
Effect of Substituting Fish Oil with Camelina Oil on Growth Performance, Fatty Acid Profile, Digestibility, Liver Histology, and Antioxidative Status of Red Seabream (Pagrus major)

 ,
,  , , ,
, , ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Husbandry
2.3. Performance Variables and Biometric Indices
2.4. Proximate Composition Analysis, Fatty Acid and Digestibility Assessment
2.5. Blood Hematological Parameters
2.6. Antioxidants Activity
2.7. Hepatic Histopathological Assessment
2.8. Statistical Analysis
3. Results
3.1. Growth Performance Variables
3.2. Proximate Composition of Fish Whole Body
3.3. Apparent Nutrient Digestibility
3.4. Blood Chemical Parameter
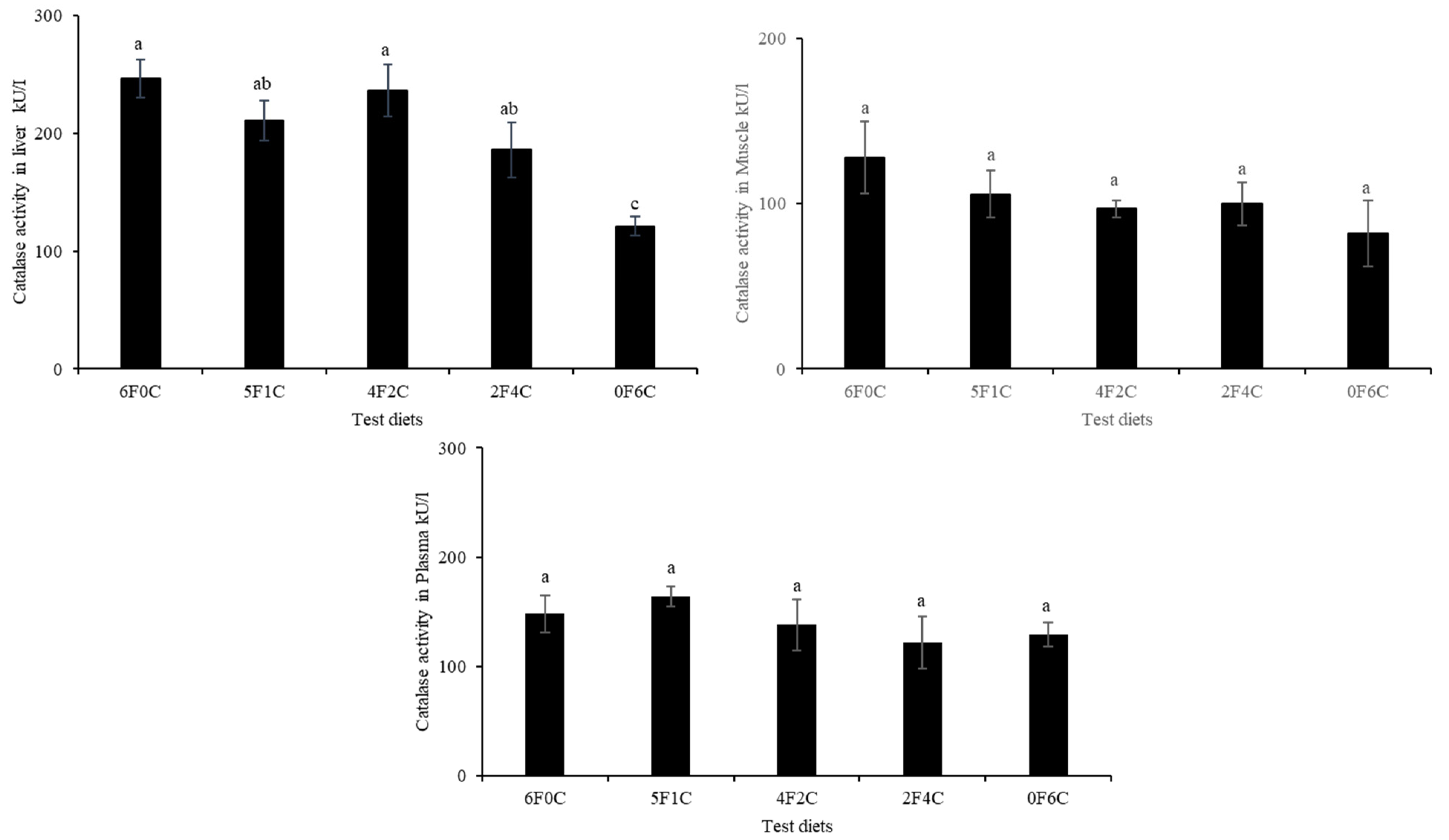
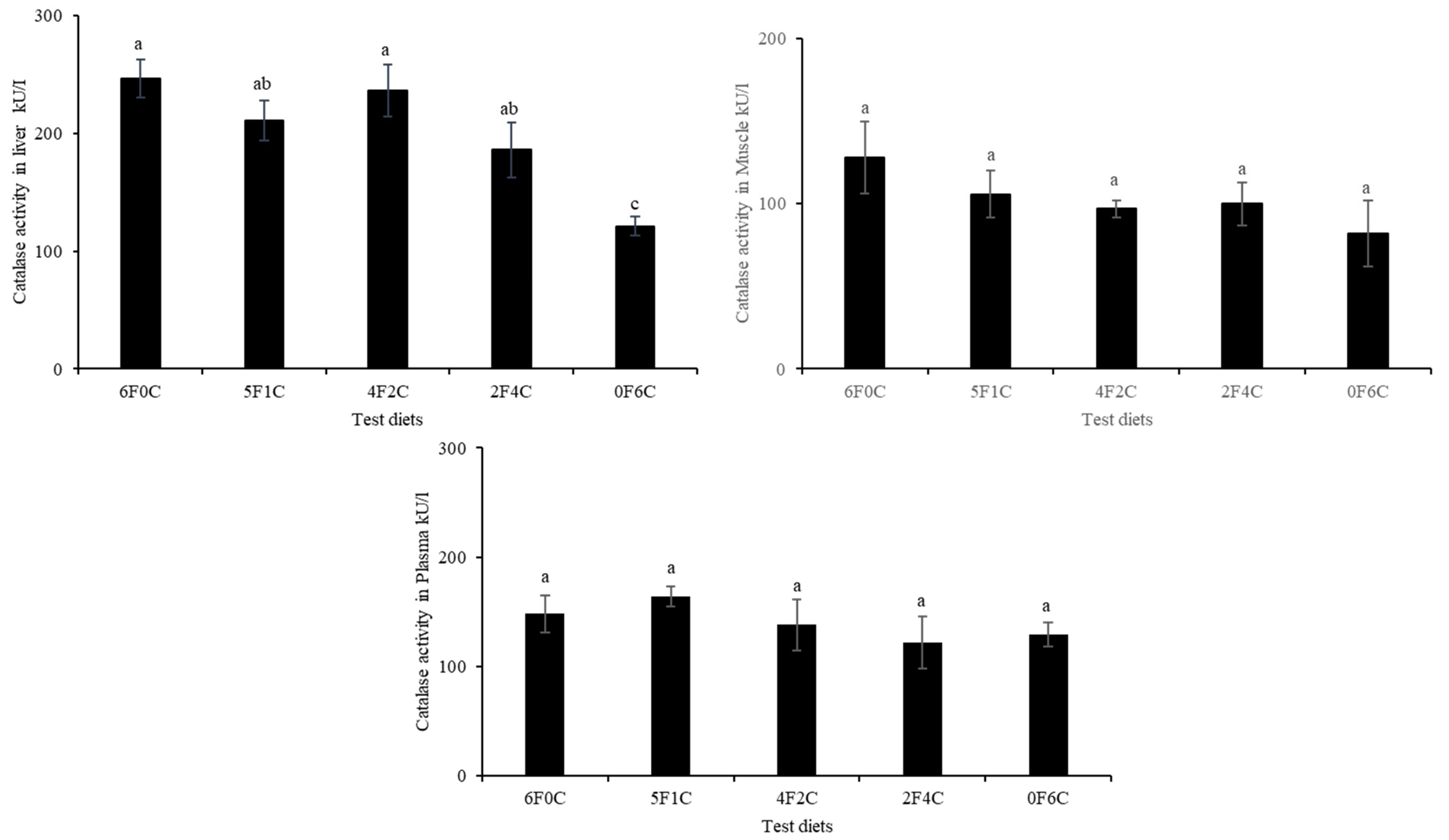
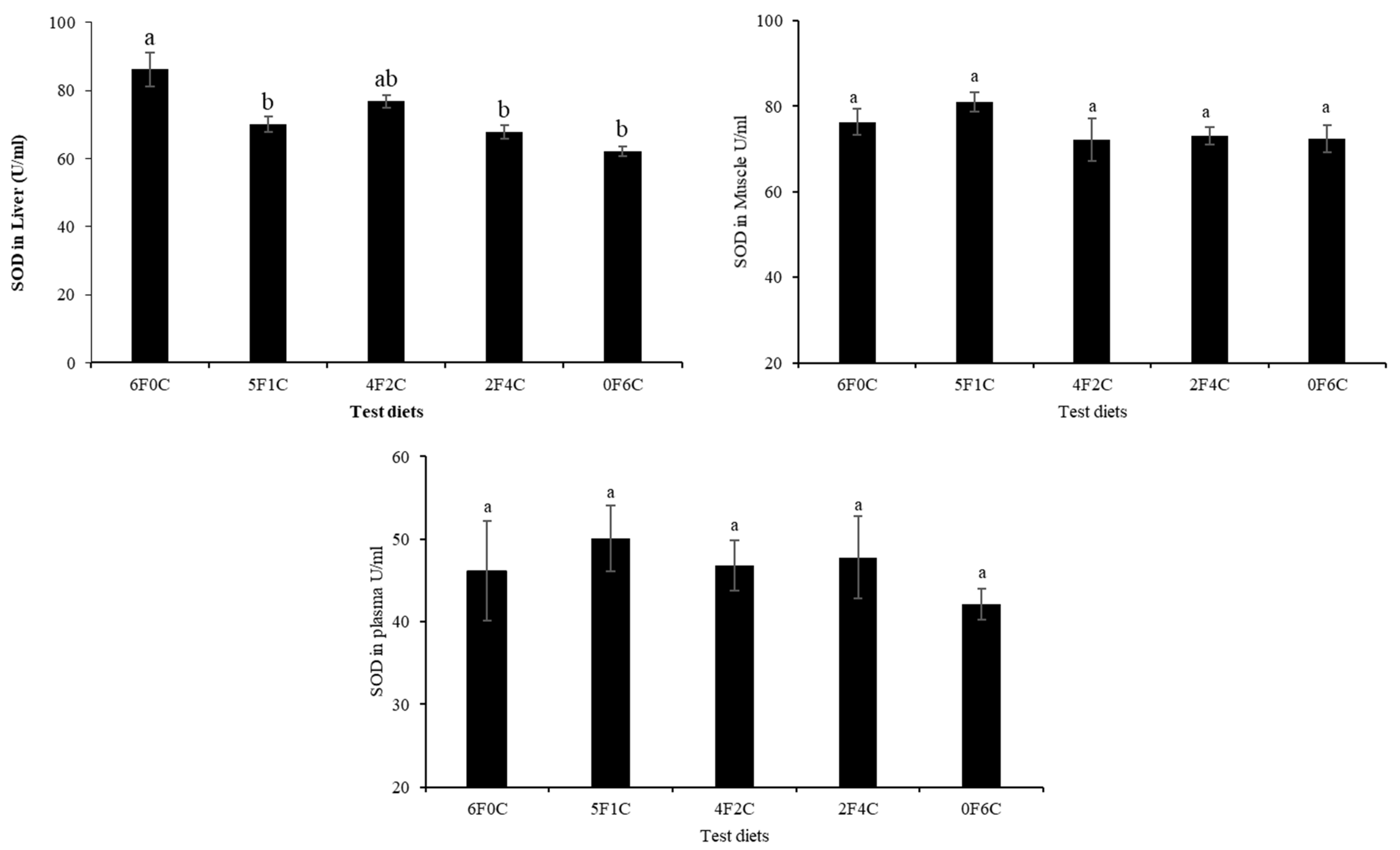
3.5. Antioxidants Capacity
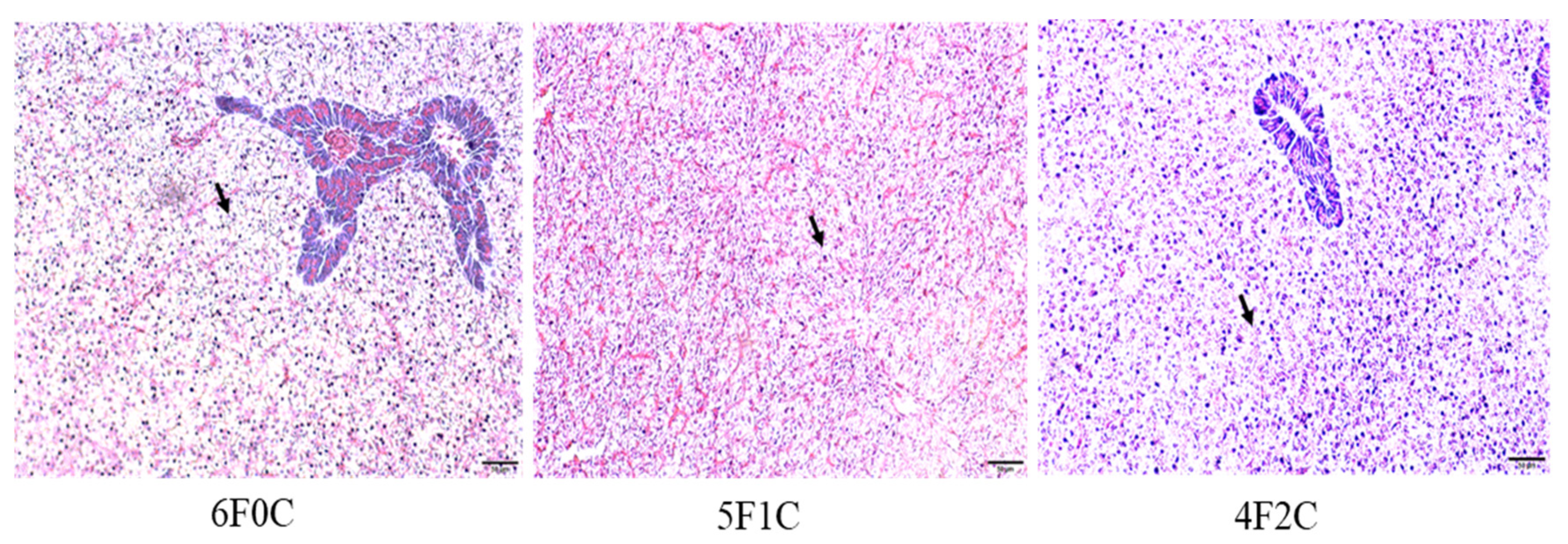
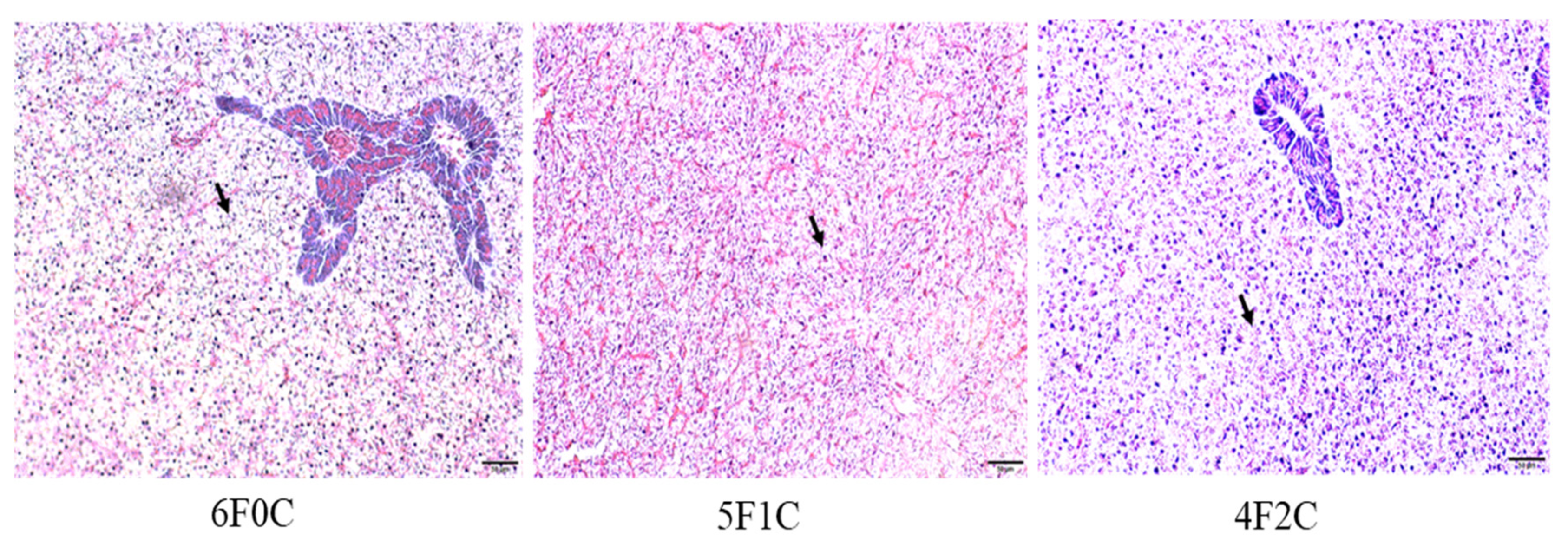
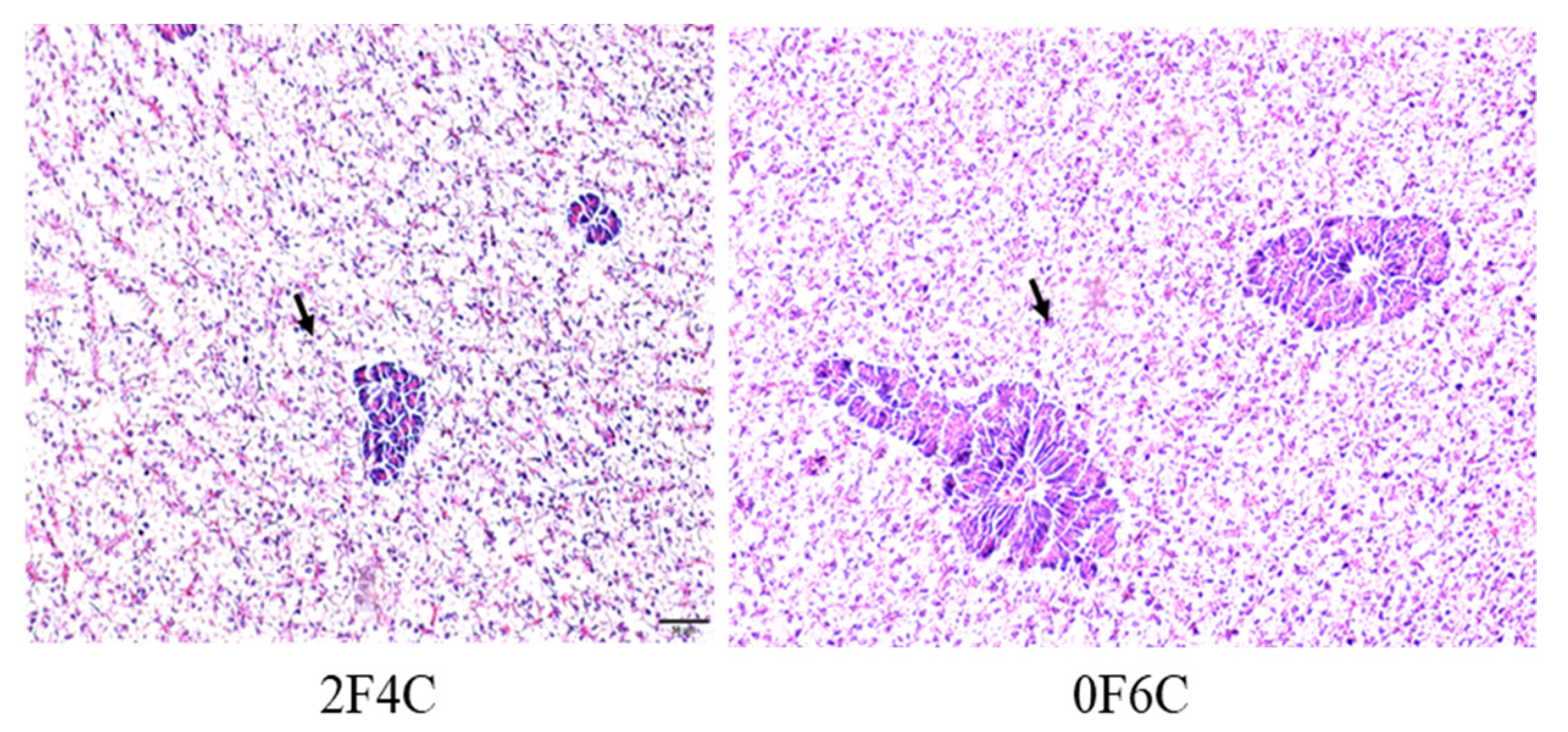
3.6. Hepatic Histological Examination
4. Discussion
4.1. Growth Performance and Nutrient Utilization
4.2. Fatty Acid Digestibility
4.3. Blood Chemistry
4.4. Lipid Peroxidation
4.5. Histomorphology of Liver
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fenton, J.I.; Hord, N.G.; Ghosh, S.; Gurzell, E.A. Immunomodulation by dietary long chain omega-3 fatty acids and the potential for adverse health outcomes. Prostaglandins Leukot. Essent. Fatty Acids 2013, 89, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Emery, J.A.; Norambuena, F.; Trushenski, J.; Turchini, G.M. Uncoupling EPA and DHA in fish nutrition: Dietary demand is limited in Atlantic salmon and effectively met by DHA alone. Lipids 2016, 51, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.-K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Dawood, M.A.O. Nutritional immunity of fish intestines: Important insights for sustainable aquaculture. Rev. Aquac. 2021, 13, 642–663. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Ali, M.F.; Amer, A.A.; Gewaily, M.S.; Mahmoud, M.M.; Alkafafy, M.; Assar, D.H.; Soliman, A.A.; Van Doan, H. The influence of coconut oil on the growth, immune, and antioxidative responses and the intestinal digestive enzymes and histomorphometry features of Nile tilapia (Oreochromis niloticus). Fish Physiol. Biochem. 2021, 1–12. [Google Scholar] [CrossRef]
