
Role of Infection and Immunity in Bovine Perinatal Mortality: Part 2. Fetomaternal Response to Infection and Novel Diagnostic Perspectives


{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Diagnosis of Infectious Stillbirth
3. Development of Immune System in Bovine Fetuses and Response to Antigenic Stimulation in Utero
3.1. Development of the Cellular Components of the Bovine Fetal Immune System
Complement in the Fetus
3.2. Humoral Immune Response of the Bovine Fetus
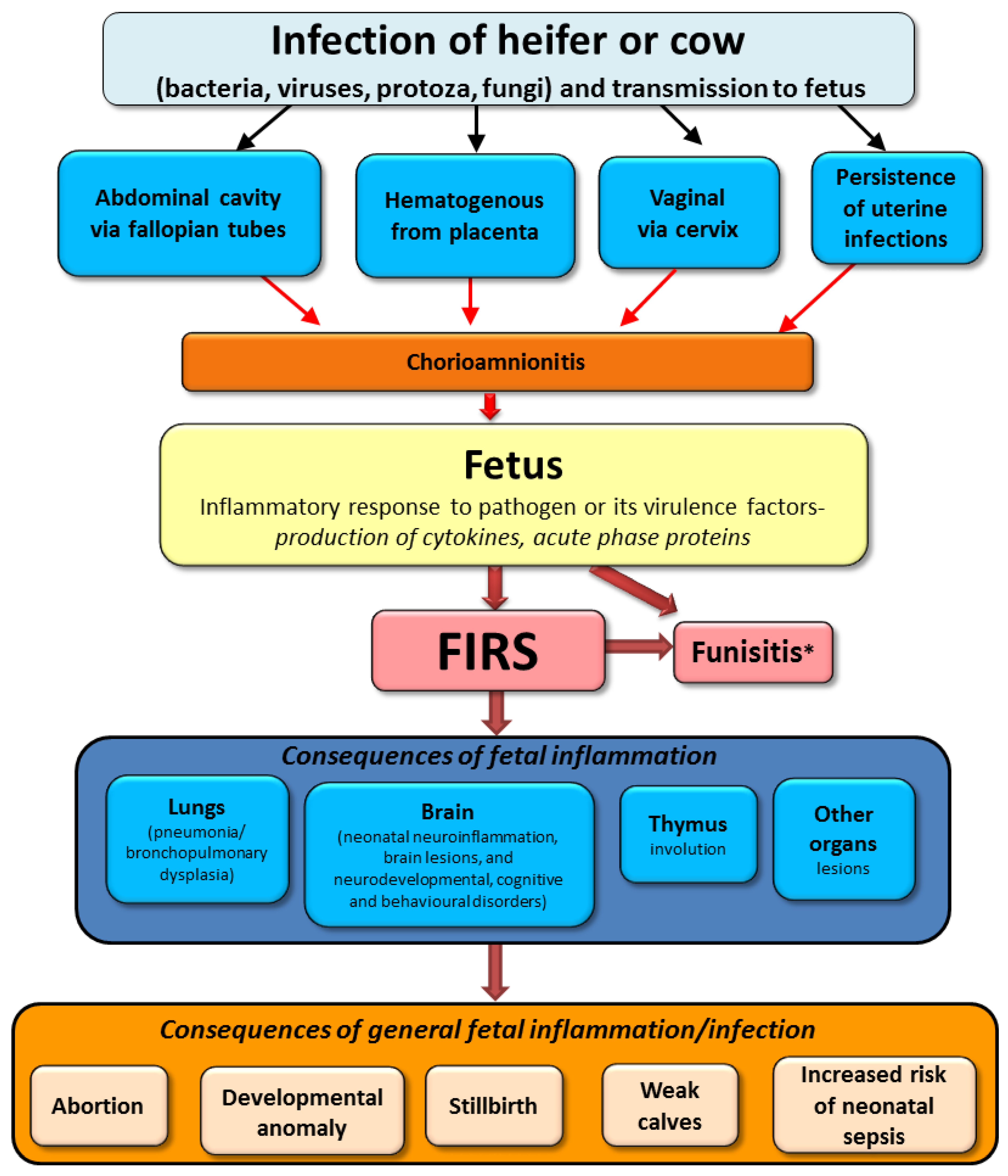
3.3. Fetal Inflammatory Response Syndrome (FIRS)
3.3.1. FIRS in Humans Fetuses
3.3.2. FIRS in Animal Fetuses
3.4. Brain
3.5. Lungs
3.6. Thymus
4. Novel Approaches to Diagnosis Causes of Stillbirth
4.1. Monitoring of Fetus Responses
4.1.1. Fetal Humoral Immune Response
4.1.2. Fetal Acute Phase Response
4.1.3. Fetal Metabolic Response-Perspectives
4.2. Monitoring of Dam Nonspecific Responses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berglund, B.; Steinbock, L.; Elvander, M. Causes of stillbirth and time of death in Swedish Holstein calves examined post mortem. Acta Vet. Scand. 2003, 44, 111–120. [Google Scholar] [CrossRef]
- Mee, J.F.; Berry, D.P.; Cromie, A.R. Prevalence of, and risk factors associated with, perinatal calf mortality in pasture-based Holstein-Friesian cows. Animal 2008, 2, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Cuttance, E.; Laven, R. Perinatal mortality risk factors in dairy calves. Vet. J. 2019, 253, 105394. [Google Scholar] [CrossRef]
- Ghavi Hossein-Zadeh, N.; Nejati-Javaremi, A.; Miraei-Ashtiani, S.R.; Kohram, H. An Observational Analysis of Twin Births, Calf Stillbirth, Calf Sex Ratio, and Abortion in Iranian Holsteins. J. Dairy Sci. 2008, 91, 4198–4205. [Google Scholar] [CrossRef] [PubMed]
- Fourichon, C.; Beaudeau, F.; Bareille, N.; Seegers, H. Incidence of health disorders in dairy farming systems in western France. Livest. Prod. Sci. 2001, 68, 157–170. [Google Scholar] [CrossRef]
- Chassagne, M.; Barnouin, J.; Chacornac, J.P. Risk factors for stillbirth in Holstein heifers under field conditions in France: A prospective survey. Theriogenology 1999, 51, 1477–1488. [Google Scholar] [CrossRef]
- López Helguera, I.; Behrouzi, A.; Kastelic, J.P.; Colazo, M.G. Risk factors associated with dystocia in a tie stall dairy herd. Can. J. Anim. Sci. 2016, 96, 135–142. [Google Scholar] [CrossRef]
- Mee, J.F.; Sánchez-Miguel, C.; Doherty, M. Influence of modifiable risk factors on the incidence of stillbirth/perinatal mortality in dairy cattle. Vet. J. 2014, 199, 19–23. [Google Scholar] [CrossRef]
- Hansen, M.; Misztal, I.; Lund, M.S.; Pedersen, J.; Christensen, L.G. Undesired phenotypic and genetic trend for stillbirth in danish holsteins. J. Dairy Sci. 2004, 87, 1477–1486. [Google Scholar] [CrossRef]
- Mee, J.F. Epidemiology of bovine perinatal mortality. In Bovine Prenatal, Perinatal and Neonatal Medicine; Szenci, O., Mee, J.F., Bleul, U., Taverne, M.A.M., Eds.; Hungarian Association for Buiatrics: Budapest, Hungary, 2021; in press. [Google Scholar]
- Heringstad, B.; Chang, Y.M.; Svendsen, M.; Gianola, D. Genetic analysis of calving difficulty and stillbirth in norwegian red cows. J. Dairy Sci. 2007, 90, 3500–3507. [Google Scholar] [CrossRef]
- Mahnani, A.; Sadeghi-Sefidmazgi, A.; Keshavarzi, H. Performance and financial consequences of stillbirth in Holstein dairy cattle. Animal 2018, 12, 617–623. [Google Scholar] [CrossRef]
- Khodakaram-Tafti, A.; Ikede, B.O. A retrospective study of sporadic bovine abortions, stillbirths, and neonatal abnormalities in Atlantic Canada, from 1990 to 2001. Can. Vet. J. 2005, 46, 635–637. [Google Scholar]
