
Microbial Dynamics and In Vitro Degradation of Plant Secondary Metabolites in Hanwoo Steer Rumen Fluids



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Animals, and Diet
2.2. Fermentation Media Preparation
2.3. In Vitro Degradation of Polyphenolics Using Daisy Incubator
2.4. Detection of Phenolic and Flavonoid Degradation by Rumen Fluid by HPLC-DAD Analysis
2.5. Microbiome Study by Pyrosequencing Analysis
2.6. Statistical Analysis
3. Results
3.1. Hydroxycinnamic Acid Status in Rumen Fluids after Various Incubation Periods
3.2. Flavonol and Flavone Changes in Rumen Fluids at Different Incubation Periods
3.3. Flavonol Glycosides and Ester Derivative Changes in Rumen Fluids at Different Incubation Periods
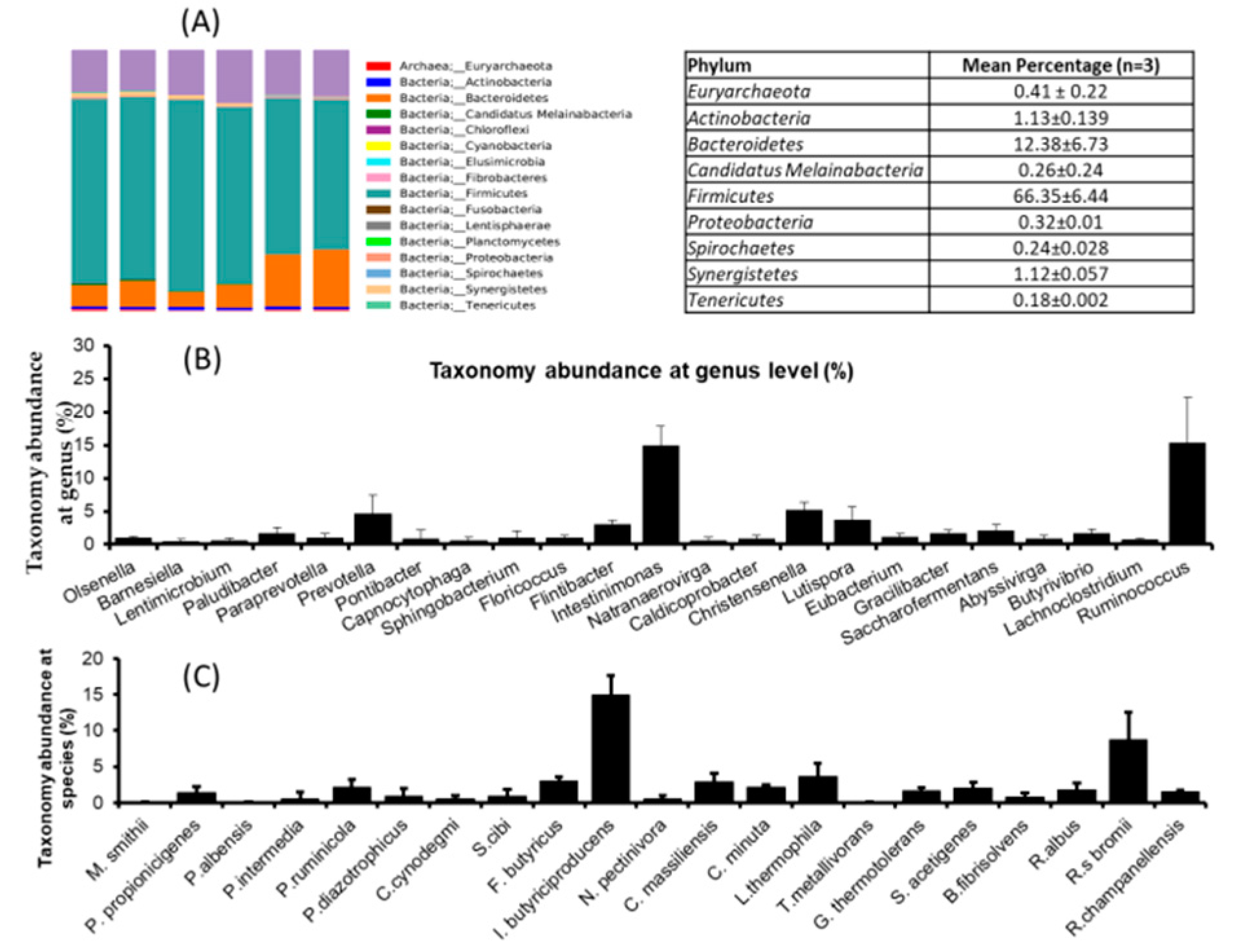
3.4. Microbial Community Dynamics of Rumen Fluid
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bjørklund, G.; Chirumbolo, S. Role of oxidative stress and antioxidants in daily nutrition and human health. Nutrition 2017, 33, 311–321. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Rumin. Res. 2003, 49, 241–256. [Google Scholar] [CrossRef]
- Silanikove, N.; Perevolotsky, A.; Provenza, F.D. Use of tannin-binding chemicals to assay for tannins and their negative postingestive effects in ruminants. Anim. Feed Sci. Technol. 2001, 91, 69–81. [Google Scholar] [CrossRef]
- Vasta, V.; Luciano, G. The effects of dietary consumption of plants secondary compounds on small ruminants’ products quality. Small Rumin. Res. 2011, 101, 150–159. [Google Scholar] [CrossRef]
- Trnková, A.; Šancová, K.; Zapletalová, M.; Kašparovská, J.; Dadáková, K.; Křížová, L.; Lochman, J.; Hadrová, S.; Ihnatová, I.; Kašparovský, T. Determination of in vitro isoflavone degradation in rumen fluid. J. Dairy Sci. 2018, 101, 5134–5144. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, M.; Schröter, D.; Esders, S.; Neugart, S.; Farquharson, F.M.; Duncan, S.H.; Schreiner, M.; Louis, P.; Maul, R.; Rohn, S. Chlorogenic acid versus amaranth’s caffeoylisocitric acid—Gut microbial degradation of caffeic acid derivatives. Food Res. Int. 2017, 100, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Trotta, R.J.; Klotz, J.L.; Harmon, D.L. Effects of source and level of dietary energy supplementation on in vitro digestibility and methane production from tall fescue-based diets. Anim. Feed Sci. Technol. 2018, 242, 41–47. [Google Scholar] [CrossRef]
