
Morphological Variations of Wild Populations of Brycon dentex (Characidae, Teleostei) in the Guayas Hydrographic Basin (Ecuador). The Impact of Fishing Policies and Environmental Conditions

 ,
,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
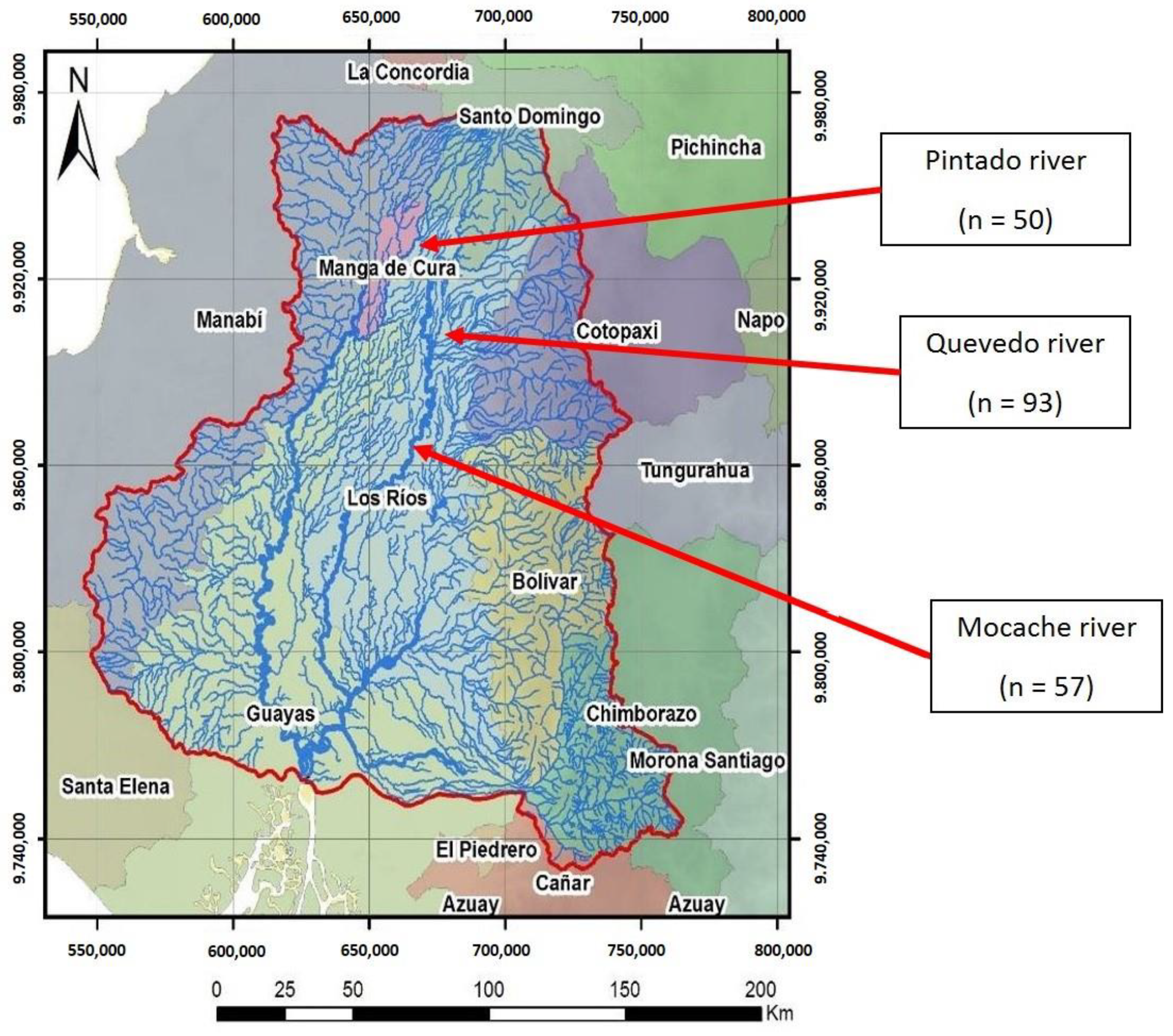
2.1. Data Collection and Study Area
2.2. Body Measurements
2.3. Fulton Condition Coefficient (K)
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schluter, D. The Ecology of Adaptative Radiation; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Ndiwas, T.C.; Nyingi, D.W.; Claude, J.; Agnése, J.F. Morphological variation of wild populations of Nile tilapia (Oreochromis niloticus) living in extreme environmental conditions in the Kenyan Rift-Valley. Environ. Biol Fish. 2016, 99, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Escanta-Molina, R.; Jiménez-Prado, P. Uso de la morfometría geométrica para establecer contrastes biológicos y ambientales en poblaciones de peces del río Teaone. Rev. Cient. Hallazgos 2019, 4, 55–69. [Google Scholar]
- Collyer, M.L.; Stockwell, C.A.; Adams, D.C.; Reiser, M.H. Phenotypic plasticity and contemporary evolution in introduced populations: Evidence from translocated populations of white sands pupfish (Cyrpinodon tularosa). Ecol. Res. 2007, 22, 902–910. [Google Scholar] [CrossRef] [Green Version]
- Gagnaire, P.A.; Normandeau, E.; Pavey, S.A.; Bernatchez, L. Mapping phenotypic, expression and transmission ratio distortion QTL using RAD markers in the Lake Whitefish (Coregonus clupeaformis). Mol. Ecol. 2013, 22, 3036–3048. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Mieles, G.; Irvine, K.; Griensven, A.V.; Arias-Hidalgo, M.; Torres, A.; Mynett, A.E. Relationships between aquatic biotic communities and water quality in a tropical river–wetland system (Ecuador). Environ. Sci. Policy 2013, 34, 115–127. [Google Scholar] [CrossRef]
- Aguilar, C.; González-Sansón, G.; Cabrera, Y.; Ruiz, A.; Allen Curry, R. Inter-habitat variation in density and size composition of reef fishes from the Cuban Northwestern shelf. Rev. Biol. Trop. 2014, 62, 589–602. [Google Scholar]
- Kerezsy, A.; Arthington, A.H.; Balcombe, S.R. Fish Distribution in Far Western Queensland, Australia: The Importance of Habitat, Connectivity and Natural Flows. Diversity 2014, 6, 380–395. [Google Scholar] [CrossRef] [Green Version]
- Arceo-Carranza, D.; Gamboa, E.; Teutli-Hernández, C.; Badillo-Alemán, M.; Herrera-Silveira, J.A. Los peces como indicador de restauración de áreas de manglar en la costa norte de Yucatán. Rev. Mex. Biodivers. 2016, 87, 489–496. [Google Scholar] [CrossRef] [Green Version]
- FAO. El Estado de la Biodiversidad Para la Alimentación y la Agricultura; Comisión de Recursos Genéticos Para la Alimentación y la Agricultura; Evaluaciones: Rome, Italy, 2019. [Google Scholar]
- FAO. Realización de Encuestas y Seguimiento de los Recursos Zoogenéticos; Directrices FAO, Producción y Sanidad Animal: Rome, Italy, 2012. [Google Scholar]
- FAO. Plan de Acción Mundial Sobre los Recursos Zoogenéticos y la Declaración de Interlaken; FAO: Rome, Italy, 2007. [Google Scholar]
- Gonzalez-Martinez, A.; De-Pablos-Heredero, C.; González, M.; Rodriguez, J.; Barba, C.; García, A. Usefulness of discriminant analysis in the morphometric differentiation of six native freshwater species from Ecuador. Animals 2021, 11, 111. [Google Scholar] [CrossRef]