- SAC CVS DISEASE SURVEILLANCE REPORT Surveillance: Causes of abortion and stillbirth in cattle in Scotland. Vet. Rec. 2009, 165, 677–680. [CrossRef]
- Waldner, C.L.; Kennedy, R.I.; Rosengren, L.B.; Pollock, C.M.; Clark, E.G. Gross postmortem and histologic examination findings from abortion losses and calf mortalities in western Canadian beef herds. Can. Vet. J. 2010, 51, 1227–1238. [Google Scholar]
- Syrjälä, P.; Anttila, M.; Dillard, K.; Fossi, M.; Collin, K.; Nylund, M.; Autio, T. Causes of bovine abortion, stillbirth and neonatal death in Finland 1999–2006. Acta Vet. Scand. 2007, 49, S3. [Google Scholar] [CrossRef]
- Muskens, J.; Wouda, W.; von Bannisseht-Wijsmuller, T.; van Maanen, C. Prevalence of Coxiella burnetii infections in aborted fetuses and stillborn calves. Vet. Rec. 2012, 170, 260. [Google Scholar] [CrossRef] [PubMed]
- Jawor, P.; Król, D.; Mee, J.F.; Sołtysiak, Z.; Dzimira, S.; Larska, M.; Stefaniak, T. Infection exposure, detection and causes of death in perinatal mortalities in Polish dairy herds. Theriogenology 2017, 103. [Google Scholar] [CrossRef] [PubMed]
- Kirkbride, C.A. Etiologic agents detected in a 10-year study of bovine abortions and stillbirths. J. Vet. Diagn. Investig. 1992, 4, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Mee, J.F.; Jawor, P.; Stefaniak, T. Role of Infection and Immunity in Bovine Perinatal Mortality: Part 1. Causes and Current Diagnostic Approaches. Animals 2021, 11, 1033. [Google Scholar] [CrossRef] [PubMed]
- Mee, J.F.; Szenci, O. Selected pathological causes of bovine stillbirth illustrated with photographic images. Magy. Allatorvosok Lapja 2012, 134, 718–725. [Google Scholar]
- Cutlip, R.; McClurkin, A.W. Lesions and pathogenesis of disease in young calves experimentally induced by a bovine adenovirus type 5 isolated from a calf with weak calf syndrome. Am. J. Vet. Res. 1975, 36, 1095–1098. [Google Scholar]
- Fulton, R.W.; Whitley, E.M.; Johnson, B.J.; Ridpath, J.F.; Kapil, S.; Burge, L.J.; Cook, B.J.; Confer, A.W. Prevalence of bovine viral diarrhea virus (BVDV) in persistently infected cattle and BVDV subtypes in affected cattle in beef herds in south central United States. Can. J. Vet. Res. 2009, 73, 283–291. [Google Scholar]
- Haddad, J.P.; Dohoo, I.R.; VanLeewen, J. A review of Neospora caninum in dairy and beef cattle—A Canadian perspective. Can. Vet. J. 2005, 46, 230–243. [Google Scholar]
- Smyth, J.A.; Fitzpatrick, D.A.; Ellis, W.A. Stillbirth/perinatal weak calf syndrome: A study of calves infected with Leptospira. Vet. Rec. 1999, 145, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Done, J.T.; Terlecki, S.; Richardson, C.; Harkness, J.W.; Sands, J.J.; Patterson, D.S.; Sweasey, D.; Shaw, I.G.; Winkler, C.E.; Duffell, S.J. Bovine virus diarrhoea-mucosal disease virus: Pathogenicity for the fetal calf following maternal infection. Vet. Rec. 1980, 106, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Hauth, J.C.; Andrews, W.W. Intrauterine Infection and Preterm Delivery. N. Engl. J. Med. 2000, 342, 1500–1507. [Google Scholar] [CrossRef]
- Miller, R.B. A summary of some of the pathogenetic mechanisms involved in bovine abortion. Can. Vet. J. 1977, 18, 87–95. [Google Scholar]
- Kirkbride, C.A. Bacterial Agents Detected in a Lo-Year Study of Bovine Abortions and Stillbirths. J. Vet. Diagn. Investig. 1993, 5, 64–68. [Google Scholar] [CrossRef]
- Zhang, L.; Xia, Y.; Tang, F.; Li, S.; Yang, L.; Wang, B. The regulation of intrauterine immune cytokines and chemokines during early pregnancy in the bovine. Large Anim. Rev. 2015, 21, 23–31. [Google Scholar]
