
3D Insights into the Effects of Captivity on Wolf Mastication and Their Tooth Marks; Implications in Ecological Studies of Both the Past and Present

 ,
,  , , , ,
, , , ,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. The Impact of Wolves Both Past and Present
1.2. State of the Art in Wolf Tooth Mark Analyses
2. Materials and Methods
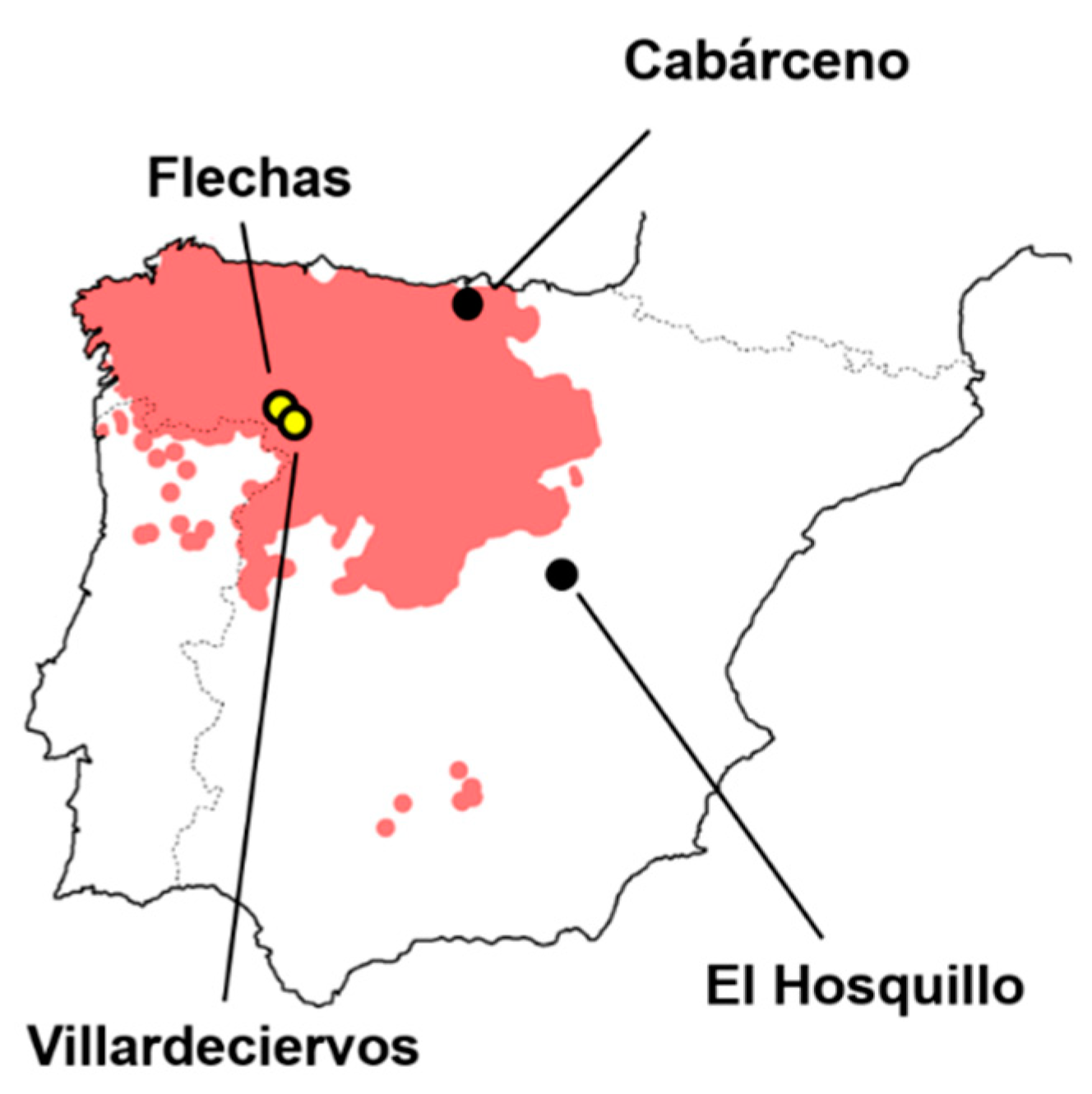
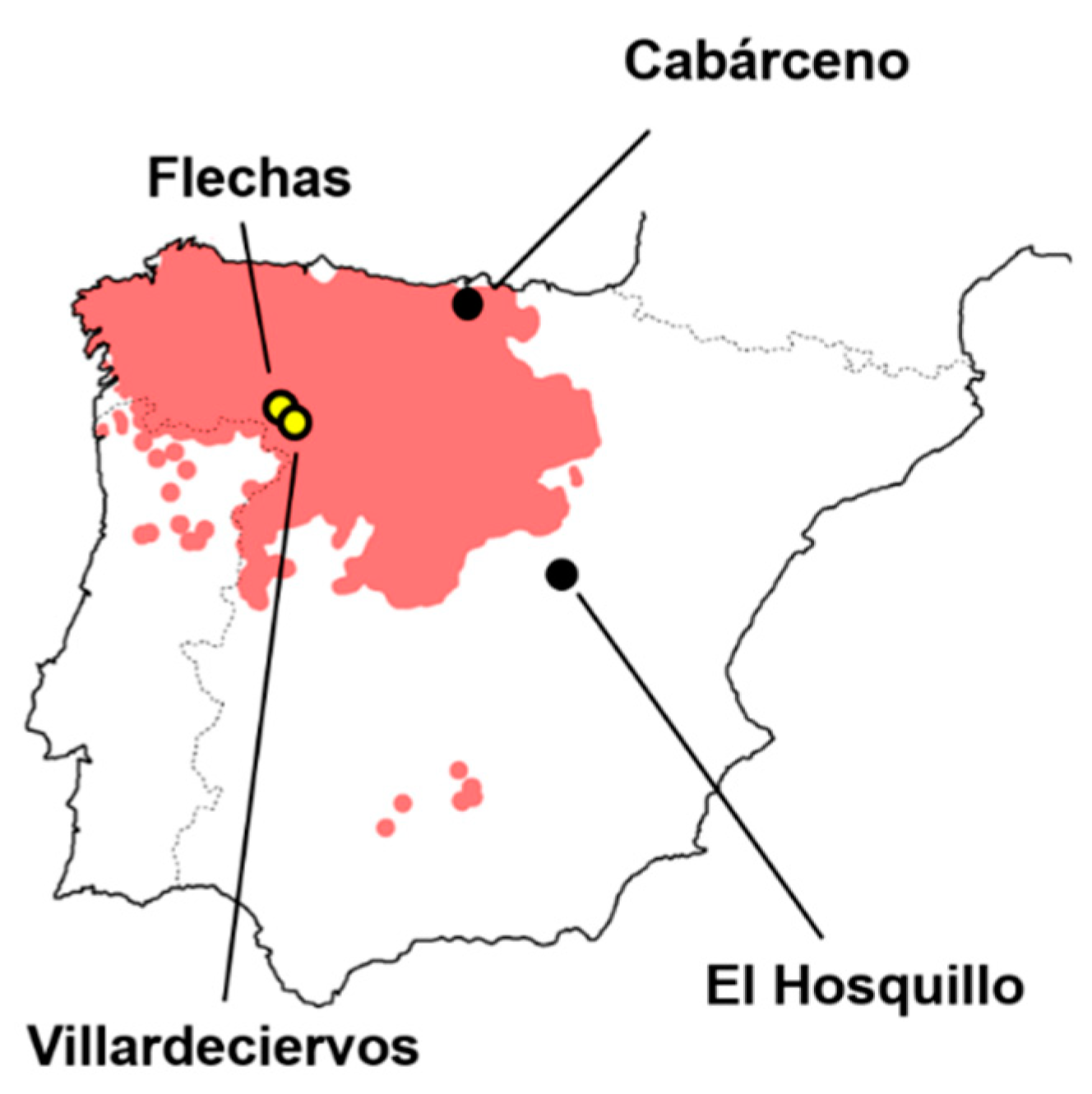
2.1. Samples
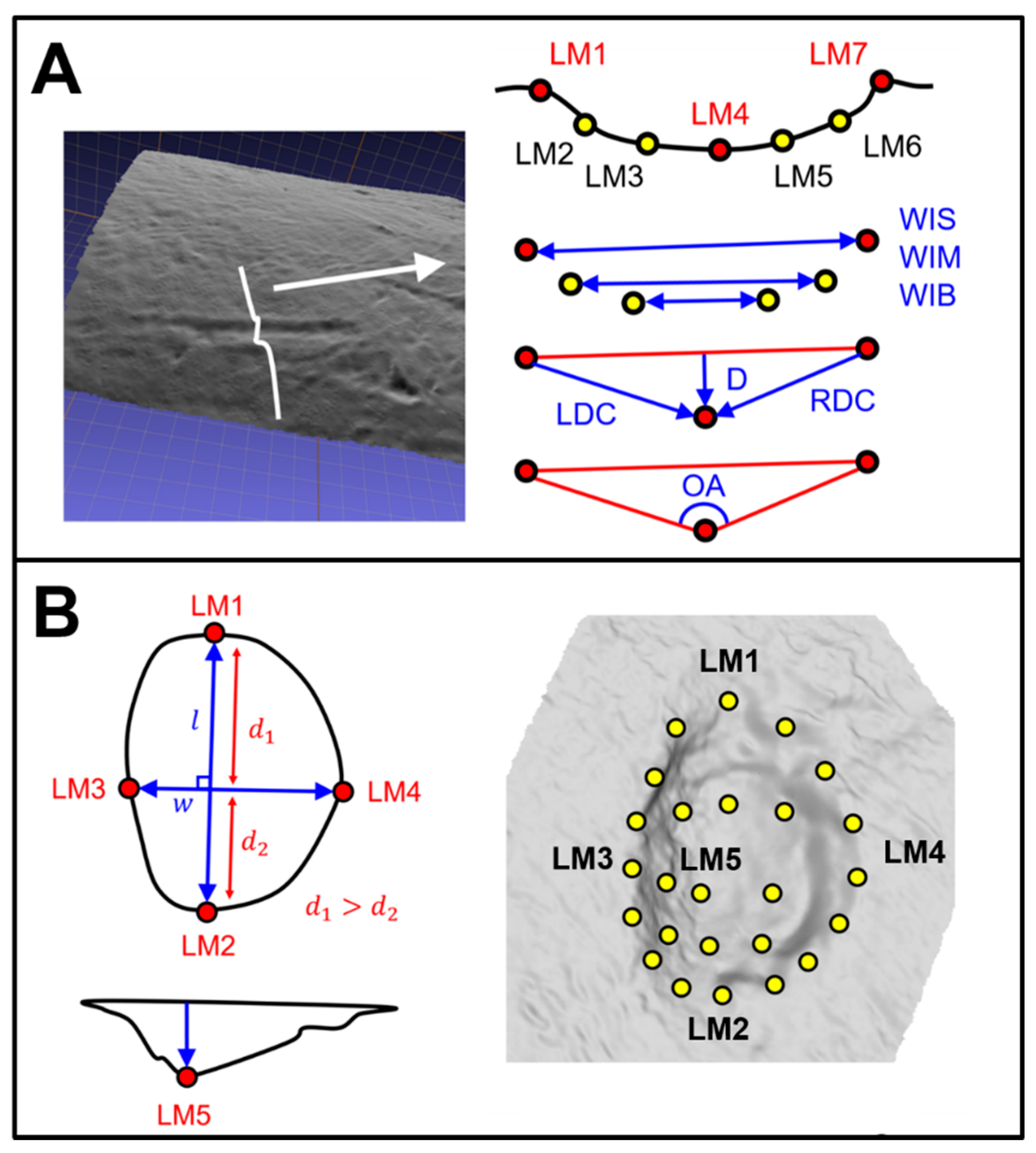
2.2. Data Collection
2.3. Statistical Analyses
2.3.1. Metric Analyses
2.3.2. Geometric Morphometric Analyses
2.3.3. Hypothesis Testing
3. Results
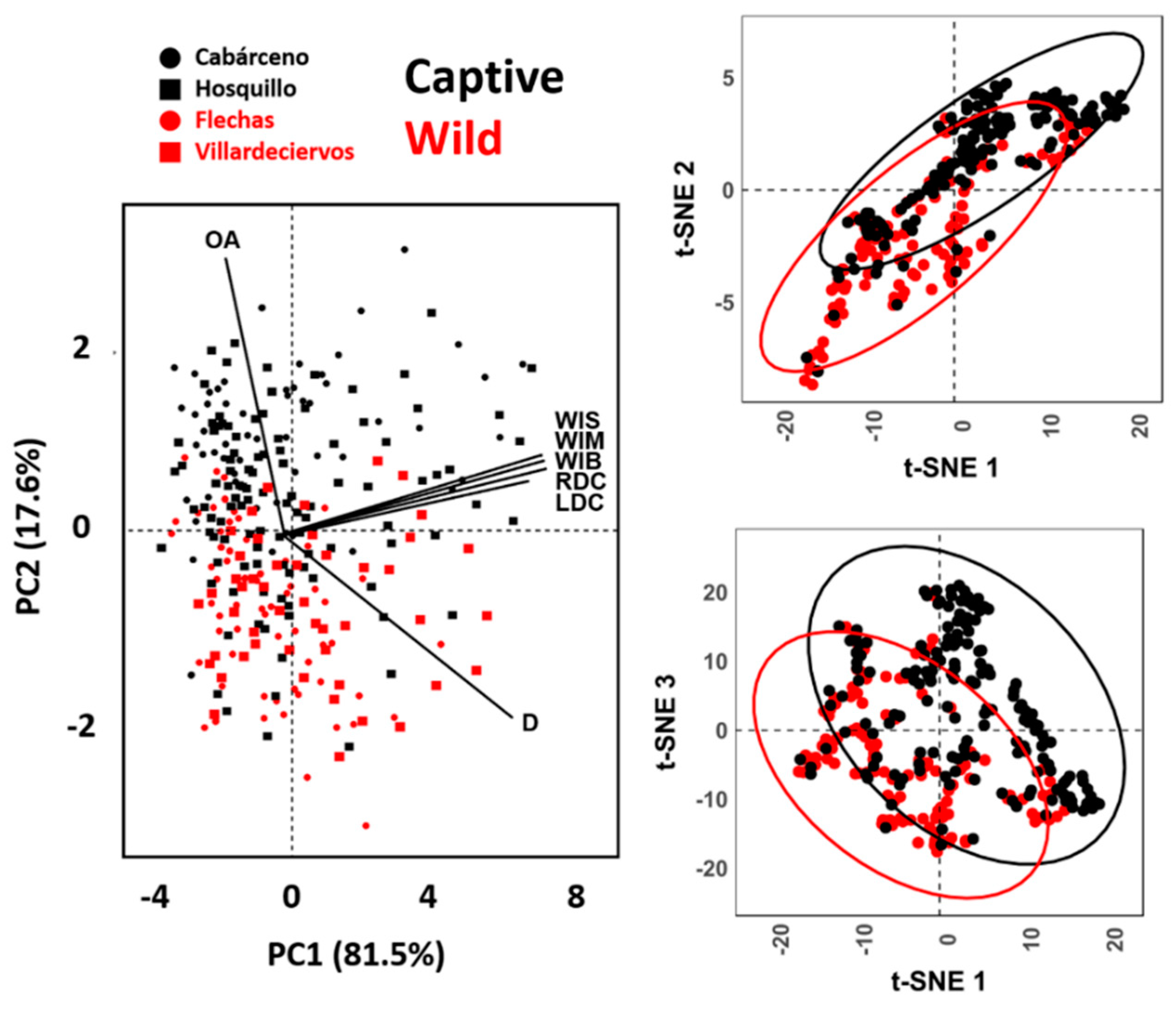
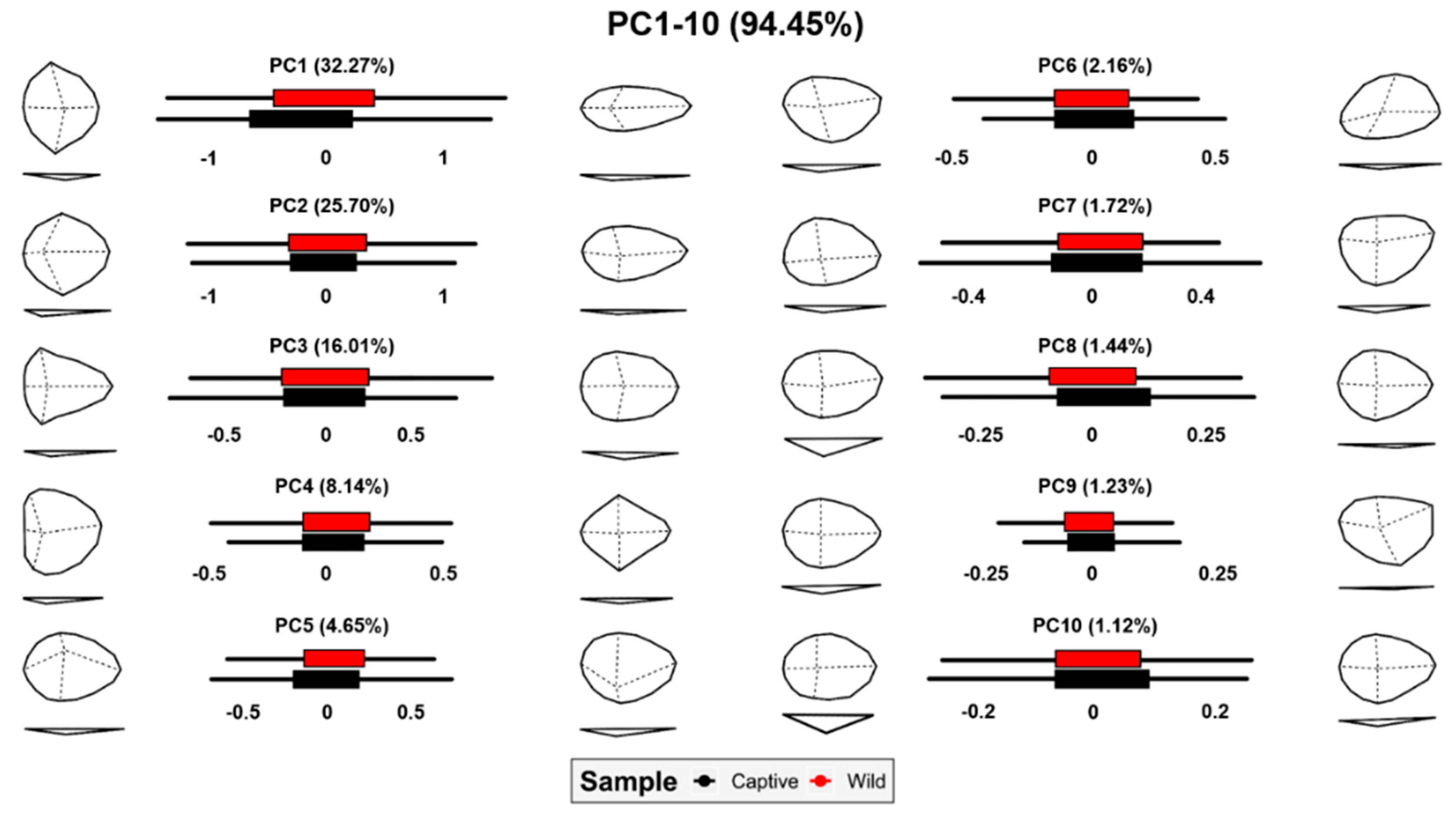
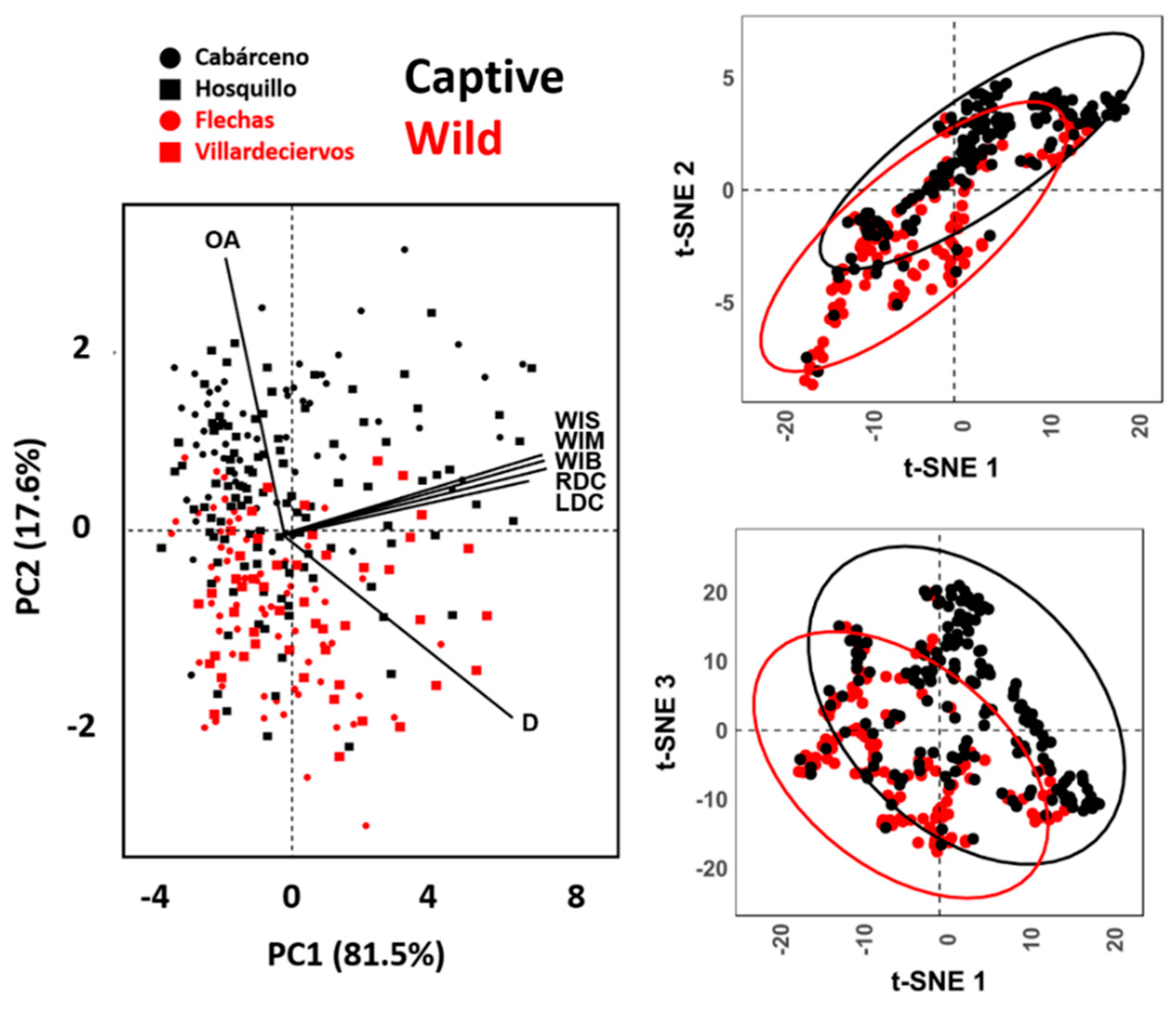
3.1. Morphometric Analyses
3.1.1. Analyses of Opening Angles (OA)
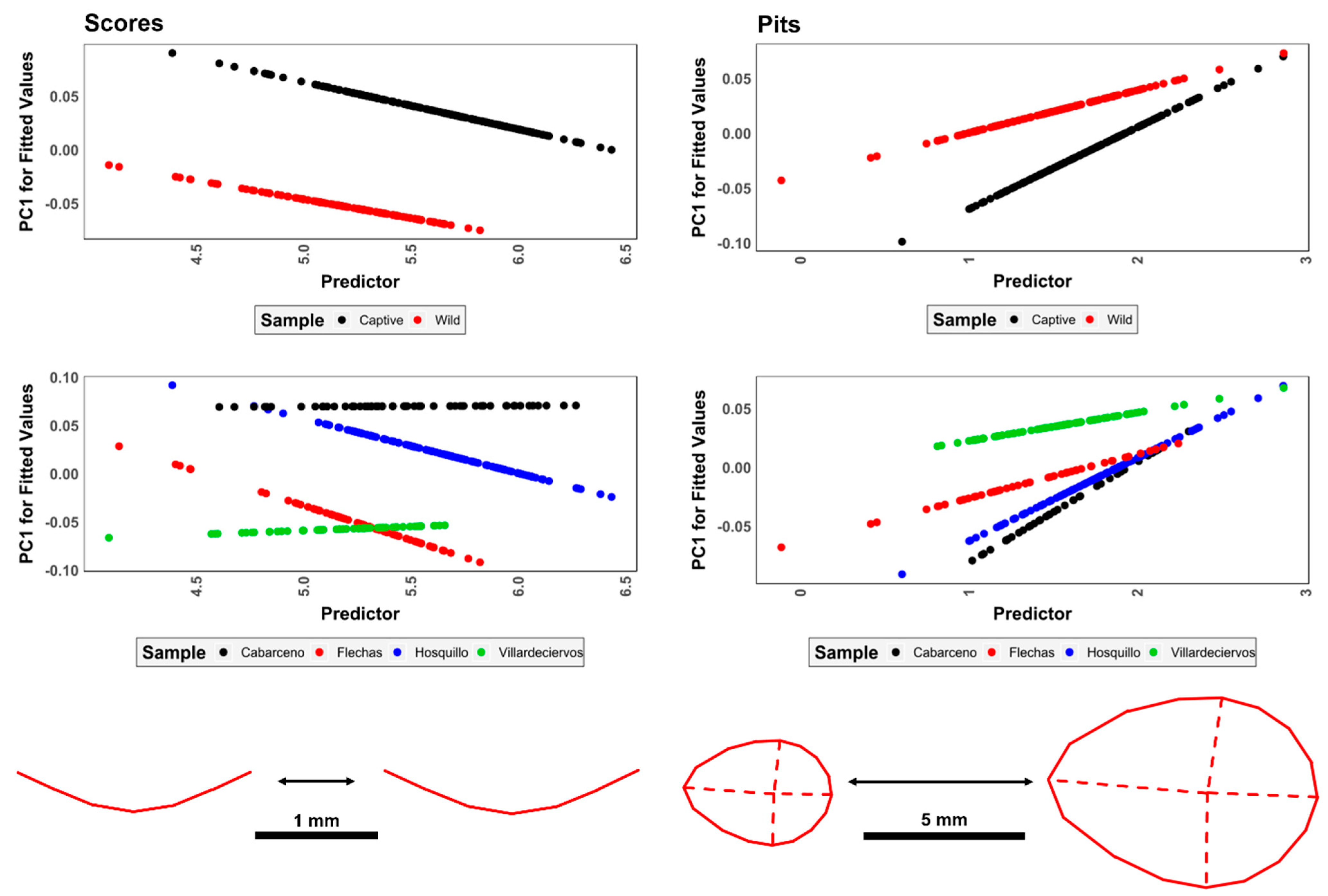
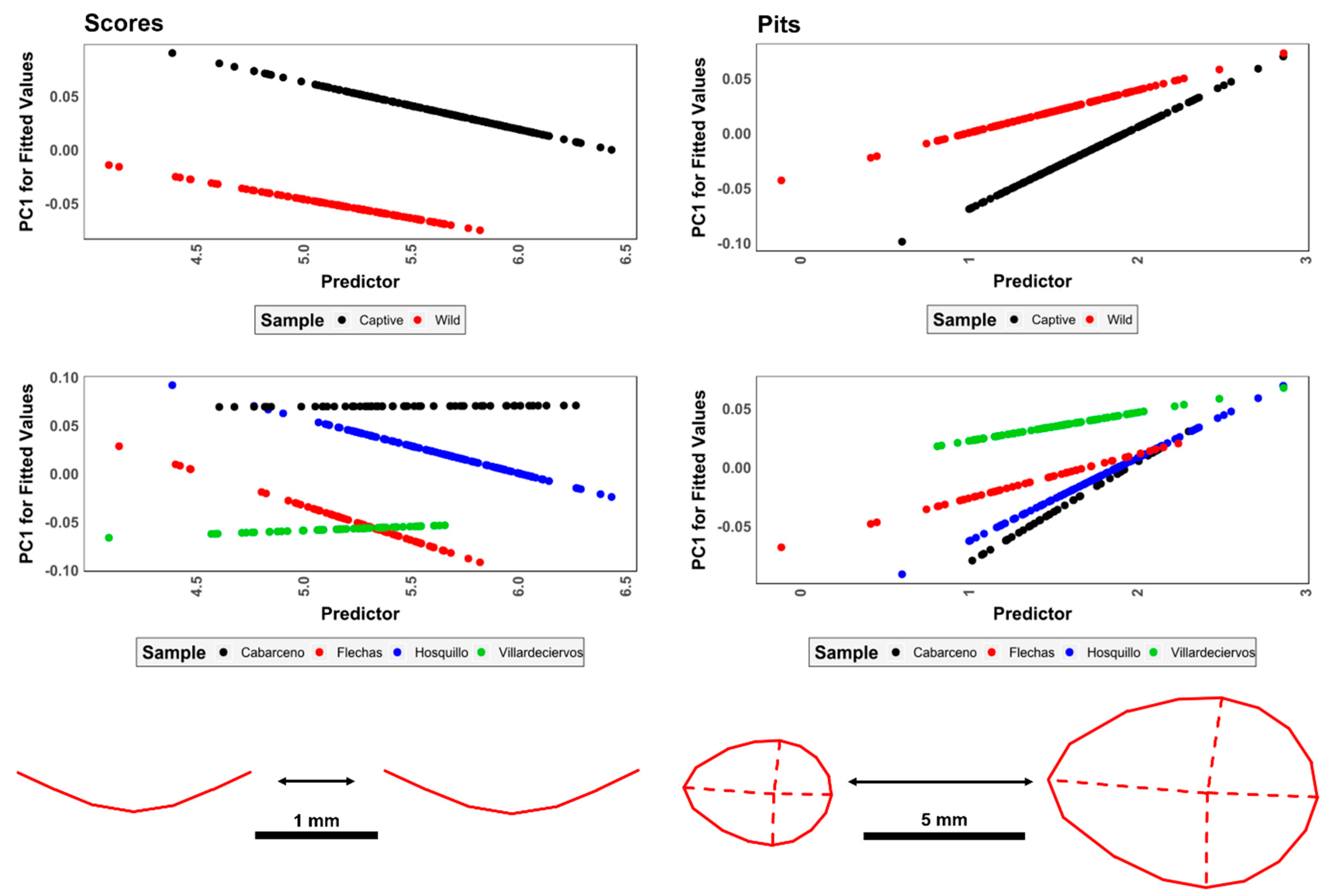
3.1.2. Analyses of Measurements
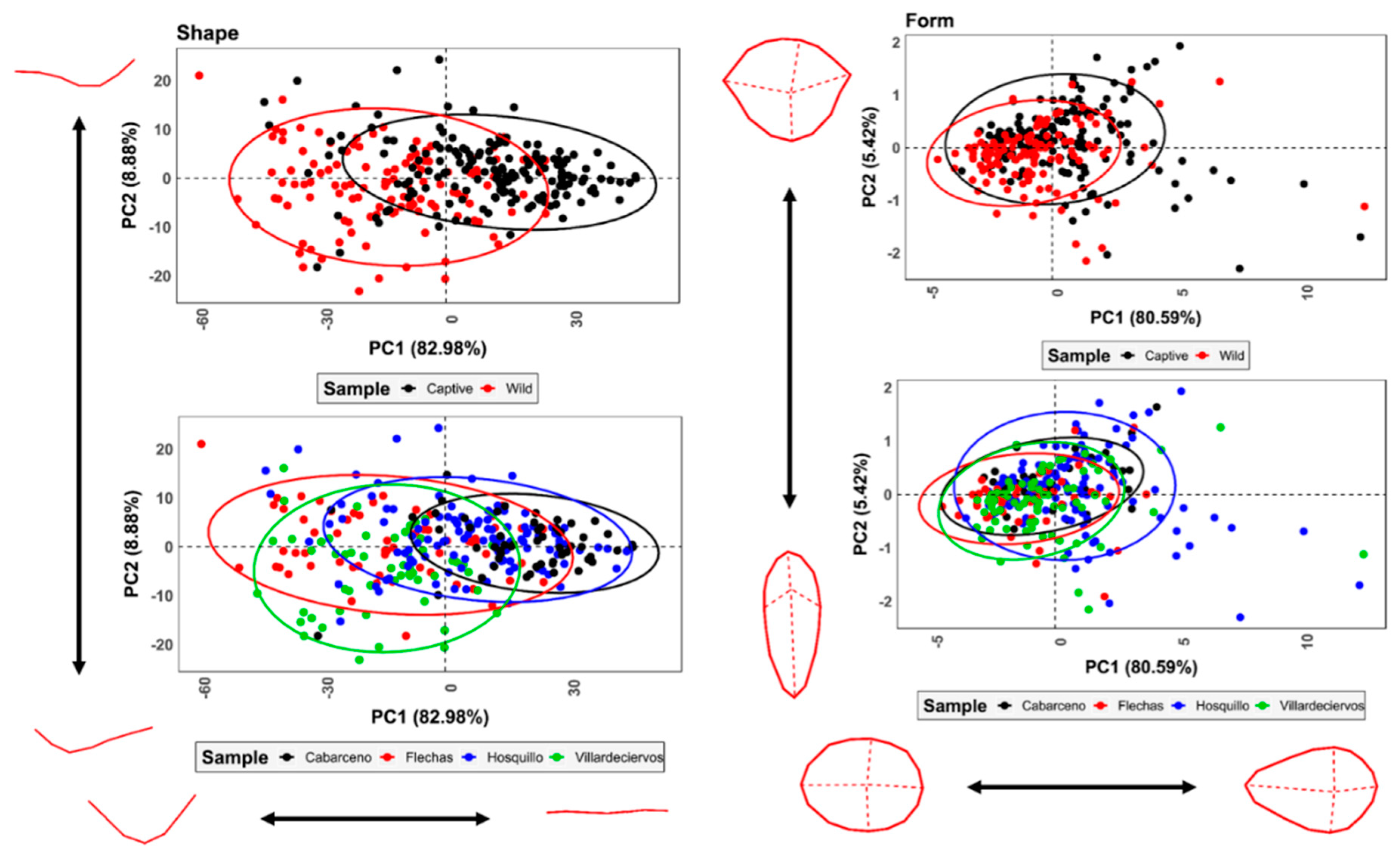
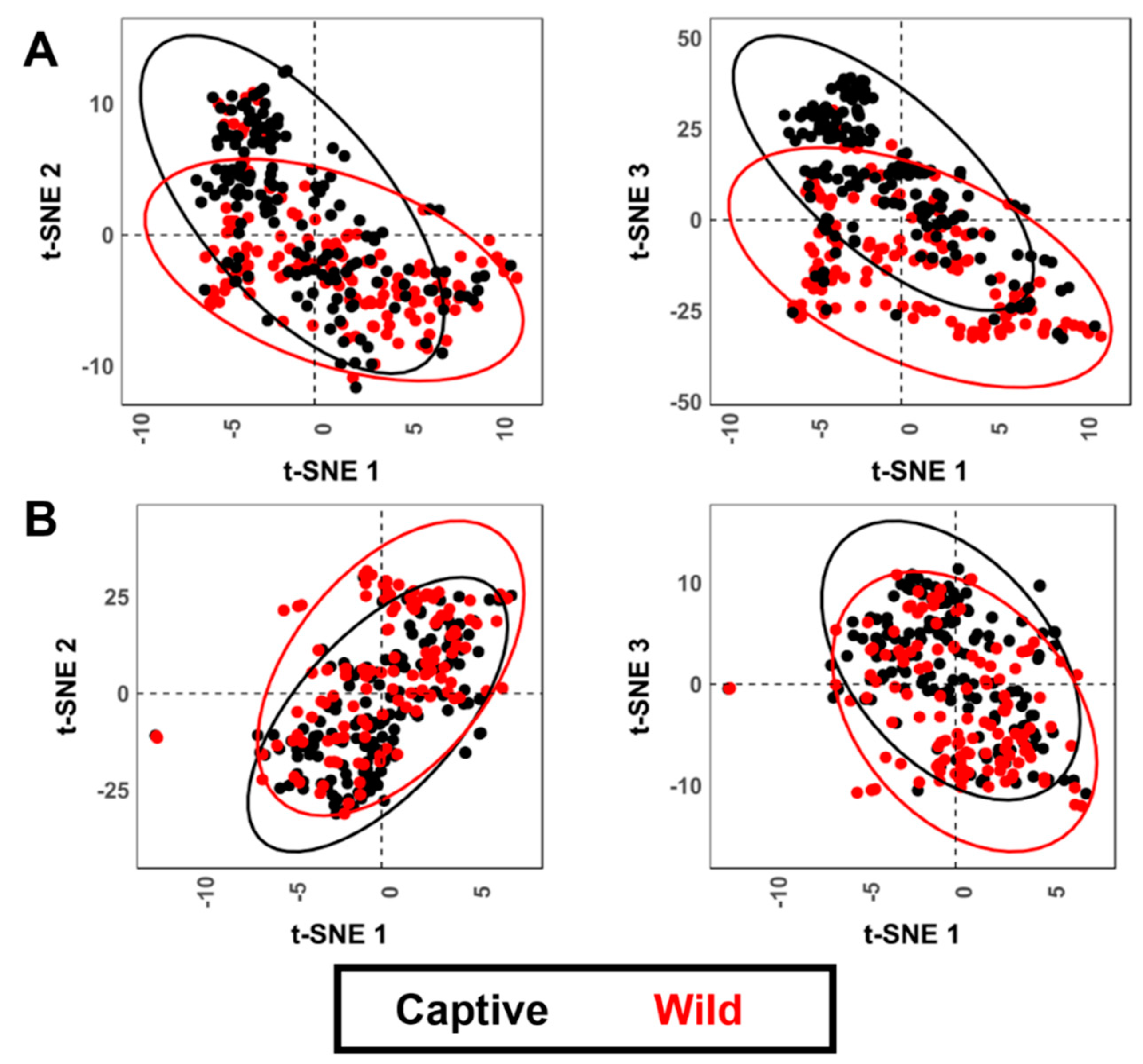
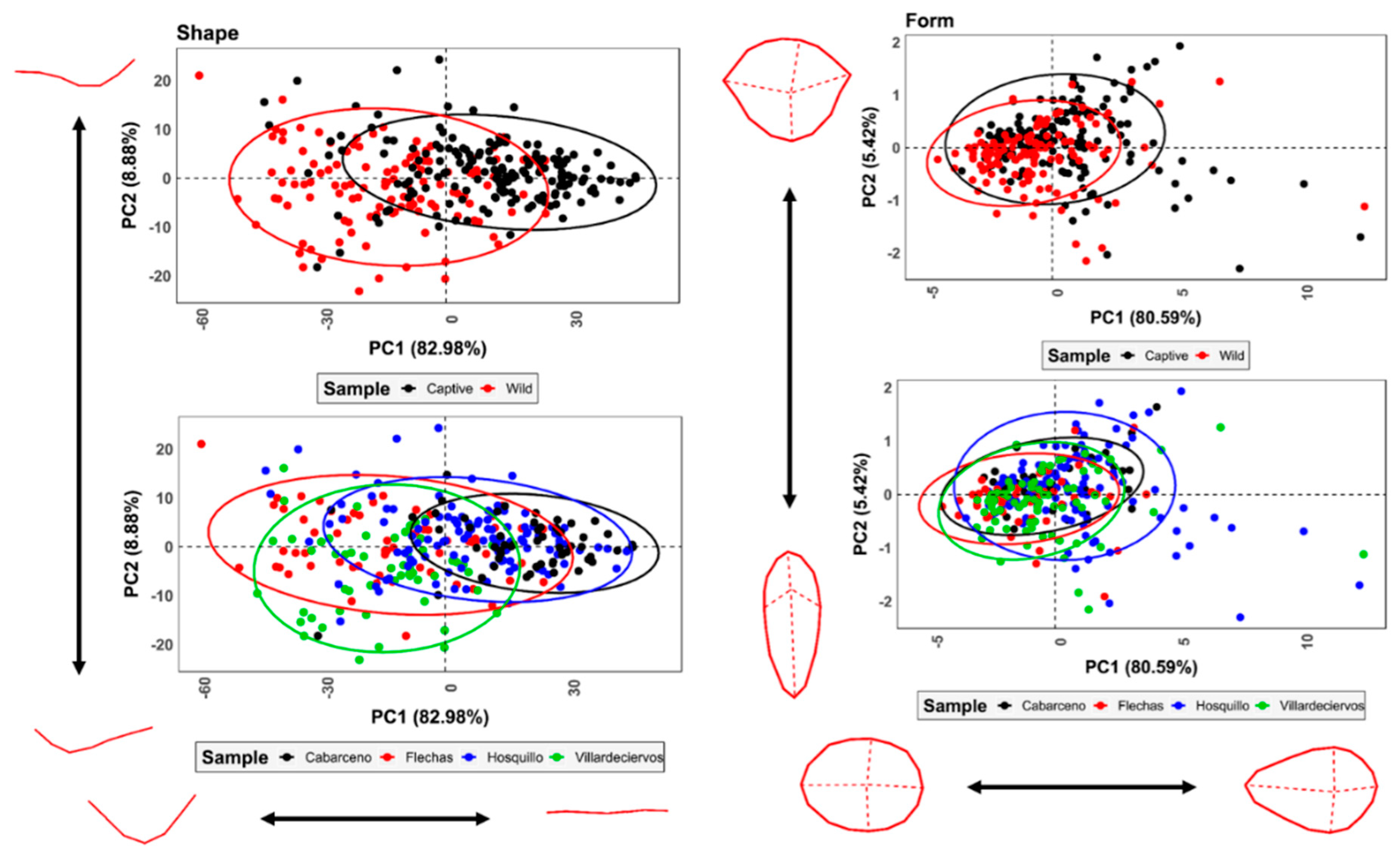
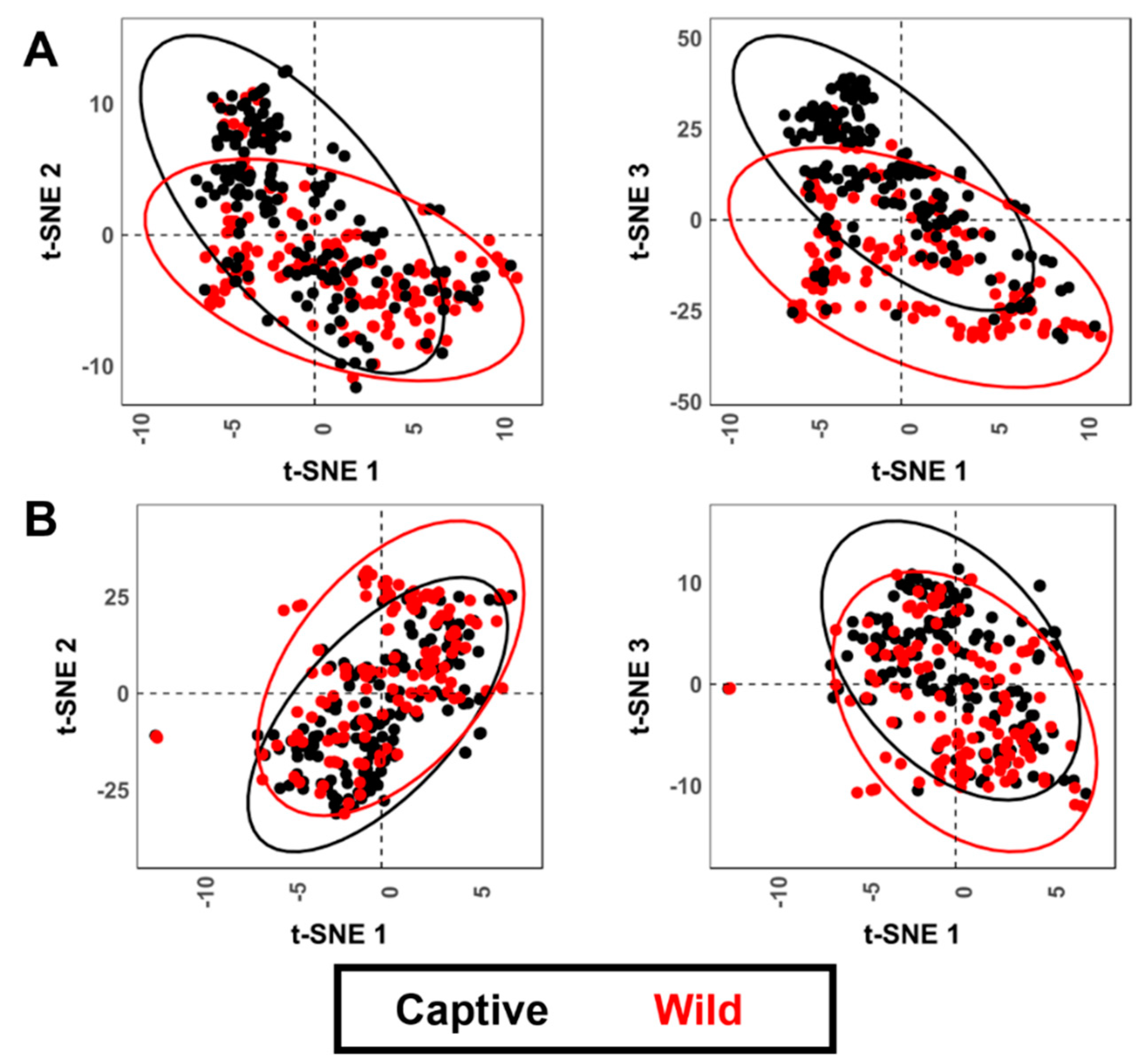
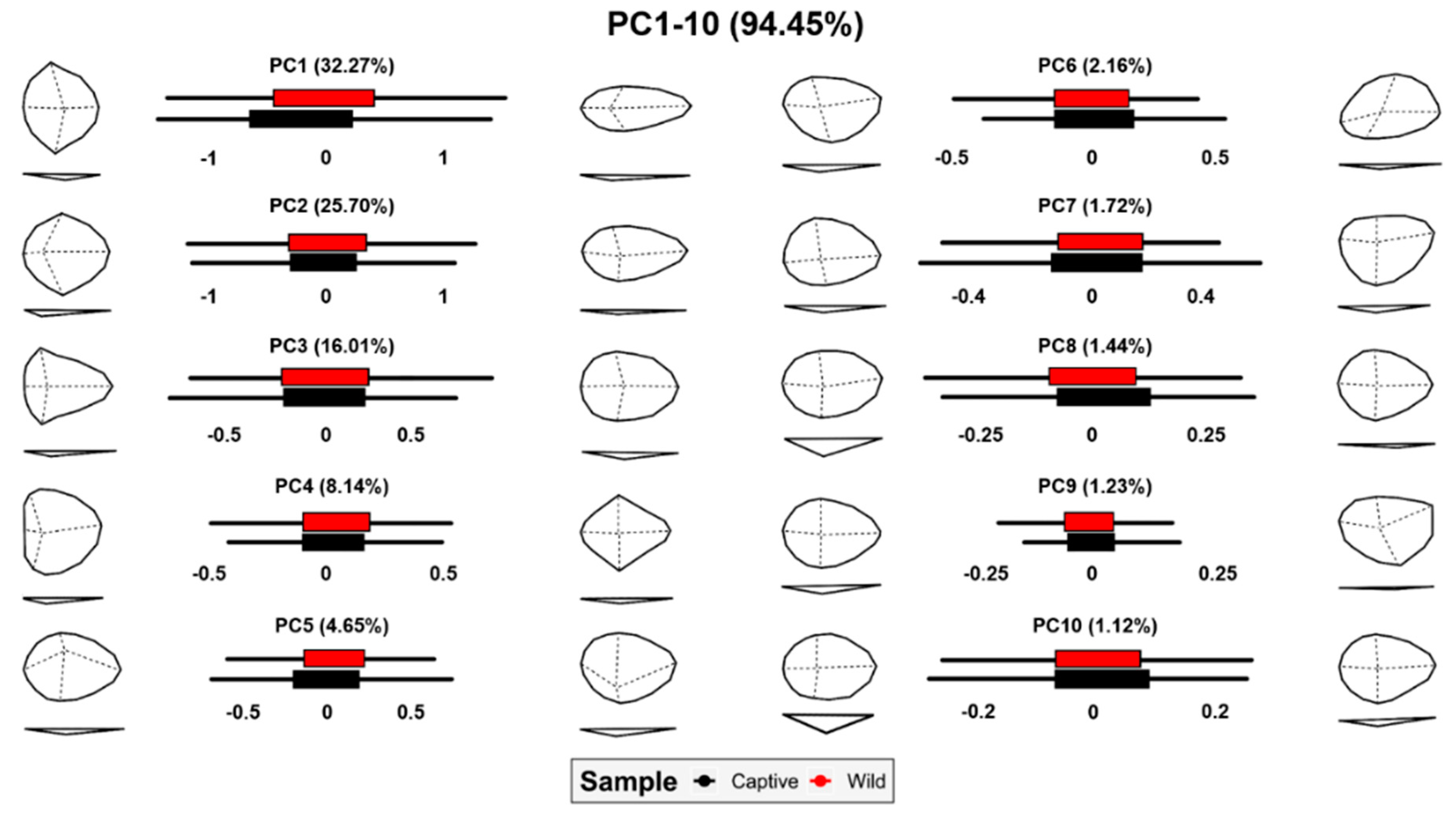
3.2. Geometric Morphometrics
3.2.1. Allometric Analyses
3.2.2. Analysis of Variance
4. Discussion
4.1. Interpretations behind Morphological Variability
4.1.1. Physiological Stress of Captivity
4.1.2. The Biomechanics of Mastication & Additional Reflections on Courtenay et al. “The Effects of Prey Size on Carnivore Tooth Mark Morphologies”
4.2. Implications for Carnivore Based Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lozano, S.; Mateos, A.; Rodríguez, J. Exploring paleo food-webs in the European Early and Middle Pleistocene: A Network Analysis. Quat. Int. 2016, 413, 44–54. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, G.; Palmqvist, P.; Ros-Montoya, S.; Espigares, M.P.; Martínez-Navarro, B. Resource availability and competition intensity in the carnivore guild of the Early Pleistocene site of Venta Micena (Orce, Baza Basin, SE Spain). Quat. Sci. Rev. 2017, 164, 154–167. [Google Scholar] [CrossRef]
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species Cum Characteribus, Differentiis. Synonymis, Locis. Tomus, I. Editio Decima, Reformata; Lautentius Salvius: Stockholm, Sweden, 1758. [Google Scholar]
- Perri, A.R.; Mitchell, K.J.; Mouton, A.; Álvarez-Carretero, S.; Hulme-Beaman, A.; Haile, J.; Jamieson, A.; Meachen, J.; Lin, A.T.; Schubert, B.W.; et al. Dire wolves were the last of an ancient New World canid lineage. Nature 2021, 591, 87–91. [Google Scholar] [CrossRef]
- Brugal, J.P.; Boudadi-Maligne, M. Quaternary small to large canids in Europe: Taxonomic status and biochronological contribution. Quat. Int. 2011, 243, 171–182. [Google Scholar] [CrossRef]
- Sardella, R.; Bertè, D.; Iurino, D.A.; Cherin, M.; Tagliacozzo, A. The wolf from Grotta Romanelli (Apulia, Italy) and its implications in the evolutionary history of Canis lupus in the Late Pleistocene of Southern Italy. Quat. Int. 2014, 328, 179–195. [Google Scholar] [CrossRef]