- Betancor, M.; Sprague, M.; Usher, S.; Sayanova, O.; Campbell, P.; Napier, J.A.; Tocher, D.R. A nutritionally-enhanced oil from transgenic Camelina sativa effectively replaces fish oil as a source of eicosapentaenoic acid for fish. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budin, J.T.; Breene, W.M.; Putnam, D.H. Some compositional properties of camelina (Camelina sativa l. Crantz) seeds and oils. J. Am. Oil Chem. Soc. 1995, 72, 309–315. [Google Scholar] [CrossRef]
- Lu, J.; Tibbetts, S.M.; Lall, S.P.; Anderson, D.M. Use of dietary oil, solvent-extracted meal and protein concentrate from camelina sativa for rainbow trout, Oncorhynchus mykiss, at the early fry stage. Aquaculture 2020, 524, 735252. [Google Scholar] [CrossRef]
- Huyben, D.; Rimoldi, S.; Ceccotti, C.; Montero, D.; Betancor, M.; Iannini, F.; Terova, G. Effect of dietary oil from Camelina sativa on the growth performance, fillet fatty acid profile and gut microbiome of gilthead sea bream (Sparus aurata). PeerJ 2020, 8, e10430. [Google Scholar] [CrossRef]
- Betancor, M.B.; MacEwan, A.; Sprague, M.; Gong, X.; Montero, D.; Han, L.; Napier, J.A.; Norambuena, F.; Izquierdo, M.; Tocher, D.R. Oil from transgenic Camelina sativa as a source of EPA and DHA in feed for European sea bass (Dicentrarchus labrax L.). Aquaculture 2021, 530, 735759. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Effect of replacement of fish oil with camelina (Camelina sativa) oil on growth, lipid class and fatty acid composition of farmed juvenile Atlantic cod (Gadus morhua). Fish Physiol. Biochem. 2013, 39, 1441–1456. [Google Scholar] [CrossRef] [PubMed]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Changes in tissue lipid and fatty acid composition of farmed rainbow trout in response to dietary camelina oil as a replacement of fish oil. Lipids 2014, 49, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Full substitution of fish oil with camelina (Camelina sativa) oil, with partial substitution of fish meal with camelina meal, in diets for farmed Atlantic salmon (Salmo salar) and its effect on tissue lipids and sensory quality. Food Chem. 2014, 157, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Betancor, M.B.; Sprague, M.; Sayanova, O.; Usher, S.; Metochis, C.; Campbell, P.J.; Napier, J.A.; Tocher, D.R. Nutritional evaluation of an EPA-DHA oil from transgenic Camelina sativa in feeds for post-smolt Atlantic salmon (Salmo salar L.). PLoS ONE 2016, 11, e0159934. [Google Scholar] [CrossRef]