- Gonzalez-Martinez, A.; Lopez, M.; Molero, H.M.; Rodriguez, J.; González, M.; Barba, C.; García, A. Morphometric and Meristic Characterization of Native Chame Fish (Dormitator latifrons) in Ecuador Using Multivariate Analysis. Animals 2020, 10, 1805. [Google Scholar] [CrossRef]
- Cocha-Alulema, A.P. Análisis de la Variación Morfológica de Hoplias Malabaricus (Bloch 1794), Asociada al Tipo de Hábitat Utilizando Morfometría Geométrica. BSc, Facultad de Ciencias Biológicas, Universidad Central del Ecuador: Quito, Ecuador, 2018. Available online: http://www.dspace.uce.edu.ec/handle/25000/15123 (accessed on 14 June 2021).
- Dauda, A.; Abbaya, H.Y.; Ebegbulem, V.N. Application of multifactorial discriminant analysis of morphostructural differentiation of sheep. J. Genet. Genet. Eng. 2018, 2, 11–16. [Google Scholar]
- Siddik, M.A.B.; Hanif, M.A.; Chaklader, M.R.; Nahar, A.; Mahmud, S. Fishery biology of gangetic whiting Sillaginopsis panijus (Hamilton, 1822) endemic to Ganges delta, Bangladesh, Egypt. J. Aquat. Res. 2015, 41, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Hanif, M.A.; Siddik, M.A.B.; Islam, M.A.; Chaklader, M.R.; Nahar, A. Multivariate morphometric variability in sardine, Amblygaster clupeoides (Bleeker, 1849), from the Bay of Bengal coast, Bangladesh. JoBAZ 2019, 80, 53. [Google Scholar] [CrossRef]
- Bilikis Iyabo, U. Length weight relationship, condition factor and diet composition of brycinus nurse (characiformes: Alestidae) in a tropical flood river basin. Cont. J. Fish. Aquat. Sci. 2014, 8, 25–34. [Google Scholar]
- Mazon Paredes, E.; Herrera Rodriguez, M.; Mazon Paredes, C.; Garcia Martinez, A.; Delgado Pertinez, M.; Guzman Guerrero, J.L. Bromatological composition of palm kernel meal according to its origin and production periods potential use of palm kernel meal in animal feed. J. Oil Palm Res. 2020, 32, 639–646. [Google Scholar]
- Teimori, A.; Schulz-Mirbach, T.; Esmaeili, H.R.; Reichenbacher, B. Geographical differentiation of Aphanius dispar (Teleostei: Cyprinodontidae) from southern Iran. J. Zool. Syst. Evol. Res. 2012, 50, 289–304. [Google Scholar] [CrossRef]
- Aguirre, W.W.; Shervette, V.R.; Navarrete, R.; Calle, P.; Agorastos, S. Morphological and Genetic Divergence of Hoplias microlepis (Characiformes: Erythrinidae) in Rivers and Artificial Impoundments of Western Ecuador. Copeia 2013, 2, 312–323. [Google Scholar] [CrossRef]
- Sobczuk, D.; Komosa, M. Morphological differentiation of polish Arabian horses-multivariate analysis. Bull. Vet. Inst. Pulawy 2012, 56, 623–629. [Google Scholar] [CrossRef] [Green Version]
- N’goran, K.E.; Kouassi, N.C.; Loukou, N.E.; Dayo, G.S.M.; Yapi-Gnaore, C.V. Multivariate analysis for morphological caracteristics of N’Dama cattle breed in two agroecological zones of Côte d’Ivoire. Eur. Sci. J. 2018, 14, 602–621. [Google Scholar]
- Hanif, M.A.; Siddik, M.A.B.; Chaklader, M.R.; Pham, H.D.; Kleindienst, R. Length-weight relationships of three catfish species from a tributary of the Dhaleshwari River, Bangladesh. J. Appl. Ichthyol. 2017, 33, 1261–1262. [Google Scholar] [CrossRef]
- Islam, M.A.; Siddik, M.A.B.; Hanif, M.A.; Chaklader, M.R.; Nahar, A.; Ilham, I. Length–weight relationships of four small indigenous fish species from an inland artisanal fishery, Bangladesh. J. Appl. Ichthyol. 2017, 33, 851–852. [Google Scholar] [CrossRef]