- Sinz, S.; Kunz, C.; Liesegang, A.; Braun, U.; Marquardt, S.; Soliva, C.R.; Kreuzer, M. In vitro bioactivity of various pure flavonoids in ruminal fermentation, with special reference to methane formation. Czech J. Anim. Sci. 2018, 63, 293–304. [Google Scholar] [CrossRef]
- Kong, L.; Yang, C.; Dong, L.; Diao, Q.; Si, B.; Ma, J.; Tu, Y. Rumen Fermentation Characteristics in Pre- and Post-Weaning Calves upon Feeding with Mulberry Leaf Flavonoids and Candida tropicalis Individually or in Combination as a Supplement. Anim. Open Access J. 2019, 9, 990. [Google Scholar] [CrossRef]
- Aguiar, S.C.d.; Paula, E.M.d.; Yoshimura, E.H.; Santos, W.B.R.d.; Machado, E.; Valero, M.V.; Santos, G.T.d.; Zeoula, L.M. Effects of phenolic compounds in propolis on digestive and ruminal parameters in dairy cows. Rev. Bras. Zootec. 2014, 43, 197–206. [Google Scholar] [CrossRef][Green Version]
- Ungerfeld, E.M. Inhibition of Rumen Methanogenesis and Ruminant Productivity: A Meta-Analysis. Front. Vet. Sci. 2018, 5, 113. [Google Scholar] [CrossRef]
- Tong, J.; Zhang, H.; Yang, D.; Zhang, Y.; Xiong, B.; Jiang, L. Illumina sequencing analysis of the ruminal microbiota in high-yield and low-yield lactating dairy cows. PLoS ONE 2018, 13, e0198225. [Google Scholar] [CrossRef]
- Zhan, J.; Liu, M.; Wu, C.; Su, X.; Zhan, K.; Zhao, G.Q. Effects of alfalfa flavonoids extract on the microbial flora of dairy cow rumen. Asian Australas J. Anim. Sci. 2017, 30, 1261–1269. [Google Scholar] [CrossRef]
- Holden, L. Comparison of Methods of In Vitro Dry Matter Digestibility for Ten Feeds. J. Dairy Sci. 1999, 82, 1791–1794. [Google Scholar] [CrossRef]
- ANKOM Technology. A. Procedures for Fiber and In Vitro Analysis. Available online: http://www.ankom.com/homepage.html (accessed on 25 February 2020).