- Toyes-Vargas, E.A.; Parrish, C.C.; Viana, M.T.; Carreón-Palau, L.; Magallón-Servín, P.; Magallón-Barajas, F.J. Replacement of fish oil with camelina (Camelina sativa) oil in diets for juvenile tilapia (var. GIFT Oreochromis niloticus) and its effect on growth, feed utilization and muscle lipid composition. Aquaculture 2020, 523, 735177. [Google Scholar] [CrossRef]
- Eidhin, D.N.; Burke, J.; O’Beirne, D. Oxidative stability of ω3-rich camelina oil and camelina oil-based spread compared with plant and fish oils and sunflower spread. J. Food Sci. 2003, 68, 345–353. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S. Effects of dietary inactivated Pediococcus pentosaceus on growth performance, feed utilization and blood characteristics of red sea bream, Pagrus major juvenile. Aquac. Nutr. 2016, 22, 923–932. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S. Immune responses and stress resistance in red sea bream, Pagrus major, after oral administration of heat-killed Lactobacillus plantarum and vitamin C. Fish Shellfish Immunol. 2016, 54, 266–275. [Google Scholar] [CrossRef]
- Seong, T.; Matsutani, H.; Haga, Y.; Kitagima, R.; Satoh, S. First step of non-fish meal, non-fish oil diet development for red seabream, (Pagrus major), with plant protein sources and microalgae Schizochytrium sp. Aquac. Res. 2019, 50, 2460–2468. [Google Scholar] [CrossRef]
- Seong, T.; Kitagima, R.; Haga, Y.; Satoh, S. Non-fish meal, non-fish oil diet development for red sea bream, Pagrus major, with plant protein and graded levels of Schizochytrium sp.: Effect on growth and fatty acid composition. Aquac. Nutr. 2020, 26, 1173–1185. [Google Scholar] [CrossRef]
- Komilus, C.F.; Shichi, N.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Michael, F.R.; Gao, J.; Makita, C. Influences of palm oil blended with fish oil on growth performances and lipid profiles of red seabream Pagrus major. Aquac. Sci. 2008, 56, 317–326. [Google Scholar]
- AOAC. Association of official analytical chemists. In Official Methods of Analysis of Official Analytical Chemists International, 16th ed.; AOAC: Washington, DC, USA, 1998. [Google Scholar]
- Ackman, R.G. Remarks on official methods employing boron trifluoride in the preparation of methyl esters of the fatty acids of fish oils. J. Am. Oil Chem. Soc. 1998, 75, 541–545. [Google Scholar] [CrossRef]