- Chaklader, M.R.; Siddik, M.A.B.; Hanif, M.A.; Nahar, A.; Sultan, M.; Piria, M. Morphometric and meristic variation of endangered Pabda catfish, Ompok pabda (Hamilton-Buchanan, 1822) from southern coastal waters of Bangladesh. Pak. J. Zool. 2016, 48, 681–687. [Google Scholar]
- Yılmaz, S.; Yazıcıoğlu, O.; Erbaşaran, M.; Esen, S.; Zengin, M.; Polat, N. Length-weight relationship and relative condition factor of white bream, Blicca bjoerkna (L., 1758), from Lake Ladik, Turkey. J. Black Sea Medit. Environ. 2012, 18, 380–387. [Google Scholar]
- Barriga, R. Peces de los Afluentes de la Costa de Ecuador. In Cuencas Pericontinentales de Colombia, Ecuador, Perú y Venezuela; Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia; Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt: Bogotá, Colombia, 2015. [Google Scholar]
- Rodríguez, J.; González, A.; Angón, E.; Vivas, R.; Barba, C.; González, M.A.; Peña, F.; García, A. Efecto del tamaño de las reproductoras en la producción de alevines de Cichlasoma festae en condiciones semicontroladas en Ecuador. ITEA 2020, 116, 93–105. [Google Scholar] [CrossRef]
- Jiménez-Prado, P.; Aguirre, W.; Laaz-Moncayo, E.; Navarrete-Amaya, R.; Nugra-Salazar, F.; Rebolledo-Monsalve, E.; Zárate-Hugo, E.; Torres-Noboa, A.; Valdiviezo-Rivera, J. Guía de Peces Para Aguas Continentales en la Vertiente Occidental del Ecuador; Pontificia Universidad Católica del Ecuador Sede Esmeraldas (PUCESE); Universidad del Azuay (UDA) y Museo Ecuatoriano de Ciencias Naturales (MECN) del Instituto Nacional de Biodiversidad: Esmeraldas, Ecuador, 2015; p. 416. [Google Scholar]
- Barnhill, B.; Lopez, E.; Les, A. Estudio Sobre Biología de los Peces del Río Vinces; Instituto Nacional de Pesca de Ecuador, Boletín Científico Técnico, 1974; Volume 3. Available online: http://institutopesca.gob.ec/wp-content/uploads/2017/07/BCT.-VOL.-3-1.pdf (accessed on 14 June 2021).
- Revelo, W.; Laaz, E. Catálogo de peces de aguas continentales de la provincia de los Ríos-Ecuador. Bol. Espec. 2012, 3, 1–56. [Google Scholar]
- Valdiviezo-Rivera, J.; Garzón-Santomaro, C.; Inclán-Luna, D.; Mena_Jaén, J.; González-Romero, D. Ecosistemas Dulceacuícolas de la Provincia de El Oro: Peces y Macroinvertebrados Acuáticos como Indicadores Biológicos del Páramo al Manglar; Publicación Miscelánea N° 10: Serie de Publicaciones GADPEO-INABIO: Quito, Ecuador, 2018. [Google Scholar]
- Ortega, H.; Hidalgo, M.; Correa, E.; Espino, J.; Chocano, L.; Trevejo, G.; Meza Cortijo, A.M.; Quispe, R. Lista Anotada de los Peces de Aguas Continentales del Perú: Estado Actual del Conocimiento, Distribución, Usos y Aspectos de Conservación; Ministerio del Ambiente, Dirección General de Diversidad Biológica-Museo de Historia Natural, UNMSM: Lima, Peru, 2011; p. 48. [Google Scholar]
- Triviño, J.L. Características Morfométricas, Merísticas, Físicas y Químicas Del Pescado Ratón Silvestre (Leporinus Ecuadorensis) en la Zona de Babahoyo-2017; Proyecto de Investigación; Universidad Técnica Estatal de Quevedo: Quevedo, Ecuador, 2017. [Google Scholar]
- Revelo, W. Aspectos biológicos y pesqueros de las principales especies de peces en el sistema hídrico de la provincia de los Ríos, durante 2009. Bol. Cient. Tec. 2010, 20, 53–84. Available online: https://aquadocs.org/bitstream/handle/1834/4790/2.%20PUBLICACION%202009%20AGUA%20DULCE.pdf?sequence=1 (accessed on 14 June 2021).
