholar] [CrossRef]
- Alexander, M.E.; Dick, J.T.; O’Connor, N.E.; Haddaway, N.R.; Farnsworth, K.D. Functional responses of the intertidal amphipod Echinogammarus marinus: Effects of prey supply, model selection and habitat complexity. Mar. Ecol. Prog. Ser. 2012, 468, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Akre, B.G.; Johnson, D.M. Switching and sigmoid functional response curves by damselfly naiads with alternative prey available. J. Anim. Ecol. 1979, 703–720. [Google Scholar] [CrossRef]
- Hammill, E.; Petchey, O.L.; Anholt, B.R. Predator functional response changed by induced defenses in prey. Am. Nat. 2010, 176, 723–731. [Google Scholar] [CrossRef] [Green Version]
- Gebauer, R.; Veselý, L.; Vanina, T.; Buřič, M.; Kouba, A.; Drozd, B. Prediction of ecological impact of two alien gobiids in habitat structures of differing complexity. Can. J. Fish. Aquat. Sci. 2019, 76, 1954–1961. [Google Scholar] [CrossRef]
- Alexander, M.; Kaiser, H.; Weyl, O.; Dick, J. Habitat simplification increases the impact of a freshwater invasive fish. Environ. Biol. Fishes 2015, 98, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Jeschke, J.M.; Tollrian, R. Density-dependent effects of prey defences. Oecologia 2000, 123, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Blake, M.; Hart, P. The vulnerability of juvenile signal crayfish to perch and eel predation. Freshw. Biol. 1995, 33, 233–244. [Google Scholar] [CrossRef]
- Webb, P. Effect of body form and response threshold on the vulnerability of four species of teleost prey attacked by largemouth bass (Micropterus salmoides). Can. J. Fish. Aquat. Sci. 1986, 43, 763–771. [Google Scholar] [CrossRef]
- Sih, A.; Christensen, B. Optimal diet theory: When does it work, and when and why does it fail? Anim. Behav. 2001, 61, 379–390. [Google Scholar] [CrossRef]
- Lawton, J.; Beddington, J.; Bonser, R. Switching in invertebrate predators. In Ecological Stability; Usher, M.B., Williamson, M.H., Eds.; Chapman and Hall: London, UK, 1974; pp. 141–158. [Google Scholar]
- Kipp, R.; Hébert, I.; Lacharité, M.; Ricciardi, A. Impacts of predation by the Eurasian round goby (Neogobius melanostomus) on molluscs in the upper St. Lawrence River. J. Great Lakes Res. 2012, 38, 78–89. [Google Scholar] [CrossRef]
- Bhagat, Y.; Ruetz, C.R., III; Akins, A.L. Differential habitat use by the round goby (Neogobius melanostomus) and Dreissena spp. in coastal habitats of eastern Lake Michigan. J. Great Lakes Res. 2015, 41, 1087–1093. [Google Scholar] [CrossRef]
- Tytler, P.; Calow, P. Fish Energetics: New Perspectives; Croom Helm: Sydney, Australia, 1985; p. 348. [Google Scholar]
- Hart, P.; Ison, S. The influence of prey size and abundance, and individual phenotype on prey choice by the three-spined stickleback, Gasterosteus aculeatus L. J. Fish Biol. 1991, 38, 359–372. [Google Scholar] [CrossRef]
- Kislalioglu, M.; Gibson, R. Prey ‘handling time’and its importance in food selection by the 15-spined stickleback, Spinachia spinachia (L.). J. Exp. Mar. Biol. Ecol. 1976, 25, 151–158. [Google Scholar] [CrossRef]
- Diggins, T.P.; Kaur, J.; Chakraborti, R.K.; DePinto, J.V. Diet choice by the exotic round goby (Neogobius melanostomus) as influenced by prey motility and environmental complexity. J. Great Lakes Res. 2002, 28, 411–420. [Google Scholar] [CrossRef]