- Regidor-Cerrillo, J.; Arranz-Solís, D.; Benavides, J.; Gómez-Bautista, M.; Castro-Hermida, J.A.; Mezo, M.; Pérez, V.; Ortega-Mora, L.M.; González-Warleta, M. Neospora caninum infection during early pregnancy in cattle: How the isolate influences infection dynamics, clinical outcome and peripheral and local immune responses. Vet. Res. 2014, 45, 10. [Google Scholar] [CrossRef]
- Cortese, V.S. Neonatal Immunology. Vet. Clin. North Am. Food Anim. Pract. 2009, 25, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Schultz, R.D.; Dunne, H.W.; Heist, C.E. Ontogeny of the bovine immune response. Infect. Immun. 1973, 7, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Stefaniak, T.; Chełmońska-Soyta, A.; Bajzert, J.; Jawor, P.; Rzasa, A.; Sitnik, O. Prenatal and postnatal development of immune system in ruminants. Med. Weter. 2012, 68, 534–539. [Google Scholar]
- Schultz, R.D.; Confer, F.; Dunne, H.W. Occurrence of blood cells and serum proteins in bovine fetuses and calves. Can. J. Comp. Med. 1971, 35, 93–98. [Google Scholar]
- Banks, K.L.; McGuire, T.C. Neonatal immunology. In Veterinary Clinical Immunology; Halliwell, R.E.W., Gorman, N.T., Eds.; Saunders: Philadelphia, PA, USA, 1989; pp. 193–204. [Google Scholar]
- Tizzard, I.R. Immunity in the fetus and newborn. In Veterinary Immunology: An Introduction; Saunders: Philadelphia, PA, USA, 2000; pp. 210–221. [Google Scholar]
- Innes, E.A.; Wright, S.; Bartley, P.; Maley, S.; Macaldowie, C.; Esteban-Redondo, I.; Buxton, D. The host-parasite relationship in bovine neosporosis. Vet. Immunol. Immunopathol. 2005, 108, 29–36. [Google Scholar] [CrossRef]
- Yasuda, M.; Fujino, M.; Nasu, T.; Murakami, T. Histological studies on the ontogeny of bovine gut-associated lymphoid tissue: Appearance of T cells and development of IgG+ and IgA+ cells in lymphoid follicles. Dev. Comp. Immunol. 2004, 28, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Montejo, S.; Collantes-Fernández, E.; Blanco-Murcia, J.; Rodríguez-Bertos, A.; Risco-Castillo, V.; Ortega-Mora, L.M. Experimental infection with a low virulence isolate of Neospora caninum at 70 days gestation in cattle did not result in foetopathy. Vet. Res. 2009, 40. [Google Scholar] [CrossRef] [PubMed]
- Gibney, E.H.; Kipar, A.; Rosbottom, A.; Guy, C.S.; Smith, R.F.; Hetzel, U.; Trees, A.J.; Williams, D.J.L. The extent of parasite-associated necrosis in the placenta and foetal tissues of cattle following Neospora caninum infection in early and late gestation correlates with foetal death. Int. J. Parasitol. 2008, 38, 579–588. [Google Scholar] [CrossRef]
- Osburn, B.I.; MacLachlan, N.J.; Terrell, T.G. Ontogeny of the immune system. J. Am. Vet. Med. Assoc. 1982, 181, 1049–1052. [Google Scholar]
- Woodard, L.F.; Eckblad, W.P.; Olson, D.P.; Bull, R.C.; Everson, D.O. Hemolytic complement levels of neonatal calves delivered from protein-energy malnourished dams and subjected to cold stress. Cornell Vet. 1980, 70, 266–271. [Google Scholar]
- Ellis, W.A.; Logan, E.F.; O’Brien, J.J. Serum immunoglobulins in aborted and non-aborted bovine foetuses. Clin. Exp. Immunol. 1978, 33, 136–141. [Google Scholar]
- Tierney, T.J.; Simpson-Morgan, M.W. The immune response of foetal calves. Vet. Immunol. Immunopathol. 1997, 57, 229–238. [Google Scholar] [CrossRef]
- Mullaney, T.P.; Newman, L.E.; Whitehair, C.K. Humoral immune response of the bovine fetus to in utero vaccination with attenuated bovine coronavirus. Am. J. Vet. Res. 1988, 49, 156–159. [Google Scholar]
- Conner, G.H.; Richardson, M.; Carter, G.R.; Wamukoya, J.P. Immune responses of the bovine fetus. J. Dairy Sci. 1977, 60, 289–293. [Google Scholar] [CrossRef]