- Bartolini-Lucenti, S.; Bukhsianidze, M.; Martínez-Navarro, B.; Lodkipanidze, D. The Wolf from Dmanisi and Augmented Reality: Review, Implications, and Opportunities. Front. Earth Sci. 2020, 8, 131. [Google Scholar] [CrossRef]
- Crégut-Bonnoure, E. Famille des Canidae. In Les Grands Mammiféres Plio-Pléistocènes d’Europe; Guérin, C., Patou-Mathis, M., Eds.; Elsevier Masson: Paris, France, 1996; pp. 156–166. [Google Scholar]
- Ripoll, M.P.; Morales-Pérez, J.V.; Serra, A.S.; Tortosa, E.A.; Montañana, I.S. Presence of the genus Cuon in the Upper Pleistocene and initial Holocene sites of the Iberian Peninsula: New remains identified in archaeological contexts of the Mediterranean region. J. Archaeol. Sci. 2010, 37, 437–450. [Google Scholar] [CrossRef]
- Mallye, J.B.; Costamagno, S.; Boudadi-Maligne, M.; Prucca, A.; Lautoulandie, V.; Thiébaut, C.; Mourre, V. Dhole (Cuon alpinus) as a bone accumulator and new taphonomic agent? The case of Noisetier cave (French Pyrenees). J. Taphon. 2012, 10, 317–547. [Google Scholar]
- Martínez-Navarro, B.; Lucenti, S.B.; Palmqvist, P.; Ros-Montoya, S.; Madurell-Malapeira, J.; Espigares, M.P. A new species of dog from the Early Pleistocene site of Venta Micena (Orce, Baza Basin, Spain). Comptes Rendus Palevol 2021, 20, 297–314. [Google Scholar] [CrossRef]
- Coumont, M.P. Proposition d’un référentiel taphonomique fossile de faunes issues d’avens-pèges. Ann. De Paléontologie 2009, 95, 1–20. [Google Scholar] [CrossRef]
- Castel, J.C.; Coumont, M.P.; Boudadi-Maligne, M.; Prucca, A. Rôle et Origine des Grandes Carnivores dans les Accumulations Naturelles. Le Cas de Loups (Canis lupus) de l’Igue du Gral (Sauliac-sur-Célé, Lot, France). Rev. De Paléobiologie Genève 2010, 29, 411–425. [Google Scholar]
- Campmas, E.; Michel, P.; Costamagno, S.; El Hajraoui, M.A.; Nespoulet, R. Which predators are responsable for faunal accumulations at the Late Pleistocene layers of El Harhoura 2 Cave (Témara, Morocco)? Comptes Rendus Palevol 2017, 16, 333–350. [Google Scholar] [CrossRef]
- Mech, L.D. The Wolf: The Ecology and Behaviour of an Endangered Species; Natural History Press: New York, NY, USA, 1970. [Google Scholar]
- Haynes, G. Prey bones and predators: Potential ecologic information from analysis of bone sites. Ossa 1980, 7, 75–97. [Google Scholar]
- Haynes, G. Bone Modification and Skeletal Disturbances by Natural Agencies: Studies in North America. Ph.D. Thesis, The Catholic University of America, Washington, DC, USA, 1981. [Google Scholar]
- Haynes, G. Utilization and skeletal disturbances of North American Prey Carcasses. Arctic 1982, 35, 266–281. [Google Scholar] [CrossRef]
- Yravedra, J.; Lagos, L.; Bárcena, F. The Wild Wolf (Canis lupus) as a dispersal agent of animal carcasses in Northwestern Spain. J. Taphon. 2012, 10, 227–248. [Google Scholar]