- Kuppusamy, P.; Lee, K.D.; Song, C.E.; Ilavenil, S.; Srigopalram, S.; Arasu, M.V.; Choi, K.C. Quantification of major phenolic and flavonoid markers in forage crop Lolium multiflorum using HPLC-DAD. Rev. Bras. Farmacogn. 2018, 28, 282–288. [Google Scholar] [CrossRef]
- Lee, H.J.; Jung, J.Y.; Oh, Y.K.; Lee, S.S.; Madsen, E.L.; Jeon, C.O. Comparative survey of rumen microbial communities and metabolites across one caprine and three bovine groups, using bar-coded pyrosequencing and ¹H nuclear magnetic resonance spectroscopy. Appl. Environ. Microbiol. 2012, 78, 5983–5993. [Google Scholar] [CrossRef]
- Oskoueian, E.; Abdullah, N.; Oskoueian, A. Effects of Flavonoids on Rumen Fermentation Activity, Methane Production, and Microbial Population. BioMed Res. Int. 2013, 2013, 349129. [Google Scholar] [CrossRef]
- Soleim, H.A.; Scheline, R.R. Metabolism of xenobiotics by strains of intestinal bacteria. Acta Pharmacol. Toxicol. 1972, 31, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Ephraim, E.; Odenyo, A.; Ashenafi, M. Screening for Tannin Degradation by Rumen and Faecal Samples of Wild and Domestic Animals in Ethiopia. World J. Microbiol. Biotechnol. 2005, 21, 803–809. [Google Scholar] [CrossRef]
- Aderao, G.N.; Sahoo, A.; Bhatt, R.S.; Kumawat, P.K.; Soni, L. In vitro rumen fermentation kinetics, metabolite production, methane and substrate degradability of polyphenol rich plant leaves and their component complete feed blocks. J. Anim. Sci. Technol. 2018, 60, 26. [Google Scholar] [CrossRef]
- Chesson, A.; Stewart, C.S.; Wallace, R.J. Influence of plant phenolic acids on growth and cellulolytic activity of rumen bacteria. Appl. Environ. Microbiol. 1982, 44, 597–603. [Google Scholar] [CrossRef]
- De Nardi, R.; Marchesini, G.; Li, S.; Khafipour, E.; Plaizier, K.J.C.; Gianesella, M.; Ricci, R.; Andrighetto, I.; Segato, S. Metagenomic analysis of rumen microbial population in dairy heifers fed a high grain diet supplemented with dicarboxylic acids or polyphenols. BMC Vet. Res. 2016, 12, 29. [Google Scholar] [CrossRef]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef]
- Murray, A.H.; Iason, G.R.; Stewart, C. Effect of simple phenolic compounds of heather (Calluna vulgaris) on rumen microbial activity in vitro. J. Chem. Ecol. 1996, 22, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.G.; Gates, D.M.; Ingledew, W.M.; Jones, G.A. Products of anaerobic phloroglucinol degradation by Coprococcus sp. Pe15. Can. J. Microbiol. 1976, 22, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Marostegan de Paula, E.; Samensari, R.B.; Machado, E.; Pereira, L.M.; Maia, F.J.; Yoshimura, E.H.; Franzolin, R.; Faciola, A.P.; Zeoula, L. Effects of phenolic compounds on ruminal protozoa population, ruminal fermentation, and digestion in water buffaloes. Livest. Sci. 2016, 185, 136–141. [Google Scholar] [CrossRef]
- Martinez-Fernandez, G.; Denman, S.E.; Cheung, J.; McSweeney, C.S. Phloroglucinol Degradation in the Rumen Promotes the Capture of Excess Hydrogen Generated from Methanogenesis Inhibition. Front. Microbiol. 2017, 8, 1871. [Google Scholar] [CrossRef]