- Wamukoya, J.P.; Conner, G.H. Local immune responses in the bovine fetus vaccinated in utero with Escherichia coli antigen. Am. J. Vet. Res. 1976, 37, 159–163. [Google Scholar]
- Romero, R.; Gomez, R.; Ghezzi, F.; Yoon, B.H.; Mazor, M.; Edwin, S.S.; Berry, S.M. A fetal systemic inflammatory response is followed by the spontaneous onset of preterm parturition. Am. J. Obstet. Gynecol. 1998, 179, 186–193. [Google Scholar] [CrossRef]
- Jung, E.; Romero, R.; Yeo, L.; Diaz-Primera, R.; Marin-Concha, J.; Para, R.; Lopez, A.M.; Pacora, P.; Gomez-Lopez, N.; Yoon, B.H.; et al. The fetal inflammatory response syndrome: The origins of a concept, pathophysiology, diagnosis, and obstetrical implications. Semin. Fetal Neonatal. Med. 2020, 25, 101146. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.L.; Allred, E.N.; Kliman, H.J.; Zambrano, E.; Doss, B.J.; Husain, A.; Pflueger, S.M.V.; Chang, C.H.; Livasy, C.A.; Roberts, D.; et al. Histological characteristics of singleton placentas delivered before the 28th week of gestation. Pathology 2008, 40, 372–376. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vaisbuch, E.; Romero, R.; Gomez, R.; Kusanovic, J.P.; Mazaki-Tovi, S.; Chaiworapongsa, T.; Hassan, S.S. An elevated fetal interleukin-6 concentration can be observed in fetuses with anemia due to Rh alloimmunization: Implications for the understanding of the fetal inflammatory response syndrome. J. Mater. Fetal Neonatal Med. 2011, 24, 391–396. [Google Scholar] [CrossRef]
- Takahashi, N.; Nagamatsu, T.; Fujii, T.; Takahashi, K.; Tsuchida, Y.; Fujio, K.; Fujii, T. Extremely high levels of multiple cytokines in the cord blood of neonates born to mothers with systemic autoimmune diseases. Cytokine 2020, 127, 154926. [Google Scholar] [CrossRef]
- Bashiri, A.; Burstein, E.; Mazor, M. Cerebral palsy and fetal inflammatory response syndrome: A review. J. Perinat. Med. 2006, 34, 5–12. [Google Scholar] [CrossRef]
- Chaiworapongsa, T.; Romero, R.; Kim, J.C.; Kim, Y.M.; Blackwell, S.C.; Yoon, B.H.; Gomez, R. Evidence for fetal involvement in the pathologic process of clinical chorioamnionitis. Am. J. Obstet. Gynecol. 2002, 186, 1178–1182. [Google Scholar] [CrossRef] [PubMed]
- D’Alquen, D.; Kramer, B.W.; Seidenspinner, S.; Marx, A.; Berg, D.; Groneck, P.; Speer, C.P. Activation of umbilical cord endothelial cells and fetal inflammatory response in preterm infants with chorioamnionitis and funisitis. Pediatr. Res. 2005, 57, 263–269. [Google Scholar] [CrossRef][Green Version]
- Eckersall, P.D.; Conner, J.G. Bovine and canine acute phase proteins. Vet. Res. Commun. 1988, 12, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Gotsch, F.; Romero, R.; Kusanovic, J.P.; Mazaki-Tovi, S.; Pineles, B.L.; Erez, O.; Espinoza, J.; Hassan, S.S. The fetal inflammatory response syndrome. Clin. Obstet. Gynecol. 2007, 50, 652–683. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Chaiyasit, N.; Yoon, B.H.; Kim, Y.M. Acute chorioamnionitis and funisitis: Definition, pathologic features, and clinical significance. Am. J. Obstet. Gynecol. 2015, 213, S29–S52. [Google Scholar] [CrossRef]
- Thomas, W.; Speer, C.P. Chorioamnionitis: Important risk factor or innocent bystander for neonatal outcome? Neonatology 2011, 99, 177–187. [Google Scholar] [CrossRef]
- Grigsby, P.L.; Hirst, J.J.; Scheerlinck, J.P.; Phillips, D.J.; Jenkin, G. Fetal responses to maternal and intra-amniotic lipopolysaccharide administration in sheep. Biol. Reprod. 2003, 68, 1695–1702. [Google Scholar] [CrossRef]