- Sigurgisladottir, S.; Lall, S.P.; Parrish, C.C.; Ackman, R.G. Cholestane as a digestibility marker in the absorption of polyunsaturated fatty acid ethyl esters in atlantic salmon. Lipids 1992, 27, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.Y.; Oo, A.N.; Higgs, D.A.; Brauner, C.J.; Satoh, S. Effect of dietary canola oil level on the growth performance and fatty acid composition of juvenile red sea bream, Pagrus major. Aquaculture 2007, 271, 420–431. [Google Scholar] [CrossRef]
- Koshio, S. Red sea bream, Pagrus major. Nutr. Requir. Feed. Finfish Aquac. 2002, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Yang, Z.; Cheng, Y.; Wang, J.; Zhou, J. Effects of the complete replacement of fish oil with linseed oil on growth, fatty acid composition, and protein expression in the Chinese mitten crab (Eriocheir sinensis). Proteome Sci. 2018, 16, 1–11. [Google Scholar] [CrossRef]
- Bell, J.G.; Strachan, F.; Good, J.E.; Tocher, D.R. Effect of dietary echium oil on growth, fatty acid composition and metabolism, gill prostaglandin production and macrophage activity in atlantic cod (Gadus morhua L.). Aquac. Res. 2006, 37, 606–617. [Google Scholar] [CrossRef]
- Osmond, A.T.Y.; Arts, M.T.; Bazinet, R.P.; Napier, J.A.; Han, L.; Colombo, S.M. Transgenic camelina oil is an effective source of eicosapentaenoic acid and docosahexaenoic acid in diets for farmed rainbow trout, in terms of growth, tissue fatty acid content, and fillet sensory properties. J. World Aquac. Soc. 2021. [Google Scholar] [CrossRef]
- Ishikawa, M.; Teshima, S.I.; Kanazawa, A.; Koshio, S. Measurements of digestibilities of cholesterol and fatty acids using 5α-cholestane as an inert marker in the tilapia, Oreochromis niloticus, and the freshwater prawn, Macrobrachium rosenbergii. J. Appl. Ichthyol. 1997, 13, 31–35. [Google Scholar] [CrossRef]
- Ye, C.L.; Anderson, D.M.; Lall, S.P. The effects of camelina oil and solvent extracted camelina meal on the growth, carcass composition and hindgut histology of Atlantic salmon (Salmo salar) parr in freshwater. Aquaculture 2016, 450, 397–404. [Google Scholar] [CrossRef]