- Ovodov, N.; Crockford, S.J.; Kuzmin, Y.V.; Higham, T.F.G.; Hodgins, G.W.L.; Plicht, J. A 33,000-Year-Old incipient dog from the Altai Mountains of Siberia: Evidence of the Earliest Domestication Disrupted by the Last Glacial Maximum. PLoS ONE 2011, 6, e22821. [Google Scholar] [CrossRef] [Green Version]
- Germonpré, M.; Fedorov, S.; Danilov, P.; Galeta, P.; Jimenez, E.L.; Sablin, M.; Losey, R.J. Palaeolithic and prehisotric dogs and Pleistocene wolves from Yakutia: Identification of Isolated Skulls. J. Archaeol. Sci. 2017, 78, 1–19. [Google Scholar] [CrossRef]
- Drake, A.G.; Coquerelle, M.; Colombeau, G. 3D morphometric analysis of fossil canid skulls contradicts the suggested domestication of dogs during the Late Palaeolithic. Sci. Rep. 2015, 5, 8299. [Google Scholar] [CrossRef] [Green Version]
- Wilczyński, J.; Haynes, G.; Sobczyk, Ł.; Svoboda, J.; Roblíčková, M.; Wojtal, P. Friend or Foe? Large canid remains from Pavlovian sites and their archaeozoological context. J. Anthr. Arch. 2020, 59, 101197. [Google Scholar] [CrossRef]
- Larson, G.; Karlsson, E.K.; Perri, A.; Webster, M.T.; Ho, S.Y.W.; Peters, J.; Stahl, P.W.; Piper, P.J.; Lingaas, F.; Fredholm, M.; et al. Rethinking dog domestication by integrating genetics, archaeological and biogeography. Proc. Natl. Acad. Sci. USA 2012, 109, 8878–8883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cayuela, L. Habitat evaluation for the Iberian wolf Canis lupus in Picos de Europa National Park, Spain. Appl. Geogr. 2004, 24, 199–215. [Google Scholar] [CrossRef]
- Blanco, J.C.; Cortes, Y. Ecological and social constraints of wolf recovery in Spain, A New Era for Wolves and People. In Wolf Recovery, Human Attitudes and Policy; Musiani, M., Boitani, L., Paquet, P.C., Eds.; University of Calargy Press: Calargy, AB, Canada, 2009; pp. 41–66. [Google Scholar]
- Woodroffe, R.; Redpath, S.M. When the hunter becomes the hunted. Science 2015, 348, 1312–1314. [Google Scholar] [CrossRef]
- Pimenta, V.; Barroso, I.; Boitani, L.; Beja, P. Wolf predation on cattle in Portugal: Assessing the effects of husbandry systems. Biol. Conserv. 2017, 207, 17–26. [Google Scholar] [CrossRef]
- Pimenta, V.; Barroso, I.; Boitani, L.; Beja, P. Risks a la carte: Modelling the occurrence and intensity of wolf predation on multiple livestock species. Biol. Conserv. 2018, 228, 331–342. [Google Scholar] [CrossRef]
- Ericsson, G.; Heberlein, T.A.; Karlsson, J.; Bjärvall, A.; Lundvall, A. Support for hunting as a means of wolf Canis lupus population control in Sweden. Wildlife Biol. 2004, 10, 269–276. [Google Scholar] [CrossRef]
- Lososová, J.; Kouřilová, J.; Soukupová, N. Controversial approach to wolf management in the Czech Republic. Agr. Econ. 2021, 67, 1–10. [Google Scholar] [CrossRef]
- Yravedra, J.; Maté-González, M.Á.; Courtenay, L.A.; González-Aguilera, D.; Fernández-Fernández, M. The use of canid tooth marks on bone for the identification of livestock predation. Sci. Rep. 2019, 9, 16301. [Google Scholar] [CrossRef] [Green Version]