- Gavilanes, A.W.D.; Strackx, E.; Kramer, B.W.; Gantert, M.; Van den Hove, D.; Steinbusch, H.; Garnier, Y.; Cornips, E.; Steinbusch, H.; Zimmermann, L.; et al. Chorioamnionitis induced by intraamniotic lipopolysaccharide resulted in an interval-dependent increase in central nervous system injury in the fetal sheep. Am. J. Obstet. Gynecol. 2009, 200, 437.e1–437.e8. [Google Scholar] [CrossRef]
- Jawor, P.; Mee, J.F.; Stefaniak, T. Perinatal immuno/inflammatory responses in the presence or absence of bovine fetal infection. BMC Vet. Res. 2018, 14. [Google Scholar] [CrossRef]
- Jawor, P.; Stefaniak, T.; Sołtysiak, Z.; Dzimira, S.; Bednarski, M. Salmonella enterica serovar Stanley intrauterine infection in a stillborn calf—Case report. Acta Vet. Brno 2013, 82, 363–367. [Google Scholar] [CrossRef]
- Castell, J.V.; Geiger, T.; Gross, V.; Andus, T.; Walter, E.; Hirano, T.; Kishimoto, T.; Heinrich, P.C. Plasma clearance, organ distribution and target cells of interleukin-6/hepatocyte-stimulating factor in the rat. Eur. J. Biochem. 1988, 177, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Laster, S.M.; Tompkins, M.; Brown, T.; Xu, J.-S.; Altier, C.; Gomez, W.; Benfield, D.; McCaw, M.B. In Utero Infection by Porcine Reproductive and Respiratory Syndrome Virus Is Sufficient to Increase Susceptibility of Piglets to Challenge by Streptococcus suis Type II. J. Virol. 2001, 75, 4889–4895. [Google Scholar] [CrossRef] [PubMed]
- Zahl, P.A.; Bjerknes, C. Induction of Decidua-Placental Hemorrhage in Mice by the Endotoxins of Certain Gram-Negative Bacteria. Proc. Soc. Exp. Biol. Med. 1943, 54, 329–332. [Google Scholar] [CrossRef]
- Nguyen, D.N.; Thymann, T.; Goericke-Pesch, S.K.; Ren, S.; Wei, W.; Skovgaard, K.; Damborg, P.; Brunse, A.; van Gorp, C.; Kramer, B.W.; et al. Prenatal Intra-Amniotic Endotoxin Induces Fetal Gut and Lung Immune Responses and Postnatal Systemic Inflammation in Preterm Pigs. Am. J. Pathol. 2018, 188, 2629–2643. [Google Scholar] [CrossRef]
- Elovitz, M.A.; Brown, A.G.; Breen, K.; Anton, L.; Maubert, M.; Burd, I. Intrauterine inflammation, insufficient to induce parturition, still evokes fetal and neonatal brain injury. Int. J. Dev. Neurosci. 2011, 29, 663–671. [Google Scholar] [CrossRef]
- Wong, D.; Dorovini-Zis, K.; Vincent, S.R. Cytokines, nitric oxide, and cGMP modulate the permeability of an in vitro model of the human blood-brain barrier. Exp. Neurol. 2004, 190, 446–455. [Google Scholar] [CrossRef]
- Hagberg, H.; Mallard, C. Effect of inflammation on central nervous system development and vulnerability. Curr. Opin. Neurol. 2005, 18, 117–123. [Google Scholar] [CrossRef]
- Perry, V.H.; Teeling, J. Microglia and macrophages of the central nervous system: The contribution of microglia priming and systemic inflammation to chronic neurodegeneration. Semin. Immunopathol. 2013, 35, 601–612. [Google Scholar] [CrossRef]
- Perry, V.H.; Hume, D.A.; Gordon, S. Immunohistochemical localization of macrophages and microglia in the adult and developing mouse brain. Neuroscience 1985, 15, 313–326. [Google Scholar] [CrossRef]
- Püntener, U.; Booth, S.G.; Perry, V.H.; Teeling, J.L. Long-term impact of systemic bacterial infection on the cerebral vasculature and microglia. J. Neuroinflamm. 2012, 9, 668. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, D.L.; Van Olphen, A.; Van Campen, H.; Hansen, T.R. Infectious Disease The Fetal Brain in Bovine Viral Diarrhea Virus-infected Calves: Lesions, Distribution, and Cellular Heterogeneity of Viral Antigen at 190 Days Gestation. Vet. Pathol. 2008, 45, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Ohmann, H.B. Experimental fetal infection with bovine viral diarrhea virus. II. Morphological reactions and distribution of viral antigen. Can. J. Comp. Med. Rev. Can. Med. Comp. 1982, 46, 363–369. [Google Scholar]