- Morais, S.; Koven, W.; Rønnestad, I.; Teresa Dinis, M.; Conceição, L.E.C. Dietary protein: Lipid ratio and lipid nature affects fatty acid absorption and metabolism in a teleost larva. Br. J. Nutr. 2005, 93, 813–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchini, G.M.; Mentasti, T.; Caprino, F.; Giani, I.; Panseri, S.; Bellagamba, F.; Moretti, V.M.; Valfré, F. The relative absorption of fatty acids in brown trout (Salmo trutta) fed a commercial extruded pellet coated with different lipid sources. Ital. J. Anim. Sci. 2005, 4, 241–252. [Google Scholar] [CrossRef]
- Dossou, S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Dawood, M.A.O.; El Basuini, M.F.; Olivier, A.; Zaineldin, A.I. Growth performance, blood health, antioxidant status and immune response in red seabream (Pagrus major) fed Aspergillus oryzae fermented rapeseed meal (rm-koji). Fish Shellfish Immunol. 2018, 75, 253–262. [Google Scholar] [CrossRef]
- El Basuini, M.F.; El-Hais, A.M.; Dawood, M.A.O.; Abou-Zeid, A.E.-S.; El-Damrawy, S.Z.; Khalafalla, M.M.E.L.S.; Koshio, S.; Ishikawa, M.; Dossou, S. Effect of different levels of dietary copper nanoparticles and copper sulfate on growth performance, blood biochemical profiles, antioxidant status and immune response of red seabream (Pagrus major). Aquaculture 2016, 455, 32–40. [Google Scholar] [CrossRef]
- Arab-Tehrany, E.; Jacquot, M.; Gaiani, C.; Imran, M.; Desobry, S.; Linder, M. Beneficial effects and oxidative stability of omega-3 long-chain polyunsaturated fatty acids. Trends Food Sci. Technol. 2012, 25, 24–33. [Google Scholar] [CrossRef]
- Jin, M.; Yuan, Y.; Lu, Y.; Ma, H.; Sun, P.; Li, Y.; Qiu, H.; Ding, L.; Zhou, Q. Regulation of growth, tissue fatty acid composition, biochemical parameters and lipid related genes expression by different dietary lipid sources in juvenile black seabream, Acanthopagrus schlegelii. Aquaculture 2017, 479, 25–37. [Google Scholar] [CrossRef]
- Zuo, R.; Ai, Q.; Mai, K.; Xu, W. Effects of conjugated linoleic acid on growth, non-specific immunity, antioxidant capacity, lipid deposition and related gene expression in juvenile large yellow croaker (Larmichthys crocea) fed soyabean oil-based diets. Br. J. Nutr. 2013, 110, 1220–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radovanović, T.B.; Borković-Mitić, S.S.; Perendija, B.R.; Despotović, S.G.; Pavlović, S.; Cakić, P.; Saičić, Z.S. Superoxide dismutase and catalase activities in the liver and muscle of barbel (Barbus barbus) and its intestinal parasite (Pomphoryinchus laevis) from the danube river, serbia. Arch. Biol. Sci. 2010, 62, 97–105. [Google Scholar] [CrossRef]