- Dawkins, W.B. Cave Hunting, Researches of the Evidence of Caves Respecting the Early Inhabitants of Europe; Macmillian & Co.: London, UK, 1874. [Google Scholar]
- Martin, H. Reserches sur L’evolution du Mousterien dans le Gisement de la Quina Charante. In Premier Volume—Industrie Osseuse; Schleicher Freres: Paris, France, 1907–1910. [Google Scholar]
- Binford, L.R. Bones: Ancient Men and Modern Myths; Academic Press Inc.: New York, NY, USA, 1981. [Google Scholar]
- Blumenschine, R.J. Percussion marks, tooth marks and the experimental determinations of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. J. Hum. Evol. 1995, 29, 21–51. [Google Scholar] [CrossRef]
- Selvaggio, M.M.; Wilder, J. Identifying the involvement of multiple carnivore taxa with archaeological bone assemblages. J. Archaeol. Sci. 2001, 28, 465–470. [Google Scholar] [CrossRef]
- Delaney-Rivera, C.; Plummer, T.W.; Hodgson, J.A.; Forrest, F.; Hertel, F.; Oliver, J.S. Pits and pitfalls: Taxonomic variability and patterning in tooth mark dimensions. J. Archaeol. Sci. 2009, 36, 2597–2608. [Google Scholar] [CrossRef]
- Andrés, M.; Gidna, A.O.; Yravedra, J.; Domínguez-Rodrigo, M. A study of dimensional differences of tooth marks (pits and scores) on bones modified by small and large carnivores. J. Archaeol. Anthropol. Sci. 2012, 4, 209–219. [Google Scholar] [CrossRef]
- Yravedra, J.; García-Vargas, E.; Maté-González, M.Á.; Aramendi, J.; Palomeque-González, J.; Vallés-Iriso, J.; Matesanz-Vicente, J.; González-Aguilera, D.; Domínguez-Rodrigo, M. The use of Micro-Photogrammetry and Geometric Morphometrics for identifying carnivore agency in bone assemblage. J. Archaeol. Sci: Rep. 2017, 14, 106–115. [Google Scholar] [CrossRef]
- Aramendi, J.; Maté-González, M.Á.; Yravedra, J.; Cruz-Ortega, M.; Arriaza, M.C.; González-Aguilera, D.; Baquedano, E.; Domínguez-Rodrigo, M. Discerning carnivore agency through the three-dimensional study of tooth pits: Revisiting crocodile feeding at FLK-Zinj and FLK NN3 (Olduvai Gorge, Tanzania). Palaeogeog. Palaeoclim. Palaeoecol. 2017, 488, 93–102. [Google Scholar] [CrossRef]
- Arriaza, M.C.; Yravedra, J.; Domínguez-Rodrigo, M.; Maté-González, M.Á.; García-Vargas, E.; Palomeque-González, J.F.; Aramendi, J.; González-Aguilera, D.; Baquedano, E. On applications of micro-photogrammetry and geometric morphometrics to studies of tooth-mark morphology: The modern Olduvai Carnivore Site (Tanzania). Palaeogeog. Palaeoclim. Palaeoecol. 2017, 488, 103–112. [Google Scholar] [CrossRef]
- Arriaza, M.C.; Aramendi, J.; Maté-González, M.Á.; Yravedra, J.; Stratford, D. Characterising leopard as taphonomic agent through the use of micro-photogrammetric reconstruction of tooth marks and pit to score ratio. Hist. Biol. 2019, 33, 176–185. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Yravedra, J.; Huguet, R.; Aramendi, J.; Maté-González, M.Á.; González-Aguilera, D.; Arriaza, M.C. Combining machine learning algorithms and geometric morphometrics: A study of carnivore tooth marks. Palaeogeog. Palaeoclim. Palaeoecol. 