- Binkhorst, G.J.; Journée, D.L.H.; Wouda, W.; Strayer, P.J.; Vos, J.H.; Binkhorstl, G.J.; Journée, D.L.H.; Wouda, W.; Strayer, P.J.; Vos, J.H. Neurological disorders, virus persistence and hypomyelination in calves due to intra-uterine infections with bovine virus diarrhoea virus I. Clinical symptoms and morphological lesions Neurological disorders, virus persistence and hypomyelination in calve. Vet. Q. 1983, 5, 145–155. [Google Scholar] [CrossRef]
- Barr, B.C.; Rowe, J.D.; Sverlow, K.W.; Bondurant, R.H.; Ardans, A.A.; Oliver, M.N.; Conrad, P.A. Experimental reproduction of bovine fetal Neospora infection and death with a bovine Neospora isolate. J. Vet. Diagn. Investig. 1994, 6, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Wouda, W.; Moen, A.R.; Visser, I.J.R.; Van Knapen, F. Bovine fetal neosporosis: A comparison of epizootic and sporadic abortion cases and different age classes with regard to lesion severity and immunohistochemical identification of organisms in brain, heart, and liver. J. Vet. Diagn. Investig. 1997, 9, 180–185. [Google Scholar] [CrossRef]
- Speer, C.P. Chorioamnionitis, postnatal factors and proinflammatory response in the pathogenetic sequence of bronchopulmonary dysplasia. Neonatology 2009, 95, 353–361. [Google Scholar] [CrossRef]
- Bhandari, A.; Bhandari, V. Pitfalls, problems, and progress in bronchopulmonary dysplasia. Pediatrics 2009, 123, 1562–1573. [Google Scholar] [CrossRef]
- Hütten, M.C.; Kramer, B.W. Patterns and etiology of acute and chronic lung injury: Insights from experimental evidence. Zhongguo Dang Dai Er Ke Za Zhi 2014, 16, 448–459. [Google Scholar]
- Kallapur, S.G.; Moss, T.J.M.; Ikegami, M.; Jasman, R.L.; Newnham, J.P.; Jobe, A.H. Recruited inflammatory cells mediate endotoxin-induced lung maturation in preterm fetal lambs. Am. J. Respir. Crit. Care Med. 2005, 172, 1315–1321. [Google Scholar] [CrossRef]
- Smyth, J.A.; Ellis, W.A. Stillbirth/perinatal weak calf syndrome: Radiographic examination for growth retardation lines. Vet. Rec. 1996, 139, 599–600. [Google Scholar] [PubMed]
- Smyth, J.A.; Goodall, E.A.; McCoy, M.A.; Ellis, W.A. Stillbirth/perinatal weak calf syndrome: A study of calves with an abnormal thyroid gland. Vet. Rec. 1996, 139, 11–16. [Google Scholar] [CrossRef]
- Drozdowska, J.; Cousens, C.; Finlayson, J.; Collie, D.; Dagleish, M.P. Structural Development, Cellular Differentiation and Proliferation of the Respiratory Epithelium in the Bovine Fetal Lung. J. Comp. Pathol. 2016, 154, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Castleman, W.L.; Lay, J.C. Morphometric and ultrastructural study of postnatal lung growth and development in calves. Am. J. Vet. Res. 1990, 51, 789–795. [Google Scholar] [PubMed]
- Kuypers, E.; Collins, J.J.P.; Jellema, R.K.; Wolfs, T.G.A.M.; Kemp, M.W.; Nitsos, I.; Pillow, J.J.; Polglase, G.R.; Newnham, J.P.; Germeraad, W.T.V.; et al. Ovine Fetal Thymus Response to Lipopolysaccharide-Induced Chorioamnionitis and Antenatal Corticosteroids. PLoS ONE 2012, 7, e38257. [Google Scholar] [CrossRef]
- Kuypers, E.; Wolfs, T.G.A.M.; Collins, J.J.P.; Jellema, R.K.; Newnham, J.P.; Kemp, M.W.; Kallapur, S.G.; Jobe, A.H.; Kramer, B.W. Intraamniotic lipopolysaccharide exposure changes cell populations and structure of the ovine fetal thymus. Reprod. Sci. 2013, 20, 946–956. [Google Scholar] [CrossRef]