- Sohel, S.; Mattila, J.; Lindström, K. Effects of turbidity on prey choice of three-spined stickleback Gasterosteus aculeatus. Mar. Ecol. Prog. Ser. 2017, 566, 159–167. [Google Scholar] [CrossRef]
- Błońska, D.; Grabowska, J.; Kobak, J.; Jermacz, Ł.; Bącela-Spychalska, K. Feeding preferences of an invasive Ponto-Caspian goby for native and non-native gammarid prey. Freshw. Biol. 2015, 60, 2187–2195. [Google Scholar] [CrossRef]
- Beggel, S.; Brandner, J.; Cerwenka, A.; Geist, J. Synergistic impacts by an invasive amphipod and an invasive fish explain native gammarid extinction. BMC Ecol. 2016, 16, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandner, J.; Auerswald, K.; Cerwenka, A.F.; Schliewen, U.K.; Geist, J. Comparative feeding ecology of invasive Ponto-Caspian gobies. Hydrobiologia 2013, 703, 113–131. [Google Scholar] [CrossRef]
- Carman, S.M.; Janssen, J.; Jude, D.J.; Berg, M.B. Diel interactions between prey behaviour and feeding in an invasive fish, the round goby, in a North American river. Freshw. Biol. 2006, 51, 742–755. [Google Scholar] [CrossRef]
- Siepielski, A.M.; Wang, J.; Prince, G. Nonconsumptive predator-driven mortality causes natural selection on prey. Evolution 2014, 68, 696–704. [Google Scholar] [CrossRef]
- Preisser, E.L.; Bolnick, D.I.; Benard, M.F. Scared to death? The effects of intimidation and consumption in predator–prey interactions. Ecology 2005, 86, 501–509. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.M.; Akre, B.G.; Crowley, P.H. Modeling arthropod predation: Wasteful killing by damselfly naiads. Ecology 1975, 56, 1081–1093. [Google Scholar] [CrossRef]
- Veselý, L.; Boukal, D.S.; Buřič, M.; Kozák, P.; Kouba, A.; Sentis, A. Effects of prey density, temperature and predator diversity on nonconsumptive predator-driven mortality in a freshwater food web. Sci. Rep. 2017, 7, 18075. [Google Scholar] [CrossRef]
- Kruuk, H. Surplus killing by carnivores. J. Zool. 1972, 166, 233–244. [Google Scholar] [CrossRef]
- Jedrzejewska, B.; Jedrzejewski, W. Seasonal surplus killing as hunting strategy of the weasel Mustela nivalis-test of a hypothesis. Acta Theriol. 1989, 34, 347–360. [Google Scholar] [CrossRef] [Green Version]
- Short, J.; Kinnear, J.; Robley, A. Surplus killing by introduced predators in Australia—Evidence for ineffective anti-predator adaptations in native prey species? Biol. Conserv. 2002, 103, 283–301. [Google Scholar] [CrossRef]
- McKee, M.; Wrona, F.; Scrimgeour, G.; Culp, J. Importance of consumptive and non-consumptive prey mortality in a coupled predator–prey system. Freshw. Biol. 1997, 38, 193–201. [Google Scholar] [CrossRef]
- Holling, C.S. The functional response of invertebrate predators to prey density. Mem. Entomol. Soc. Can. 1966, 98, 5–86. [Google Scholar] [CrossRef]
- Lee, V.A.; Johnson, T.B. Development of a bioenergetics model for the round goby (Neogobius melanostomus). J. Great Lakes Res. 2005, 31, 125–134. [Google Scholar] [CrossRef]
- Church, K.; Iacarella, J.C.; Ricciardi, A. Aggressive interactions between two invasive species: The round goby (Neogobius melanostomus) and the spinycheek crayfish (Orconectes limosus). Biol. Invasions 2017, 19, 425–441. [Google Scholar] [CrossRef]