2019, 522, 28–29. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Herranz-Rodrigo, D.; González-Aguilera, D.; Yravedra, J. Developments in Data Science Solutions for Carnivore Tooth Pit Classification. Sci. Rep. 2021, 11, 10209. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Herranz-Rodrigo, D.; Huguet, R.; Maté-González, M.Á.; González-Aguilera, D.; Yravedra, J. Obtaining new resolutions in carnivore tooth pit morphological analyses: A methodological update for digital taphonomy. PLoS ONE 2020, 15, e0240328. [Google Scholar] [CrossRef]
- Pante, M.C.; Muttart, M.V.; Keevil, T.L.; Blumenschine, R.J.; Njau, J.K.; Merritt, S.R. A new high-resolution 3-D quantitative method for identifying bone surface modifications with implications for the Early Stone Age archaeological record. J. Hum. Evol. 2017, 102, 1–11. [Google Scholar] [CrossRef]
- Gümrükçu, M.; Pante, M.C. Assessing the Effects of Fluvial Abrasion on Bone Surface Modifications using High-Resolution 3-D Scanning. J. Archaeol. Sci. Rep. 2018, 21, 208–221. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-García, B.; Aznarte, J.; Abellán, N.; Baquedano, E.; Domínguez-Rodrigo, M. Deep learning improves taphonomic resolution: High accuracy in differentiating tooth marks made by lions and jaguars. J. R Soc. Interface 2020, 17, 2020046. [Google Scholar] [CrossRef]
- Abellán, N.; Jiménez-García, B.; Aznarte, J.; Baquedano, E.; Domínguez-Rodrigo, M. Deep learning classification of tooth scores made by different carnivores: Achieving high accuracy when comparing African carnivore taxa and testing the hominin shift in the balance of power. Archaeol. Anthropol. Sci. 2021, 13, 31. [Google Scholar] [CrossRef]
- Gidna, A.; Yravedra, J.; Domínguez-Rodrigo, M. A cautionary note on the use of captive carnivores to model wild predator behavior: A comparison of bone modification patterns on long bones by captive and wild lions. J. Archaeol. Sci. 2013, 40, 1903–1910. [Google Scholar] [CrossRef]
- Gidna, A.; Domínguez-Rodrigo, M.; Pickering, T.R. Patterns of bovid long limb bone modification created by wild and captive leopards and their relevance to the elaboration of referential frameworks for palaeoanthropolgy. J. Archaeol. Sci. Rep. 2015, 2, 302–309. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Yravedra, J.; Maté-González, M.Á.; Vázquez-Rodríguez, J.M.; Fernández-Fernández, M.; González-Aguilera, D. The effects of prey size on carnivore tooth mark morphologies on bone; the case study of Canis lupus signatus. Hist. Biol. 2020, 1–13. [Google Scholar] [CrossRef]
- Cabrera, A. Los Lobos de España; Boletín de la Sociedad Royal Española Historia: Madrid, Spain, 1907. [Google Scholar]
- Christainsen, P.; Adolfssen, S. Bite Foreces, canine strength and skull allometry in carnivores (Mammalia, carnivore). J. Zool. 2005, 266, 133–151. [Google Scholar] [CrossRef]