- Ferrito, V.; Mannino, M.C.; Pappalardo, A.M.; Tigano, C. Morphological variation among populations of Aphanius fasciatus Nardo, 1827 (Teleostei, Cyprinodontidae) from the Mediterranean. J. Fish Biol. 2007, 70, 1–20. [Google Scholar] [CrossRef]
- Mir, F.A.; Mir, J.I.; Chandra, S. Phenotypic variation in the Snowtrout Schizothorax richardsonii (Gray, 1832) (Actinopterygii: Cypriniformes: Cyprinidae) from the Indian Himalayas. Contrib. Zool. 2013, 82, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, S.; Moqbul Hossain, M.D.; Huq, M.; Wheeler, D. Facing the hungry tide: Climate change, livelihood threats, and household responses in coastal Bangladesh. Clim. Change Econ. 2016, 7, 1650007. [Google Scholar] [CrossRef]
- Hossain, M.; Nahiduzzaman, M.; Saha, D.; Khanam, M.; Alam, M. Landmark-based morphometric and meristic variations of the endangered carp, Kalibaus labeocalbasu, from stocks of two isolated Rivers, the Jamuna and Halda and a hatchery. Zool. Stud. 2010, 49, 556–563. [Google Scholar]
- García, S.M.; Kolding, J.; Rice, J.; Rochet, M.J.; Zhou, S.; Arimoto, T.; Beyer, J.E.; Borges, L.; Bundy, A.; Dunn, D.; et al. Reconsidering the consequences of selective fisheries. Science 2012, 335, 1045–1047. [Google Scholar] [CrossRef] [Green Version]
- Ujjania, N.C.; Kohli, M.P.S. Landmark-based morphometric analysis for selected species of indian major carp (catla catla.; ham. 1822). Int. J. Food Agric. Vet. Sci. 2011, 1, 64–74. [Google Scholar]
- Quiceno-Cuartas, P.; Palacio-Baena, J.; Escobar-Cardona, J.; Werding, B. Parámetros de crecimiento individual de Petrolisthes caribensis (Werding, 1983) y Petrolisthes galathinus (Bosc, 1902) (Decapoda: Porcellanidae) en el golfo de Morrosquillo, Caribe colombiano. Actual. Biológicas 2014, 36, 163–171. [Google Scholar]
- Torres, B.; Vasseur, L.; López, R.; Lozano, P.; García, Y.; Arteaga, Y.; Bravo, C.; Barba, C.; Garcia, A. Structure and above ground biomass along an elevation small-scale gradient: Case study in an Evergreen Andean Amazon forest, Ecuador. Agrofor. Syst. 2020, 94, 1235–1245. [Google Scholar] [CrossRef]
- Gonzáles, A.; Acosta, J.; Andrade, S. Evaluación de las inundaciones de la cuenca baja del Guayas, datos y manejo. CLIRSEN. In Proceedings of the XI Congreso Ecuatoriano de la Ciencia del Suelo, Quito, Ecuador, 29–31 October 2008. [Google Scholar]
- Gobierno Provincial de Manabí. Informe Ambiental del Area Provincial de Recreación Ecológica Cascadas El Armadillo—El Pintado; Gobierno Provincial de Manabí: Portoviejo, Ecuador, 2018. [Google Scholar]
- Robin, R.R. Caracterización de la Calidad del agua para Consumo Doméstico del río Quevedo en el Cantón Quevedo, Provincia de los Ríos; BSc, University of Guayaquil: Guayaquil, Ecuador, 2015. [Google Scholar]
- Loor Castillo, E.P. Calidad del agua para la producción piscícola en el proceso de captación en la Comunidad de Pajarito del Cantón Mocache; Research Proyect, Statal University of Quevedo: Quevedo, Ecuador, 2018. [Google Scholar]
- Pacheco Bedoya, J.L. Aspectos Biológicos y Pesqueros de las Principales Especies Capturadas en el Río Babahoyo y Afluentes en el Cantón Samborondón de la Provincia del Guayas. Available online: http://www.institutopesca.gob.ec/wp-content/uploads/2018/01/Aspectos.-Biol%C3%B3gicos-y-Pesqueros-R%C3%ADo-Babahoyo-y-Afluentes-Cant%C3%B3n-Samborond%C3%B3n-2015-2.pdf (accessed on 11 May 2021).
- FAO. Perfiles Sobre la Pesca y la Acuicultura por Países La Republica de Ecuador; FAO; Fisheries and Aquaculture Department: Rome, Italy, 2013. [Google Scholar]
- MAGAP. Aspectos Pesqueros de las Principales Especies Capturadas en el Embalse Parque Lago Chongón; MAGAP, Instituto Nacional de Pesca: Guayaquil, Ecuador, 2008–2013.