- Kunzmann, S.; Glogger, K.; Been, J.V.; Kallapur, S.G.; Nitsos, I.; Moss, T.J.; Speer, C.P.; Newnham, J.P.; Jobe, A.H.; Kramer, B.W. Thymic changes after chorioamnionitis induced by intraamniotic lipopolysaccharide in fetal sheep. Am. J. Obstet. Gynecol. 2010, 202, 476.e1–476.e9. [Google Scholar] [CrossRef]
- Cromi, A.; Ghezzi, F.; Raffaelli, R.; Bergamini, V.; Siesto, G.; Bolis, P. Ultrasonographic measurement of thymus size in IUGR fetuses: A marker of the fetal immunoendocrine response to malnutrition. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 2009, 33, 421–426. [Google Scholar] [CrossRef]
- Gasisova, A.I.; Atkenova, A.B.; Ahmetzhanova, N.B.; Murzabekova, L.M.; Bekenova, A.C. Morphostructure of Immune System Organs in Cattle of Different Age. Anat. Histol. Embryol. 2017, 46, 132–142. [Google Scholar] [CrossRef]
- Takasu, M.; Shirota, K.; Ohba, Y.; Nishii, N.; Murase, T.; Miyazawa, K.; Kitagawa, H. Thymic Hypoplasia in Japanese Black Calves with Stillbirth/Perinatal Weak Calf Syndrome. J. Vet. Med. Sci. 2008, 70, 1173–1177. [Google Scholar] [CrossRef]
- Knapek, K.J.; Georges, H.M.; Van Campen, H.; Bishop, J.V.; Bielefeldt-Ohmann, H.; Smirnova, N.P.; Hansen, T.R. Fetal Lymphoid Organ Immune Responses to Transient and Persistent Infection with Bovine Viral Diarrhea Virus. Viruses 2020, 12, 816. [Google Scholar] [CrossRef]
- Ohmann, H.B. Immunoglobulin levels in non-aborted and aborted fetuses from Danish herds of cattle. Acta Vet. Scand. 1981, 22, 428–434. [Google Scholar] [CrossRef]
- Bosch, J.C.; van Lieshout, J.A.H.; de Wit, J.J.; Graat, E.A.M.; Somers, M.J.M. The serological BHV1 status of dams determines the precolostral status of their calves. Vet. Q. 2000, 22, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Gabriël, S.; Geldhof, P.; Phiri, I.K.; Cornillie, P.; Goddeeris, B.M.; Vercruysse, J. Placental transfer of immunoglobulins in cattle infected with Schistosoma mattheei. Vet. Immunol. Immunopathol. 2005, 104, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Turin, L.; Invernizzi, P.; Woodcock, M.; Grati, F.R.; Riva, F.; Tribbioli, G.; Laible, G. Bovine fetal microchimerism in normal and embryo transfer pregnancies and its implications for biotechnology applications in cattle. Biotechnol. J. 2007, 2, 486–491. [Google Scholar] [CrossRef]
- Ivanoff, M.R. Weak calf syndrome: Serum immunoglobulin concentrations in precolostral calves. Am. J. Vet. Res. 1975, 36, 1129–1131. [Google Scholar] [PubMed]
- Jawor, P.; Stefaniak, T.; Mee, J.F. Immune and inflammatory biomarkers in cases of bovine perinatal mortality with and without infection in utero. J. Dairy Sci. 2017, 100. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Holsteg, M.; Schirrmeier, H.; Hoffmann, B.; Beer, M. Natural infection of pregnant cows with Schmallenberg virus—A follow-up study. PLoS ONE 2014, 9, 1–5. [Google Scholar] [CrossRef]
- Jawor, P.; Zabek, A.; Wojtowicz, W.; Król, D.; Stefaniak, T.; Młynarz, P. Metabolomic studies as a tool for determining the post-mortem interval (PMI) in stillborn calves. BMC Vet. Res. 2019, 15, 189. [Google Scholar] [CrossRef]
- Huang, M.Z.; Cui, D.A.; Wu, X.H.; Hui, W.; Yan, Z.T.; Ding, X.Z.; Wang, S.Y. Serum metabolomics revealed the differential metabolic pathway in calves with severe clinical diarrhea symptoms. Animals 2020, 10, 769. [Google Scholar] [CrossRef]
- Blakebrough-Hall, C.; Dona, A.; D’occhio, M.J.; McMeniman, J.; González, L.A. Diagnosis of Bovine Respiratory Disease in feedlot cattle using blood 1H NMR metabolomics. Sci. Rep. 2020, 10, 115. [Google Scholar] [CrossRef]