- Shourbela, R.M.; El-Hawarry, W.N.; Elfadadny, M.R.; Dawood, M.A.O. Oregano essential oil enhanced the growth performance, immunity, and antioxidative status of Nile tilapia (Oreochromis niloticus) reared under intensive systems. Aquaculture 2021, 542, 736868. [Google Scholar] [CrossRef]
- Abdel-Warith, A.-W.A.; Younis, E.M.; Al-Asgah, N.A.; Gewaily, M.S.; El-Tonoby, S.M.; Dawood, M.A.O. Role of fucoidan on the growth behavior and blood metabolites and toxic effects of atrazine in Nile tilapia Oreochromis niloticus (linnaeus, 1758). Animals 2021, 11, 1448. [Google Scholar] [CrossRef]
- Kohen, R.; Nyska, A. Invited review: Oxidation of biological systems: Oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [Green Version]
- Dawood, M.A.O.; Noreldin, A.E.; Sewilam, H. Long term salinity disrupts the hepatic function, intestinal health, and gills antioxidative status in Nile tilapia stressed with hypoxia. Ecotoxicol. Environ. Saf. 2021, 220, 112412. [Google Scholar] [CrossRef]
- Abele, D.; Puntarulo, S. Formation of reactive species and induction of antioxidant defence systems in polar and temperate marine invertebrates and fish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2004, 138, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Mourente, G.; Bell, J.G.; Tocher, D.R. Does dietary tocopherol level affect fatty acid metabolism in fish? Fish Physiol. Biochem. 2007, 33, 269–280. [Google Scholar] [CrossRef]
- Iheanacho, S.C.; Odo, G.E. Neurotoxicity, oxidative stress biomarkers and haematological responses in African catfish (Clarias gariepinus) exposed to polyvinyl chloride microparticles. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 232, 108741. [Google Scholar] [CrossRef] [PubMed]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Noreldin, A.E.; Ali, M.A.M.; Sewilam, H. Menthol essential oil is a practical choice for intensifying the production of Nile tilapia (Oreochromis niloticus): Effects on the growth and health performances. Aquaculture 2021, 543, 737027. [Google Scholar] [CrossRef]
- Bhagat, J.; Ingole, B.; Singh, N. Glutathione s-transferase, catalase, superoxide dismutase, glutathione peroxidase, and lipid peroxidation as biomarkers of oxidative stress in snails: A review. Invertebr. Surviv. J. 2016, 13, 336–349. [Google Scholar]