- Ochoa Ubilla, B.Y.; Mendoza Nieto, K.X.; Vivas Moreira, R.; Zambrano, J.U.; Ferrer-Sánchez, Y. Ecuador Structure of catch sizes and length-weight ratio of native fish in the Abras de Mantequilla wetland, Ecuador. Cient. Tecnol. UTEQ 2016, 9, 19–27. (In Spanish) [Google Scholar]
- Diodatti, F.C.; Fonseca de Freitas, R.T.; Freato, T.A.; Pérez Ribeiro, P.A.; Solis Murgas, L.D. Parámetros morfométricos en el rendimiento de los componentes corporales de tilapia del Nilo (Oreochromis niloticus). An. Vet. Murcia 2008, 24, 45–55. [Google Scholar]
- González, M.A.; Rodríguez, J.M.; Angón, E.; Martínez, A.; García, A.; Peña, F. Characterization of morphological and meristic traits and their variations between two different populations (wild and cultured) of Cichlasoma festae, a species native to tropical Ecuadorian rivers. Arch. Anim. Breed. 2016, 59, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Caez, J.; González, A.; González, M.A.; Angón, E.; Rodríguez, J.M.; Peña, F.; Barca, C.; García, A. Application of multifactorial discriminant analysis in the morphostructural differentiation of wild and cultured populations of Vieja Azul (Andinoacara rivulatus). Turk. J. Zool. 2019, 43, 516–530. [Google Scholar] [CrossRef]
- Nash, R.R.M.; Valencia, A.H.; Geffen, A.J. The origin of Fulton’s condition factor-setting the record straight. Fisheries 2006, 31, 236–238. [Google Scholar]
- Elliott, N.G.; Haskard, K.; Koslow, J.A. Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of southern Australia. J. Fish Biol. 1995, 46, 202–220. [Google Scholar] [CrossRef]
- Revelo, W.; Castro, R. Aspectos Biológicos y Pesqueros de las Principales Especies de Peces en el Sistema Hídrico de la Provincia de los Ríos, Durante 2010; Instituto Nacional de Pesca: Guayaquil, Ecuador, 2010; p. 28. [Google Scholar]
- Choi, J.-Y.; Kim, S.-K. Effects of Aquatic Macrophytes on Spatial Distribution and Feeding Habits of Exotic Fish Species Lepomis macrochirus and Micropterus salmoides in Shallow Reservoirs in South Korea. Sustainability 2020, 12, 1447. [Google Scholar] [CrossRef] [Green Version]
- Prado, M.; Bucheli, R.; Calderón, G. Composición, distribución y abundancia del plancton en sistemas fluviales de la provincia de Los Ríos-Ecuador. Bol. Cient. Tec. 2010, 20, 1–51. [Google Scholar]
- Zhang, H.; Kang, M.; Wu, J.; Wang, C.; Li, J.; Du, H.; Yang, H.; Wei, O.V. Increasing River Temperature Shifts Impact the Yangtze Ecosystem: Evidence from the Endangered Chinese Sturgeon. Animals 2019, 9, 583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barriga, R. Lista de peces de agua dulce e intermareales del Ecuador. Rev. Politec. 2012, 30, 83–119. [Google Scholar]
- Canonico, G.C.; Arthington, A.; McCrary, J.K.; Thieme, M.L. The effects of introduced tilapias on native biodiversity. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 463–483. [Google Scholar] [CrossRef]
- Aguirre, W.E.; Navarrete, R.; Malato, G.; Calle, P.; Loh, M.K.; Vital, W.F.; Granda, J.C. Body shape variation and population genetic structure of Rhoadsia altipinna (Characidae: Rhoadsiinae) in southwestern Ecuador. Copeia 2016, 104, 554–569. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta-analysis and recommendations. J. Appl. Ichth. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Trudel, M.; Tucker, S.; Morris, J.; Higgs, D.; Welch, D. Indicators of energetic status in juvenile coho and chinook salmon. N. Am. J. Fish. Manag. 2005, 25, 374–390. [Google Scholar] [CrossRef]
- Rennie, M.D.; Verdon, R. Evaluation of condition indices for the lake whitefish, Coregonus clupeaformis. N. Am. J. Fish. Manag. 2008, 28, 1270–1293. [Google Scholar] [CrossRef]