- Bovy, H.C.; Barrios-O’Neill, D.; Emmerson, M.C.; Aldridge, D.C.; Dick, J.T. Predicting the predatory impacts of the “demon shrimp” Dikerogammarus haemobaphes, on native and previously introduced species. Biol. Invasions 2015, 17, 597–607. [Google Scholar] [CrossRef]
- Martin, P. Parthenogenesis: Mechanisms, evolution, and its relevance to the role of marbled crayfish as model organism and potential invader. In Freshwater Crayfish: A Global Overview; Kawai, T., Faulkes, Z., Scholtz, G., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 63–82. [Google Scholar]
- Holdich, D.; Reynolds, J.; Souty-Grosset, C.; Sibley, P. A review of the ever increasing threat to European crayfish from non-indigenous crayfish species. Knowl. Managt. Aquat. Ecosyst. 2009, 11, 394–395. [Google Scholar] [CrossRef] [Green Version]
- Pennuto, C.; Rupprecht, S. Upstream range expansion by invasive round gobies: Is functional morphology important? Aquat. Ecol. 2016, 50, 45–57. [Google Scholar] [CrossRef]
- Kornis, M.S.; Weidel, B.C.; Vander Zanden, M.J. Divergent life histories of invasive round gobies (Neogobius melanostomus) in Lake Michigan and its tributaries. Ecol. Freshw. Fish 2017, 26, 563–574. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure | Linear Coefficient P1 | SE | p-Value |
|---|---|---|---|
| A. aquaticus | −1.145 | 0.364 | 0.002 |
| P. virginalis | −1.107 | 0.360 | 0.002 |
| 1:1 A. aquaticus/P. virginalis | −1.047 | 0.360 | 0.004 |
| 3:1 for A. aquaticus | −1.580 | 0.365 | <10−4 |
| 3:1 for P. virginalis | −1.302 | 0.361 | <10−3 |
| Parameter | Prey | Lower Limit of 95% CI | Mean | Upper Limit of 95% CI | p-Value |
|---|---|---|---|---|---|
| Attack rate | A. aquaticus | 2.573 | 3.094 | 3.615 | <10−6 |
| P. virginalis | 4.433 | 5.640 | 6.848 | <10−6 | |
| 1:1 A. aquaticus/P. virginalis | 2.830 | 3.568 | 4.307 | <10−6 | |
| 3:1 for A. aquaticus | 5.900 | 7.724 | 9.548 | <10−6 | |
| 3:1 for P. virginalis | 4.491 | 5.825 | 7.158 | <10−6 | |
| Handling time | A. aquaticus | 0.008 | 0.010 | 0.012 | <10−6 |
| P. virginalis | 0.011 | 0.012 | 0.014 | <10−6 | |
| 1:1 A. aquaticus/P. virginalis | 0.011 | 0.013 | 0.015 | <10−6 | |
| 3:1 for A. aquaticus | 0.016 | 0.018 | 0.019 | <10−6 | |
| 3:1 for P. virginalis | 0.014 | 0.016 | 0.018 | <10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franta, P.; Gebauer, R.; Veselý, L.; Buřič, M.; Szydłowska, N.Z.; Drozd, B. The Invasive Round Goby Neogobius melanostomus as a Potential Threat to Native Crayfish Populations. Animals 2021, 11, 2377. https://doi.org/10.3390/ani11082377
Franta P, Gebauer R, Veselý L, Buřič M, Szydłowska NZ, Drozd B. The Invasive Round Goby Neogobius melanostomus as a Potential Threat to Native Crayfish Populations. Animals. 2021; 11(8):2377. https://doi.org/10.3390/ani11082377
Chicago/Turabian StyleFranta, Pavel, Radek Gebauer, Lukáš Veselý, Miloš Buřič, Natalia Z. Szydłowska, and Bořek Drozd. 2021. "The Invasive Round Goby Neogobius melanostomus as a Potential Threat to Native Crayfish Populations" Animals 11, no. 8: 2377. https://doi.org/10.3390/ani11082377
APA StyleFranta, P., Gebauer, R., Veselý, L., Buřič, M., Szydłowska, N. Z., & Drozd, B. (2021). The Invasive Round Goby Neogobius melanostomus as a Potential Threat to Native Crayfish Populations. Animals, 11(8), 2377. https://doi.org/10.3390/ani11082377