- Toledo-González, V.; Ortega-Ojeda, F.; Fonseca, G.M.; García-Ruiz, C.; Navarro-Cáceres, P.; Pérez-Lloret, P.; Marín-García, M.P. A morphological and morphometric dental analysis as a forensic tool to identify the iberian wolf (Canis lupus signatus). Animals 2020, 10, 975. [Google Scholar] [CrossRef]
- Yravedra, J.; Andrés, M.; Domínguez-Rodrigo, M. A taphonomic study of the African Wild Dog (Lycaon pictus). Archaeol. Anthropol. Sci. 2014, 6, 113–124. [Google Scholar] [CrossRef]
- Moclán, A.; Domínguez-Rodrigo, M.; Yravedra, J. Classifying agency in bone breakage: An experimental analysis of fracture planes to differentiate between hominin and carnivore dynamic and static loading using machine learning (ML) algorithms. Archaeol. Anthropol. Sci. 2019, 11, 4663–4680. [Google Scholar] [CrossRef]
- Wabakken, P.; Sand, H.; Kojola, I.; Zimmermann, B.; Arnemo, J.M.; Pedersen, H.C.; Liberg, O. Multistage, long-range natal dispersal by a global positioning system-collared Scandinavian wolf. J. Wildl. Manag. 2007, 71, 1631–1634. [Google Scholar] [CrossRef]
- Kojola, I.; Aspi, J.; Hakala, A.; Heikkinen, S.; Ilmoni, C.; Ronkainen, S. Dispersal in an Expanding Wolf Population in Finland. J. Mammal. 2006, 87, 281–286. [Google Scholar] [CrossRef]
- Kojola, I.; Kaartinen, S.; Hakala, A.; Heikkinen, S.; Voipio, H.M. Dispersal behavior and the connectivity between Wolf populations in Northern Europe. J. Wildl. Manag. 2009, 73, 309–313. [Google Scholar] [CrossRef]
- Ciucci, P.; Reggioni, W.; Maiorano, L.; Boitani, L. Long-distance dispersal of a rescued Wolf from the Northern Apennines to the Western Alps. J. Wildl. Manag. 2009, 73, 1300–1306. [Google Scholar] [CrossRef]
- Ražen, N.; Brugnoli, A.; Castagna, C.; Groff, C.; Kaczensky, P.; Kljun, F.; Kos, I.; Krofel, M.; Luštrik, R.; Majić, A.; et al. Long-distance dispersal connects Dinaric-Balkan and Alpine grey wolf (Canis lupus) populations. Eur. J. Wildl. Res. 2015, 62, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.E.; Webster, S.C.; Lance, S.L.; Love, C.N.; Hinton, T.G.; Shamovich, D.; Beasley, J.C. Evidence of long-distance dispersal of a gray wolf from the Chernobyl Exclusion Zone. Eur. J. Wildl. Research. 2018, 64, 39. [Google Scholar] [CrossRef]
- Packila, M.L.; Riley, M.D.; Spence, R.S.; Inman, R.M. Long-distance wolverine dispersal from Wyoming to historic range in Colorad. Northwest. Sci. 2017, 91, 399–407. [Google Scholar] [CrossRef]
- Barry, T.; Gurarie, E.; Cheraghi, F.; Kojola, I.; Fagan, W. Does dispersal make the heart grow bolder? Avoidance of anthropogenic habitat elements across wolf life history. Anim. Behav. 2020, 166, 219–231. [Google Scholar] [CrossRef]
- Maté-González, M.Á.; Aramendi, J.; Yravedra, J.; González-Aguilera, D. Statistical Comparison between Low-Cost Methods for 3D Characterization of Cut-Marks on Bones. Remote Sens. 2017, 9, 873. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.K. tpsDig2 v.2.29; Ecology & Evolution and Anthropology, Stony Brook University: New York, NY, USA, 2021; Available online: http://life.bio.sunsyb.edu/morph/ (accessed on 30 July 2021).