- Youson, J.H.; Holmes, J.A.; Guchardi, J.; Seelye, J.G.; Beaver, R.E.; Gersmehl, J.E.; Sower, S.A.; Beamish, F.W.H. The importance of condition factor and the influence of water temperature and photoperiod in metamorphosis of sea lampreys. Petromyzon marinus. Can. J. Fish Aquat. Sci. 1993, 50, 2448–2456. [Google Scholar] [CrossRef]
- Ekaete, A. Preliminary studies of the condition factors in five tropical fish species of a coastal state, Lagos Nigeria. Researcher 2013, 5, 1–5. [Google Scholar]
- Melvin, G.D.; Dadswell, M.J.; McKenzie, A. Usefulness of meristic and morphometric characters in discriminating populations of American shad (Alosa sapidissima) (Ostreichthys: Clupeidae) inhabiting a marine environment. Can. J. Fish. Aquat. Sci. 1992, 49, 266–280. [Google Scholar] [CrossRef]
- Caldecutt, W.C.; Adams, D.C. Morphometrics of trophic osteology in the three spine stickleback, Gasterosteus aculeatus. Copeia 1998, 4, 827–838. [Google Scholar] [CrossRef]
- Cavalcanti, M.J.; Rabello Monteiro, L.; Duarte Lopes, P.R. Landmark-based Morphometric Analysis in Selected Species of Serranid Fishes (Perciformes: Teleostei). Zoo. Stud. 1999, 38, 287–294. [Google Scholar]
- Olaya Carbó, P. Estado ecológico del sistema estuarino del Río Guayas, Cantón Durán, Ecuador: Simulación numérica de su dinámica fluvial y principios ecológicos para el diseño de actuaciones de restauración y/o recuperación. Master’s Thesis. Universidad de Alcalá, Universidad Complutense de Madrid, Universidad Rey Juan Carlos, Universidad Politécnica de Madrid, Madrid, Spain, 2016. [Google Scholar]
- Ansah, Y.B.; Frimpong, E.A.; Hallerman, E.M. Genetically-Improved Tilapia Strains in Africa: Potential Benefits and Negative Impacts. Sustainability 2014, 6, 3697–3721. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lendech, N.; Martínez-Falcón, A.P.; Schmitter-Soto, J.J.; Mejía-Mojica, H.; Sorani-Dalbón, V.; Cruz-Ruíz, G.I.; Mercado-Silva, N. Ichthyological Differentiation and Homogenization in the Pánuco Basin, Mexico. Diversity 2020, 12, 187. [Google Scholar] [CrossRef]
- De Sosa, E.; De Paula, F. Spatial and temporal hydrochemical variation of a third order river network in a quasi pristine coastal watershed, at southern Bahia, Brazil. Ann. Braz. Acad. Sci. 2013, 85, 1357–1370. [Google Scholar]
- Laaz Moncayo, E.; Torres Noboa, A. Lista de Peces continentales de la Cuenca del Río Guayas. 2014. Available online: https://condor.depaul.edu/waguirre/fishwestec/lista_peces_guayasv2.pdf (accessed on 9 June 2021).
- Brraich, O.S.; Akhter, S. Morhometric characters and meristic Counts of a Fish, Crossocheilus latius latius (Hamilton-Buchanan) from Ranjit Sagar Wetland, India. Int. J. Fish. Aquat. Stud. 2015, 2, 260–265. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Indicators | Pintado River 1 | Quevedo River 2 | Mocache River 3 |
|---|---|---|---|
| pH | 7.72 | 8.23 | 7.11 |
| Electric conductivity (μS/cm) | 161 | 95.3 | 265.54 |
| Temperature (°C) | 25.6 | 19.2 | 26.11 |
| Turbidity (NTU) | 12.9 | 5.4 | 6.4 |
| Total dissolved solids (mg/L) | 110 | 96 | 133.5 |
| Dissolved oxygen (OD, mg/L) | 4.02 | 7.05 | 2.12 |
| Measurement | Description | Acronym |
|---|---|---|
| Weight | Total weight including the gut and gonads | BW |
| Total length 1 | Tip of the upper jaw to the top of the caudal superior end of the caudal fin | TL 1 |
| Total length 2 | Tip of the upper jaw to the top of the caudal inferior end of the caudal fin | TL 2 |
| Standard length | Tip of the upper jaw to the tail base | SL |
| Head length | From the front of the upper lip to the posterior end of the opercula membrane | HL |
| Eye diameter | The greatest bony diameter of the orbit | ED |
| Pre-orbital length | Front of the upper lip to the cranial eye edge | Pre-OL |
| Pre-dorsal fin length | Front of the upper lip to the origin of the dorsal fin | Pre-DL |
| Pre-pectoral fin length | Front of the upper lip to the origin of the pectoral fin | Pre-PcL |