- Yu, H.; Gao, Q.; Dong, S.; Lan, Y.; Ye, Z.; Wen, B. Regulation of dietary glutamine on the growth, intestinal function, immunity and antioxidant capacity of sea cucumber Apostichopus japonicus (selenka). Fish Shellfish Immunol. 2016, 50, 56–65. [Google Scholar] [CrossRef]
- Long, X.; Wu, R.; Wu, X.; Hou, W.; Pan, G.; Zeng, C.; Cheng, Y. Effects of dietary fish oil replacement with blended vegetable oils on growth, lipid metabolism and antioxidant capacity of subadult swimming crab Portunus trituberculatus. Aquac. Nutr. 2019, 25, 1218–1230. [Google Scholar] [CrossRef]
- El Asely, A.M.; Reda, R.M.; Salah, A.S.; Mahmoud, M.A.; Dawood, M.A.O. Overall performances of Nile tilapia (Oreochromis niloticus) associated with using vegetable oil sources under suboptimal temperature. Aquac. Nutr. 2020, 26, 1154–1163. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S. Interaction effects of dietary supplementation of heat-killed Lactobacillus plantarum and β-glucan on growth performance, digestibility and immune response of juvenile red sea bream, Pagrus major. Fish Shellfish. Immunol. 2015, 45, 33–42. [Google Scholar] [CrossRef]
- Caballero, M.J.; Obach, A.; Rosenlund, G.; Montero, D.; Gisvold, M.; Izquierdo, M.S. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss. Aquaculture 2002, 214, 253–271. [Google Scholar] [CrossRef]
- Bell, J.G.; Tocher, D.R.; MacDonald, F.M.; Sargent, J.R. Effects of dietary borage oil [enriched in γ-linolenic acid, 18: 3 (n-6)] or marine fish oil [enriched in eicosapentaenoic acid, 20: 5 (n-3)] on growth, mortalities, liver histopathology and lipid composition of juvenile turbot (Scophthalmus maximus). Fish Physiol. Biochem. 1995, 14, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.E.; Myklebust, R.; Ringø, E.; Mayhew, T.M. The influences of dietary linseed oil and saturated fatty acids on caecal enterocytes in arctic char (Salvelinus alpinus L.): A quantitative ultrastructural study. Fish Physiol. Biochem. 2000, 22, 207–216. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient, g/kg DM | Test Diets | ||||
|---|---|---|---|---|---|
| 6F0C | 5F1C | 4F2C | 2F4C | 0F6C | |
| Brown fish meal a | 460 | 460 | 460 | 460 | 460 |
| Soybean meal b | 205 | 205 | 205 | 205 | 205 |
| Fish oil c | 60 | 50 | 40 | 20 | - |
| Camelina oil e | - | 10 | 20 | 40 | 60 |