- Bello, S.M.; Soligo, C. A new method for the quantitative analysis of cutmark micromorphology. J. Archaeol. Sci. 2008, 35, 1542–1552. [Google Scholar] [CrossRef]
- Maté-González, M.Á.; Yravedra, J.; González-Aguilera, D.; Palomeque-González, J.F.; Domínguez-Rodrigo, M. Micro-photogrammetric characterization of cut marks on bones. J. Archaeol. Sci. 2015, 62, 128–142. [Google Scholar] [CrossRef]
- Razali, N.M.; Wah, Y.B. Power comparisons of Shapiro-Wilk, Kolmogorov-Smirnov, Lilliefors and Anderson-Darling tests. J. Stat. Model. Anal. 2011, 2, 21–33. [Google Scholar]
- Höhle, J.; Höhle, M. Accuracy assessment of digital elevation models by means of robust statistical methods. ISPRS J. Photogramm. Remote Sens. 2009, 64, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Gonzálvez, P.; Garcia-Gago, J.; Gomez-Lahoz, J.; González-Aguilera, D. Confronting passive and active sensors with non-gaussian statistics. Sensors 2014, 14, 13759–13777. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Martín, M.; Rodríguez-Gonzálvez, P.; Ruiz de Oñá Crespo, E.; González-Aguilera, D. Validation of portable mobile mapping system for inspection tasks in termal and fluid-mechanical facilities. Remote Sens. 2019, 11, 2205. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. Statistical Power Analysis for Behavioural Sciences; Routledge: New York, NY, USA, 1988. [Google Scholar]
- Schurimann, D.L. A comparison of the two one-sided test procedure and the power approach for assessing the equivalence of average biovariability. J. Pharmacokinet. Biopharm. 1987, 15, 657–680. [Google Scholar] [CrossRef] [Green Version]
- Yuen, K.K.; Dixon, W.J. The approximate behaviour and performance of the two-sample trimmed t. Biometrika 1973, 60, 369–374. [Google Scholar] [CrossRef]
- Yuen, K.K. The two-sample trimmed t for unequal population variances. Biometrika 1974, 61, 165–170. [Google Scholar] [CrossRef]
- Dixon, P.M.; Saint-Maurice, P.F.; Kim, Y.; Hibbing, P.; Bai, Y.; Welk, G.J. A primer on the use of equivalence testing for evaluating measurement agreement. Med. Sci. Sports Exerc. 2018, 50, 837–845. [Google Scholar] [CrossRef] [Green Version]
- Mardia, K.V.; Jupp, P.E. Directional Statistics; John Wiley: Chichester, UK, 1999. [Google Scholar]
- Best, D.; Fisher, N. Efficient simulation of the von mises distribution. Appl. Stats. 1979, 28, 152–157. [Google Scholar] [CrossRef]
- von Mises, R. Über die “ganzzahligkeit” der atomgewichte und verwandte fragen. Phys. Z. 1918, 19, 490–500. [Google Scholar]
- Mardia, K.V. Statistics of Directional Data; Academic Press: London, UK, 1972. [Google Scholar]
- Watson, G.S.; Williams, E.J. On the construction of significance tests on the circle and on the sphere. Biometrika 1956, 43, 344–352. [Google Scholar] [CrossRef]
- Pewsey, A. Testing circular symmetry. Can. J. Stats. 2002, 30, 591–600. [Google Scholar] [CrossRef]
- Watson, G.S. Statistics on Spheres; John Wiley: New York, NY, USA, 1983. [Google Scholar]
- Fisher, N.I. Statistical Analysis of Circular Data; Cambridge University Press: Camridge, UK, 1993. [Google Scholar]
- Wheeler, S.; Watson, G.S. A distribution-free two-sample test on the circle. Biometrika 1964, 51, 256–257. [Google Scholar] [CrossRef]
- Hinton, G.E.; Roweis, S.T. Stochastic neighbor embedding. In NIPS 2002, 15, 833–840. [Google Scholar]
- Pearson, K. Note on regression and inheritance in the case of two parents. Proc. R. Soc. Lond. 1895, 58, 347–352. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Hafner Publishing Co.: New York, NY, USA, 1955. [Google Scholar]
- Martin, O. Bayesian Analysis with Python, 2nd ed.; Packt: Birmingham, UK, 2018. [Google Scholar]
- Hoffman, M.D.; Gelman, A. The No-U-Turn Sampler: Adaptively Setting Path Lengths in Hamiltonian Monte Carlo. J. Mach. Learn. Res. 2011, 15, 1593–1623. [Google Scholar]
- Krushke, J.K. Doing Bayesian Data Analysis, 2nd ed.; Academic Press: New York, NY, USA, 2014. [Google Scholar]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis; John Wiley and Sons: New York, NY, USA, 1998. [Google Scholar]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. A field comes of age: Geometric morphometrics in the 21st century. Hystrix 2013, 24, 7–14. [Google Scholar] [CrossRef]
- Oxnard, C.E. The measurement of form: Beyond biometrics. Cleft Palate J. Suppl. 1986, 23, 110–128. [Google Scholar]
- Goodall, C.R. Procrustes methods in the statistical analysis of shape. J. R Stat. Soc. B 1991, 53, 285–339. [Google Scholar] [CrossRef]
- Jungers, W.L.; Falsetti, A.B.; Wall, C.E. Shape, relative size, and size-adjustments in morphometrics. Am. J. Phys. Anthropol. 1995, 38, 137–161. [Google Scholar] [CrossRef]
- Bookstein, F.L. Principal warps: Thin plate spline and the decomposition of deformations. Trans. Pattern Anal. Mach. Intel. 1989, 11, 567–585. [Google Scholar] [CrossRef] [Green Version]
- Wasserstein, R.L.; Lazar, N.A. The ASA Statement on p-Values: Context, process and purpose. Am. Stat. 2016, 70, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Wasserstein, R.L.; Schirm, A.L.; Lazar, N.A. Moving to a world beyond “p < 0.05”. Am. Stat. 2019, 73, 1–19. [Google Scholar]
- Benjamin, D.J.; Berger, J.O. Three recommendations for improving the use of p-values. Am. stat. 2019, 73, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Colquhoun, D. The False Positive Risk: A proposal concerning what to do about p-values. Am. Stat. 2019, 73, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.A. Statistical Methods for Research Workers; Oliver and Boyd: Edinburgh, Scotland, 1925. [Google Scholar]
- Courtenay, L.A.; González-Aguilera, D.; Lagüela, S.; del Pozo, S.; Ruiz Méndez, C.; Barbero-García, I.; Román-Curto, C.; Cañueto, J.; Santos-Durán, C.; Cardeñoso-Álvarez, M.E.; et al. Hyperspectral imaging and robust statistics in non-melanoma skin cancer análisis. Biomed. Optics. Express. 2021, 12, 5107–5127. [Google Scholar] [CrossRef]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–307. [Google Scholar] [CrossRef]
- Mason, G.; Burn, C.C.; Dallaire, J.A.; Kroshko, J.; Kinkaid, H.D.; Jeschke, J.M. Plastic animals in cages: Behavioural flexibility and responses to captivity. Anim. Behav. 2013, 85, 1113–1126. [Google Scholar] [CrossRef]
- Mason, G.; Clubb, R.; Latham, N.; Vickery, S. Why and how should we use environmental enrichment to tackle stereotypic behavior? Appl. Anim. Behav. Sci. 2007, 102, 163–188. [Google Scholar] [CrossRef] [Green Version]
- Rose, P.E.; Nash, S.M.; Riley, L.M. To pace or not to pace? A review of what abnormal repetitive behavior tells us about zoo animal management. J. Vet. Behav. 2017, 20, 11–21. [Google Scholar] [CrossRef]
- Mason, G.J. Species differences in response to captivity: Stress, welfare and the comparative method. Trends Ecol. Evol. 2010, 25, 713–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, D.L. A note on the influence of visitors on the behavior and welfare of zoo-housed gorillas. Appl. Anim. Behav. Sci. 2005, 93, 13–17. [Google Scholar] [CrossRef]
- Wheat, C.H.; Temrin, H. Intrinsic Ball Retrieving in Wolf Puppies Suffests Standing Ancestral Variation for Human-Directed Play Behavior. Iscience 2020, 23, 100811. [Google Scholar] [CrossRef] [Green Version]
- Carlstead, K. Husbandry of the Fennec Fox (Fennecus zerda): Environmental conditions influencing stereotypic behavior. Int. Zoo Yb. 1991, 30, 202–207. [Google Scholar] [CrossRef]
- Clubb, R.; Mason, G. Captivity Effects on Wide-Ranging Carnivores. Nat. Brief. Comm. 2003, 425, 473. [Google Scholar] [CrossRef] [PubMed]
- Kawata, K. Zoo Animal Feeding: A Natural History Viewpoint. Der. Zool. Garten. 2008, 78, 17–42. [Google Scholar] [CrossRef]