- Kornmatitsuk, B.; Dahl, E.; Ropstad, E.; Bechers, J.F.; Gustafsson, H.; Kindahl, H. Endocrine profiles, haematology and pregnancy outcomes of late pregnant Holstein dairy heifers sired by bulls giving a high or low incidence of stillbirth. Acta Vet. Scand. 2004, 45, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Kindahl, H.; Kornmatitsuk, B.; Königsson, K.; Gustafsson, H. Endocrine changes in late bovine pregnancy with special emphasis on fetal well-being. Domest. Anim. Endocrinol. 2002, 23, 321–328. [Google Scholar] [CrossRef]
- Sorge, U.S.; Kelton, D.F.; Staufenbiel, R. Short communication: Prepartal concentration of estradiol-17β in heifers with stillborn calves. J. Dairy Sci. 2008, 91, 1433–1437. [Google Scholar] [CrossRef] [PubMed]
- Uchida, E.; Katoh, N.; Takahashi, K. Appearance of haptoglobin in serum from cows at parturition. J. Vet. Med. Sci. 1993, 55, 893–894. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.J.; Romero, R.; Park, J.Y.; Kang, J.; Hong, J.-S.; Yoon, B.H. A high concentration of fetal fibronectin in cervical secretions increases the risk of intra-amniotic infection and inflammation in patients with preterm labor and intact membranes. J. Perinat. Med. 2019, 47, 288–303. [Google Scholar] [CrossRef]
- Lis-Kuberka, J.; Berghausen-Mazur, M.; Kątnik-Prastowska, I.; Orczyk-Pawiłowicz, M. Delivery-associated presence of supramolecular fibronectin-fibrin complexes in puerperal and cord plasma. J. Matern. Fetal. Neonatal Med. 2019, 32, 3581–3588. [Google Scholar] [CrossRef]
- Jawor, P.; Krzyżanowska-Gołąb, D.; Bajzert, J.; Stefaniak, T.; Kątnik-Prastowska, I. Changes of plasma fibronectin and fibronectin-fibrin complexes in dams of stillborn dairy calves. Ir. Vet. J. 2020, 73, 17. [Google Scholar] [CrossRef]
- Krzyzanowska-Goła̧b, D.; Lemańska-Perek, A.; Pupek, M.; Lindner, K.; Polańska, B.; Porȩbska, I.; Ka̧tnik-Prastowska, I. Identification of soluble supramolecular FN-fibrin complexes in human plasma. J. Immunoass. Immunochem. 2014, 35, 412–427. [Google Scholar] [CrossRef]
- Lemańska-Perek, A.; Polańska, B.; Krzyżanowska-Gołąb, D.; Kątnik-Prastowska, I. Occurrence of soluble supra-molecular FN–fibrin complexes in the plasma of children with recurrent respiratory infection. Ann. Clin. Biochem. 2015, 52, 441–447. [Google Scholar] [CrossRef]
- Pupek, M.; Krzyżanowska-Gołąb, D.; Kotschy, D.; Witkiewicz, W.; Kwiatkowska, W.; Kotschy, M.; Kątnik-Prastowska, I. Time-dependent changes in extra-domain A-fibronectin concentration and relative amounts of fibronectin-fibrin complexes in plasma of patients with peripheral arterial disease after endovascular revascularisation. Int. Wound J. 2018. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jawor, P.; Mee, J.F.; Stefaniak, T. Role of Infection and Immunity in Bovine Perinatal Mortality: Part 2. Fetomaternal Response to Infection and Novel Diagnostic Perspectives. Animals 2021, 11, 2102. https://doi.org/10.3390/ani11072102
Jawor P, Mee JF, Stefaniak T. Role of Infection and Immunity in Bovine Perinatal Mortality: Part 2. Fetomaternal Response to Infection and Novel Diagnostic Perspectives. Animals. 2021; 11(7):2102. https://doi.org/10.3390/ani11072102
Chicago/Turabian StyleJawor, Paulina, John F. Mee, and Tadeusz Stefaniak. 2021. "Role of Infection and Immunity in Bovine Perinatal Mortality: Part 2. Fetomaternal Response to Infection and Novel Diagnostic Perspectives" Animals 11, no. 7: 2102. https://doi.org/10.3390/ani11072102
APA StyleJawor, P., Mee, J. F., & Stefaniak, T. (2021). Role of Infection and Immunity in Bovine Perinatal Mortality: Part 2. Fetomaternal Response to Infection and Novel Diagnostic Perspectives. Animals, 11(7), 2102. https://doi.org/10.3390/ani11072102