| Pre-pelvic fin length | Front of the upper lip to the origin of the pelvic fin | Pre-PvL |
| Pre-anal fin length | Front of the upper lip to the origin of the anal fin | Pre-AL |
| Dorsal fin length | From the base of the first dorsal spine to the base of the last dorsal ray | DFL |
| Dorsal fin ray length | From the base to the tip of the fifth dorsal ray | DFRL |
| Pectoral fin length | From the base to the tip of the pectoral fin | PcFL |
| Pelvic fin length | From the base to the tip of the pelvic fin | PvFL |
| Anal fin length | From the base of the first anal spine to the base of the last anal ray | AFL |
| Anal fin ray length | From the base to the tip of the last anal ray | AFRL |
| Upper jaw length | Straight line measurement between the snout tip and posterior edge of maxilla | UJL |
| Body perimeter 1 | Body perimeter at the level of the first ray of the dorsal fin | P1 |
| Body perimeter 2 | Body perimeter at the level of the first radius of the anal fin | P2 |
| Body perimeter 3 | Body perimeter at the level of the last ray of the dorsal fin | P3 |
| Body width 1 | Straight line measurement from side to side at the level of the base of the first dorsal spine | LC1 |
| Body width 2 | Straight line measurement from side to side at the level of the base of the first anal spine | LC2 |
| Body width 3 | Straight line measurement from side to side at the level of the base of the last dorsal ray | LC3 |
| Body depth 1 | Body depth at the level of the first ray of the dorsal fin | AC1 |
| Body depth 2 | Body depth at the level of the first ray of the anal fin | AC2 |
| Body depth 3 | Body depth at the level of the first radius of the caudal fin | AC3 |
| Dorsal fin rays | Number of thorns in the dorsal fin | DFR DFR |
| Pectoral fin rays | Number of thorns in the pectoral fin | PcFR |
| Pelvic fin rays | Number of thorns in the pelvic fin | PvFR |
| Anal fin rays | Number of thorns in the anal fin | AFR |
| Caudal fin rays | Number of thorns in the caudal fin | CFR |
| Parameter 1 | All | Pintado River (A) | Quevedo River (B) | Mocache River (C) | p-Value | |
|---|---|---|---|---|---|---|
| A × C | B × C | |||||
| BW | 154.47 ± 5.78 (52.79) | 172.61 ± 10.30 (41.78) | 137.95 ± 8.43 (58.92) | 165.82 ± 11.34 (51.61) | 0.663 | 0.048 |
| K | 4.32 ± 0.17 (56.82) | 5.10 ± 0.35 (48.50) | 3.53 ± 0.22 (60.81) | 4.93 ± 0.34 (51.78) | 0.744 | 0.000 |
| TL | 21.46 ± 0.23 (15.41) | 20.10 ± 0.46 (16.07) | 22.52 ± 0.30 (13.02) | 20.91 ± 0.45 (16.42) | 0.217 | 0.003 |
| SL | 15.65 ± 0.16 (14.52) | 15.20 ± 0.3 (13.62) | 16.17 ± 0.22 (13.27) | 15.2 ± 0.33 (16.35) | 0.997 | 0.012 |
| HL | 5.65 ± 0.06 (16.03) | 5.38 ± 0.11 (14.85) | 5.90 ± 0.09 (15.40) | 5.47 ± 0.12 (16.34) | 0.605 | 0.005 |
| ED | 1.29 ± 0.02 (22.14) | 1.27 ± 0.05 (25.59) | 1.32 ± 0.03 (21.34) | 1.27 ± 0.03 (20.35) | 0.942 | 0.286 |
| Pre-OL | 1.19 ± 0.02 (19.94) | 1.15 ± 0.04 (21.28) | 1.23 ± 0.02 (17.57) | 1.14 ± 0.03 (21.96) | 0.801 | 0.020 |
| Pre-DL | 12.79 ± 0.14 (15.11) | 12.28 ± 0.22 (12.45) | 13.37 ± 0.20 (14.15) | 12.29 ± 0.27 (16.81) | 0.974 | 0.001 |
| Pre-PcL | 5.87 ± 0.08 (18.06) | 5.76 ± 0.2 (24.42) | 6.05 ± 0.09 (14.42) | 5.68 ± 0.13 (16.98) | 0.711 | 0.017 |
| Pre-PvL | 11.21 ± 0.12 (15.41) | 11.02 ± 0.24 (15.11) | 10.87 ± 0.16 (14.19) | 11.93 ± 0.25 (15.72) | 0.010 | 0.000 |
| Pre-AL | 15.39 ± 0.16 (14.53) | 15.2 ± 0.32 (14.57) | 14.90 ± 0.22 (14.11) | 16.34 ± 0.29 (13.46) | 0.009 | 0.000 |
| DFL | 2.54 ± 0.04 (22.18) | 2.55 ± 0.11 (30.7.) | 2.59 ± 0.05 (18.93) | 2.45 ± 0.06 (17.69) | 0.389 | 0.076 |
| DFRL | 3.52 ± 0.05 (19.13) | 3.41 ± 0.07 (14.29) | 3.47 ± 0.07 (20.65) | 3.69 ± 0.09 (19.39) | 0.024 | 0.075 |
| PcFL | 4.35 ± 0.05 (17.25) | 4.20 ± 0.10 (16.1.) | 4.47 ± 0.07 (16.09) | 4.26 ± 0.11 (19.52) | 0.703 | 0.096 |