- Riemer, S. Effectiveness of treatments for firework feats in dogs. J. Vet. Behav. 2020, 37, 61–70. [Google Scholar] [CrossRef]
- Sherman, B.L.; Mills, D.S. Canine anxieties and phobias: An update on separation anxiety and noise adversions. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 1081–1106. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, K.C. Seperation, confinement, or noises: What is scaring that dog? Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 367–386. [Google Scholar] [CrossRef]
- Storengen, L.M.; Lingaas, F. Noise sensitivity in 17 dog breeds: Prevalence, breed risk and correlation with fear in other situations. Appl. Anim. Behav. Sci. 2015, 171, 152–160. [Google Scholar] [CrossRef]
- Haynes, G. A guide for differentiating mammalian carnivore taxa responsible for gnaw damage to herbivore limb bones. Paleobiology 1983, 9, 164–172. [Google Scholar] [CrossRef]
- Saladié, P.; Huguet, R.; Díez, C.; Rodríguez-Hidalgo, A.; Carbonell, E. Taphonomic Modifications Produced by Modern Brown Bears (Ursus arctos). Int. J. Osteoarchaeol. 2013, 23, 13–33. [Google Scholar] [CrossRef]
- Sala, N.; Arsuaga, J.L. Taphonomic studies with wild brown bears (Ursus arctos) in the mountains of northern Spain. J. Archaeol. Sci. 2013, 40, 1389–1396. [Google Scholar] [CrossRef]
- Arilla, M.; Rosell, J.; Blasco, R.; Domínguez-Rodrigo, M.; Pickering, T.R. The ‘‘Bear’’ Essentials: Actualistic Research on Ursus arctos arctos in the Spanish Pyrenees and Its Implications for Paleontology and Archaeology. PLoS ONE 2014, 9, e102457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sala, N.; Arsuaga, J.L.; Haynes, G. Taphonomic comparison of bone modifications caused by wild and captive wolves (Canis lupus). Quat. Int. 2014, 330, 126–135. [Google Scholar] [CrossRef]
- Ferretti, M.P. Evolution of Bone-Cracking Adaptations in Hyaenids (Mammali, Carnivore). Swiss J. Geosci. 2007, 100. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Min 1 | k-Hat | s-Hat | v | Uniformity | Symmetry | Central 1,2 | |||
|---|---|---|---|---|---|---|---|---|---|---|
| t | p | t | p | |||||||
| Cabárceno | 120.73 | 2.28 | 2.28 | 0.02 | 0.98 | 1.1 × 10−23 | 1.63 | 0.110 | 158.67 | 176.39 |
| Hosquillo | 112.00 | 0.25 | 1.06 | 0.02 | 0.98 | 1.2 × 10−46 | 1.77 | 0.079 | 147.09 | 175.12 |
| Flechas 3 | 101.65 | −1.31 | −0.14 | 0.03 | 0.97 | 3.1 × 10−26 | 0.24 | 0.810 | 132.51 | 160.28 |
| Villardeciervos 3 | 110.80 | −1.49 | 0.41 | 0.01 | 0.99 | 6.3 × 10−24 | 0.75 | 0.447 | 132.19 | 149.96 |
| Captive | 112.00 | 0.27 | 1.32 | 0.03 | 0.97 | 3.2 × 10−69 | 2.60 | 0.007 | 151.40 | 176.39 |
| Wild | 101.65 | −0.96 | −0.03 | 0.02 | 0.98 | 2.0 × 10−49 | 0.06 | 0.953 | 132.36 | 160.28 |
| Mean | Median | Distribution | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sample 1 | Sample 2 | Yg | p | p(H0) | Pg | p | p(H0) | Wg | p | p(H0) |
| Wild * | Captive | 138.80 | 0.0001 | 0.0025 | 78.53 | 0.0001 | 0.0025 | 69.97 | 7.3 × 10−16 | 6.9 × 10−14 |
| Cabárceno | Flechas * | 105.98 | 0.0001 | 0.0025 | 50.94 | 0.0001 | 0.0025 | 47.06 | 6.0 × 10−11 | 3.8 × 10−09 |
| Cabárceno | Hosquillo | 20.98 | 0.0001 | 0.0025 | 16.99 | 0.0001 | 0.0025 | 12.77 | 1.7 × 10−03 | 2.9 × 10−02 |
| Cabárceno | Villardeciervos * | 143.00 | 0.0001 | 0.0025 | 55.31 | 0.0001 | 0.0025 | 64.27 | 1.1 × 10−14 | 9.6 × 10−13 |
| Flechas * | Hosquillo | 50.69 | 0.0001 | 0.0025 | 29.68 | 0.0001 | 0.0025 | 21.69 | 2.0 × 10−05 | 5.9 × 10−04 |
| Flechas * | Villardeciervos * | 0.022 | 0.8864 | 0.7749 | 0.22 | 0.6978 | 0.5943 | 4.650 | 0.1000 | 0.3850 |
| Hosquillo | Villardeciervos * | 59.03 | 0.0001 | 0.0025 | 41.36 | 0.0001 | 0.0025 | 34.08 | 4.0 × 10−08 | 5.0 × 10−07 |
| Measurement | Cabárceno | Hosquillo | Flechas 1 | Villardeciervos 1 | Captive | Wild 1 | |
|---|---|---|---|---|---|---|---|
| WIS | Min. | 0.15 | 0.07 | 0.11 | 0.21 | 0.07 | 0.11 |
| Central 2 | 0.54 | 0.48 | 0.47 | 0.56 | 0.50 | 0.49 | |
| Deviation 3 | 0.40 | 0.27 | 0.22 | 0.31 | 0.33 | 0.27 | |
| Max. | 1.75 | 1.78 | 1.16 | 1.38 | 1.78 | 0.14 | |
| WIM | Min. | 0.11 | 0.05 | 0.08 | 0.15 | 0.05 | 0.08 |
| Central 2 | 0.37 | 0.33 | 0.29 | 0.39 | 0.34 | 0.33 | |
| Deviation 3 | 0.27 | 0.19 | 0.15 | 0.21 | 0.23 | 0.18 | |
| Max. | 1.18 | 1.20 | 0.80 | 0.95 | 1.20 | 0.95 | |
| WIB | Min. | 0.06 | 0.03 | 0.04 | 0.08 | 0.02 | 0.04 |
| Central 2 | 0.19 | 0.17 | 0.16 | 0.21 | 0.18 | 0.17 | |
| Deviation 3 | 0.14 | 0.10 | 0.08 | 0.11 | 0.12 | 0.10 | |
| Max. | 0.60 | 0.61 | 0.43 | 0.50 | 0.61 | 0.50 | |
| D | Min. | 0.00 | 0.01 | 0.02 | 0.04 | 0.00 | 0.02 |
| Central 2 | 0.05 | 0.07 | 0.09 | 0.12 | 0.07 | 0.11 | |
| Deviation 3 | 0.05 | 0.06 | 0.07 | 0.08 | 0.06 | 0.08 | |
| Max. | 0.24 | 0.31 | 0.31 | 0.37 | 0.31 | 0.37 | |
| RDC | Min. | 0.08 | 0.04 | 0.06 | 0.12 | 0.04 | 0.06 |
| Central 2 | 0.28 | 0.26 | 0.24 | 0.31 | 0.26 | 0.27 | |
| Deviation 3 | 0.21 | 0.15 | 0.13 | 0.18 | 0.17 | 0.15 | |
| Max. | 0.90 | 0.91 | 0.65 | 0.76 | 0.91 | 0.76 | |
| LDC | Min. | 0.08 | 0.04 | 0.06 | 0.11 | 0.04 | 0.06 |
| Central 2 | 0.29 | 0.26 | 0.24 | 0.31 | 0.27 | 0.26 | |
| Deviation 3 | 0.21 | 0.16 | 0.13 | 0.17 | 0.17 | 0.15 | |
| Max. | 0.90 | 0.92 | 0.65 | 0.78 | 0.92 | 0.78 | |
| Cabárceno | Flechas 1 | Hosquillo | ||
|---|---|---|---|---|
| Flechas 1 | |d| | 1.779 | ||
| p | 1.000 | |||
| p(H0) | 0.999 | |||
| Hosquillo | |d| | 0.483 | 1.297 | |
| p | 0.689 | 0.995 | ||
| p(H0) | 0.589 | 0.986 | ||
| Villardeciervos 1 | |d| | 2.767 | 0.988 | 2.285 |
| p | 1.000 | 0.798 | 1.000 | |
| p(H0) | 1.000 | 0.671 | 1.000 | |
| Cabárceno | Flechas 1 | Hosquillo | ||
|---|---|---|---|---|
| Flechas 1 | |d| | 0.062 | ||
| p | 2.5 × 10−11 | |||
| p(H0) | 1.7 × 10−09 | |||
| Hosquillo | |d| | 0.013 | 0.050 | |
| p | 3.2 × 10−18 | 4.9 × 10−16 | ||
| p(H0) | 3.5 × 10−16 | 4.7 × 10−14 | ||
| Villardeciervos 1 | |d| | 0.034 | 0.028 | 0.021 |
| p | 6.8 × 10−16 | 9.4 × 10−19 | 4.4 × 10−26 | |
| p(H0) | 6.5 × 10−14 | 1.0 × 10−15 | 7.0 × 10−24 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Courtenay, L.A.; Herranz-Rodrigo, D.; Yravedra, J.; Vázquez-Rodríguez, J.M.; Huguet, R.; Barja, I.; Maté-González, M.Á.; Fernández, M.F.; Muñoz-Nieto, Á.-L.; González-Aguilera, D. 3D Insights into the Effects of Captivity on Wolf Mastication and Their Tooth Marks; Implications in Ecological Studies of Both the Past and Present. Animals 2021, 11, 2323. https://doi.org/10.3390/ani11082323
Courtenay LA, Herranz-Rodrigo D, Yravedra J, Vázquez-Rodríguez JM, Huguet R, Barja I, Maté-González MÁ, Fernández MF, Muñoz-Nieto Á-L, González-Aguilera D. 3D Insights into the Effects of Captivity on Wolf Mastication and Their Tooth Marks; Implications in Ecological Studies of Both the Past and Present. Animals. 2021; 11(8):2323. https://doi.org/10.3390/ani11082323
Chicago/Turabian StyleCourtenay, Lloyd A., Darío Herranz-Rodrigo, José Yravedra, José Mª Vázquez-Rodríguez, Rosa Huguet, Isabel Barja, Miguel Ángel Maté-González, Maximiliano Fernández Fernández, Ángel-Luis Muñoz-Nieto, and Diego González-Aguilera. 2021. "3D Insights into the Effects of Captivity on Wolf Mastication and Their Tooth Marks; Implications in Ecological Studies of Both the Past and Present" Animals 11, no. 8: 2323. https://doi.org/10.3390/ani11082323
APA StyleCourtenay, L. A., Herranz-Rodrigo, D., Yravedra, J., Vázquez-Rodríguez, J. M., Huguet, R., Barja, I., Maté-González, M. Á., Fernández, M. F., Muñoz-Nieto, Á.-L., & González-Aguilera, D. (2021). 3D Insights into the Effects of Captivity on Wolf Mastication and Their Tooth Marks; Implications in Ecological Studies of Both the Past and Present. Animals, 11(8), 2323. https://doi.org/10.3390/ani11082323