- Berger, L.M.; Blank, R.; Zorn, F.; Wein, S.; Metges, C.C.; Wolffram, S. Ruminal degradation of quercetin and its influence on fermentation in ruminants. J. Dairy Sci. 2015, 98, 5688–5698. [Google Scholar] [CrossRef]
- Lengowski, M.B.; Zuber, K.H.R.; Witzig, M.; Möhring, J.; Boguhn, J.; Rodehutscord, M. Changes in Rumen Microbial Community Composition during Adaption to an In Vitro System and the Impact of Different Forages. PLoS ONE 2016, 11, e0150115. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Ingredients | % of DM |
|---|---|
| Rice straw | 12.5 |
| Timothy | 12.5 |
| concentrate | |
| Maize | 30.46 |
| Corn gluten feed | 15.56 |
| Wheat gluten | 10.01 |
| Soybean meal | 7.37 |
| Rapeseed meal | 3.12 |
| Coconut kernel meal | 3.11 |
| Palm kernel meal | 3.11 |
| Limestone | 1.33 |
| Salt | 0.36 |
| Sodium hydrogen carbonate | 0.36 |
| Vitamins and minerals * | 0.21 |
| Incubation Time (h) | Coumaric Acid (µg) | Ferulic Acid (µg) | Caffeic Acid (µg) |
|---|---|---|---|
| 0 | 178 ± 0.15 a | 164 ± 0.11 a | 167 ± 0.13 a |
| 12 | 142 ± 0.21 b | 142 ± 0.18 b | 172 ± 0.16 a |
| 24 | 112 ± 0.13 c | 132 ± 0.16 b | 143 ± 0.17 b |
| 48 | 90.0± 0.14 d | 110 ± 0.22 c | 109 ± 0.09 c |
| 72 | 54.0 ± 0.10 e | 88.0 ± 0.17 d | 68.0± 0.10 d |
| Incubation Time (h) | Myricetin (µg) | Luteolin (µg) | Quercetin (µg) | Kaemferol (µg) |
|---|---|---|---|---|
| 0 | 174 ± 0.23 a | 140 ±0.14 a | 174 ± 0.12 a | 188 ± 0.08 a |
| 12 | 178 ± 0.13 a | 133 ± 0.18 a | 184 ± 0.13 b | 179 ± 0.16 b |
| 24 | 123 ± 0.11 b | ND | 105 ± 0.08 c | 140 ± 0.14 c |
| 48 | 90.0 ± 0.07 c | ND | 70.0 ± 0.14 d | 102 ± 0.11 d |
| 72 | 71.0 ± 0.15 d | ND | 45.0 ± 0.12 e | 72.0 ± 0.15 e |
| Incubation Time (h) | Prophyl Gallate (µg) | Vitexin (µg) | Rutin (µg) |
|---|---|---|---|
| 0 | 168 ± 0.11 a | 173 ± 0.06 a | 176 ± 0.15 a |
| 12 | 171 ± 0.12 a | 179 ± 0.09 a | 177 ± 0.17 a |
| 24 | 164 ± 0.37 b | 119 ± 0.11 b | 106 ± 0.21 b |
| 48 | 140 ± 0.09 c | 81.0 ± 0.09 c | 78.0 ± 0.06 c |
| 72 | 110 ± 0.12 d | 66.0 ± 0.12 d | 39.0 ± 0.17 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Kuppusamy, P.; Jung, J.S.; Kim, K.H.; Choi, K.C. Microbial Dynamics and In Vitro Degradation of Plant Secondary Metabolites in Hanwoo Steer Rumen Fluids. Animals 2021, 11, 2350. https://doi.org/10.3390/ani11082350
Kim D, Kuppusamy P, Jung JS, Kim KH, Choi KC. Microbial Dynamics and In Vitro Degradation of Plant Secondary Metabolites in Hanwoo Steer Rumen Fluids. Animals. 2021; 11(8):2350. https://doi.org/10.3390/ani11082350
Chicago/Turabian StyleKim, Dahye, Palaniselvam Kuppusamy, Jeong Sung Jung, Kyoung Hoon Kim, and Ki Choon Choi. 2021. "Microbial Dynamics and In Vitro Degradation of Plant Secondary Metabolites in Hanwoo Steer Rumen Fluids" Animals 11, no. 8: 2350. https://doi.org/10.3390/ani11082350
APA StyleKim, D., Kuppusamy, P., Jung, J. S., Kim, K. H., & Choi, K. C. (2021). Microbial Dynamics and In Vitro Degradation of Plant Secondary Metabolites in Hanwoo Steer Rumen Fluids. Animals, 11(8), 2350. https://doi.org/10.3390/ani11082350