| PvFL | 2.94 ± 0.04 (18.16) | 2.79 ± 0.07 (16.86) | 2.98 ± 0.05 (16.30) | 3.00 ± 0.08 (21.21) | 0.065 | 0.845 |
| AFL | 5.65 ± 0.1 (24.30) | 5.06 ± 0.15 (21.27) | 5.46 ± 0.15 (26.45) | 6.44 ± 0.15 (17.23) | 0.000 | 0.000 |
| AFRL | 2.51 ± 0.04 (24.09) | 2.47 ± 0.08 (23.34) | 2.56 ± 0.07 (24.73) | 2.49 ± 0.08 (23.79) | 0.866 | 0.492 |
| UJL | 1.02 ± 0.02 (27.92) | 0.88 ± 0.02 (19.7) | 1.11 ± 0.03 (25.23) | 1.00 ± 0.04 (32.25) | 0.024 | 0.036 |
| AC1 | 5.89 ± 0.05 (12.04) | 5.71 ± 0.13 (15.48) | 5.99 ± 0.06 (9.59) | 5.88 ± 0.1 (12.27) | 0.280 | 0.292 |
| AC2 | 5.44 ± 0.05 (12.93) | 5.26 ± 0.14 (18.1) | 5.53 ± 0.05 (9.00) | 5.43 ± 0.1 (13.29) | 0.314 | 0.303 |
| AC3 | 2.37 ± 0.11 (66.14) | 3.32 ± 0.41 (87.26) | 2.05 ± 0.04 (18.96) | 2.06 ± 0.05 (19.55) | 0.002 | 0.896 |
| P1 | 13.52 ± 0.19 (19.94) | 12.35 ± 0.32 (18.17) | 13.59 ± 0.26 (18.74) | 14.41 ± 0.39 (20.48) | 0.000 | 0.073 |
| P2 | 13.99 ± 0.67 (67.41) | 15.77 ± 2.64 (117.09) | 13.52 ± 0.25 (18.11) | 13.25 ± 0.4 (22.52) | 0.313 | 0.556 |
| P3 | 5.79 ± 0.06 (15.57) | 5.29 ± 0.09 (11.32) | 6.06 ± 0.09 (14.70) | 5.77 ± 0.13 (16.68) | 0.003 | 0.067 |
| LC1 | 2.60 ± 0.03 (17.70) | 2.38 ± 0.07 (20.06) | 2.64 ± 0.04 (15.39) | 2.74 ± 0.06 (17.15) | 0.000 | 0.162 |
| LC2 | 2.67 ± 0.04 (21.5) | 2.40 ± 0.08 (24.70) | 2.76 ± 0.05 (16.74) | 2.75 ± 0.09 (23.81) | 0.005 | 0.931 |
| LC3 | 2.43 ± 0.04 (22.24) | 2.13 ± 0.08 (25.00) | 2.48 ± 0.05 (18.17) | 2.58 ± 0.08 (22.74) | 0.000 | 0.247 |
| Parameter 1 | Wilks’ Lambda | Partial Lambda | F-Remove | p-Level | Toler | 1-Toler |
|---|---|---|---|---|---|---|
| AFL | 0.52 | 0.87 | 14.00 | 0.000 | 0.62 | 0.38 |
| AC3 | 0.49 | 0.91 | 8.92 | 0.000 | 0.85 | 0.15 |
| TL | 0.49 | 0.91 | 8.74 | 0.000 | 0.74 | 0.26 |
| P3 | 0.50 | 0.90 | 9.68 | 0.000 | 0.77 | 0.23 |
| UJL | 0.48 | 0.93 | 6.55 | 0.002 | 0.89 | 0.11 |
| Pre-PvL | 0.48 | 0.94 | 5.81 | 0.004 | 0.82 | 0.18 |
| AFRL | 0.47 | 0.95 | 4.80 | 0.009 | 0.72 | 0.28 |
| Pre-DL | 0.47 | 0.96 | 4.14 | 0.017 | 0.51 | 0.49 |
| ED | 0.47 | 0.95 | 4.32 | 0.015 | 0.64 | 0.36 |
| P1 | 0.46 | 0.97 | 2.51 | 0.084 | 0.80 | 0.20 |
| P2 | 0.45 | 0.99 | 1.02 | 0.363 | 0.63 | 0.37 |
| Pre-PcL | 0.45 | 0.99 | 1.09 | 0.338 | 0.63 | 0.37 |
| LC3 | 0.47 | 0.96 | 3.94 | 0.021 | 0.19 | 0.81 |
| LC2 | 0.46 | 0.97 | 2.89 | 0.058 | 0.20 | 0.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Martinez, A.; De-Pablos-Heredero, C.; González, M.; Rodriguez, J.; Barba, C.; García, A. Morphological Variations of Wild Populations of Brycon dentex (Characidae, Teleostei) in the Guayas Hydrographic Basin (Ecuador). The Impact of Fishing Policies and Environmental Conditions. Animals 2021, 11, 1901. https://doi.org/10.3390/ani11071901
Gonzalez-Martinez A, De-Pablos-Heredero C, González M, Rodriguez J, Barba C, García A. Morphological Variations of Wild Populations of Brycon dentex (Characidae, Teleostei) in the Guayas Hydrographic Basin (Ecuador). The Impact of Fishing Policies and Environmental Conditions. Animals. 2021; 11(7):1901. https://doi.org/10.3390/ani11071901
Chicago/Turabian StyleGonzalez-Martinez, Ana, Carmen De-Pablos-Heredero, Martin González, Jorge Rodriguez, Cecilio Barba, and Antón García. 2021. "Morphological Variations of Wild Populations of Brycon dentex (Characidae, Teleostei) in the Guayas Hydrographic Basin (Ecuador). The Impact of Fishing Policies and Environmental Conditions" Animals 11, no. 7: 1901. https://doi.org/10.3390/ani11071901
APA StyleGonzalez-Martinez, A., De-Pablos-Heredero, C., González, M., Rodriguez, J., Barba, C., & García, A. (2021). Morphological Variations of Wild Populations of Brycon dentex (Characidae, Teleostei) in the Guayas Hydrographic Basin (Ecuador). The Impact of Fishing Policies and Environmental Conditions. Animals, 11(7), 1901. https://doi.org/10.3390/ani11071901