| Soybean Lecithin d | 30 | 30 | 30 | 30 | 30 |
| EPA f | 50 | 50 | 50 | 50 | 50 |
| DHA g | 50 | 50 | 50 | 50 | 50 |
| Dextrin h | 50 | 50 | 50 | 50 | 50 |
| Activated gluten i | 80 | 80 | 80 | 80 | 80 |
| Mineral mix j | 40 | 40 | 40 | 40 | 40 |
| Vitamin mix k | 40 | 40 | 40 | 40 | 40 |
| Stay C l | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
| α-Cellulose m | 24.5 | 24.5 | 24.5 | 24.5 | 24.5 |
| Proximate composition | |||||
| Crude Protein | 498 ± 3 | 513 ± 4 | 508 ± 5 | 506 ± 0 | 508 ± 9 |
| Crude Lipid | 150 ± 9 | 144 ± 8 | 143 ± 13 | 135 ± 2 | 144 ± 10 |
| Moisture | 101 ± 1 | 93 ± 1 | 100 ± 4 | 90 ± 00 | 91 ± 5 |
| Ash | 101 ± 1 | 106 ± 00 | 101 ± 3 | 104 ± 3 | 103 ± 5 |
| Energy (kJ/g) n | 2580 ± 6 | 2612 ± 14 | 2603 ± 14 | 2590 ± 4 | 2591 ± 2 |
| Fatty Acid Type | Test Diet | ||||
|---|---|---|---|---|---|
| 6F0C | 5F1C | 4F2C | 2F4C | 0F6C | |
| 14:0 | 14.0 ± 1.8 | 13.8 ± 0.6 | 14.3 ± 0.3 | 11.2 ± 0.2 | 7.2 ± 0.2 |
| 16:0 | 51.1 ± 0.9 | 64.7 ± 0.6 | 86.0 ± 3.0 | 95.8 ± 50.0 | 114.0 ± 14.5 |
| 18:0 | 82.0 ± 2.0 | 35.0 ± 0.7 | 43.9 ± 3.9 | 37.4 ± 0.5 | 23.8 ± 4.3 |
| ∑Saturated | 147.1 ± 3.6 | 132.5 ± 1.4 | 144.2 ± 2.1 | 144.4 ± 5.4 | 145 ± 4.1 |
| 16:1n-9 | 55 ± 1.8 | 50.8 ± 0.6 | 54.2 ± 0.7 | 32.9 ± 1.5 | 31.1 ± 0.1 |
| 18:1n-5 | 1.7 ± 0.5 | 1.1 ± 0.9 | 1.7 ± 0.1 | 2.7 ± 0.2 | 2.2 ± 0.6 |
| 18:1n-9 | 64.3 ± 1.2 | 86.1 ± 3.1 | 120.4 ± 2.0 | 137.1 ± 1.3 | 155.2 ± 2.6 |
| 20:1n-9 | 20.8 ± 4.0 | 17.0 ± 3.5 | 20.5 ± 1.3 | 11.0 ± 0.1 | 9.5 ± 0.3 |
| 22:1n-9 | 33.5 ± 3.5 | 17.8 ± 0.2 | 8.9 ± 0.4 | 7.9 ± 1.6 | 2.7 ± 1.5 |
| ∑ MUFA | 175.5 ± 0.9 | 172.8 ± 5.8 | 205.7 ± 4.1 | 191.6 ± 6.5 | 200.7 ± 2.8 |
| 18:2n-6 | 41.1 ± 3.5a | 52.7 ± 4.1 | 72.5 ± 3.6 | 84.8 ± 0. | 103.4 ± 11.0 |
| 18:3n-6 | 6.4 ± 0.2 | 2.2 ± 0.1 | 1.6 ± 0.2 | 1.3 ± 0.1 | 0.4 ± 0.0 |
| 20:4n-6 | 8.2 ± 6.2 | 7.8 ± 4.6 | 6.4 ± 0.4 | 3.8 ± 0.2 | 2.7 ± 0.5 |
| 22:4n-6 | 2.1 ± 0.2 | 1.7 ± 0.1 | 1.4 ± 0.1 | 0.5 ± 0.0 | 0.0 ± 0.0 |
| ∑ n-6 fatty acids | 57.8 ± 8.3 | 64.4 ± 2.5 | 81.9 ± 7.6 | 90.4 ± 1.6 | 106.5 ± 1.6 |
| 18:3n-3 | 8.7 ± 1.6 | 10.0 ± 0.0 | 10.7 ± 0.7 | 12.3 ± 0.1 | 14.1 ± 8.8 |
| 18:4n-3 | 6.5 ± 0.2 | 3.1 ± 0.1 | 2.2 ± 0.1 | 1.8 ± 0.3 | 0.0 ± 0.0 |
| 20:3n-3 | 2.4 ± 0.5 | 1.4 ± 0.1 | 1.2 ± 0.1 | 0.5 ± 0.5 | 0.2 ± 0.2 |
| 20:4n-3 | 10.5 ± 0.2 | 6.5 ± 0.1 | 3.3 ± 0.3 | 3.5 ± 0.1 | 1.5 ± 0.4 |
| 20:5n-3 | 58.1 ± 20.2 | 57.0 ± 20.0 | 31.0 ± 2.0 | 21.3 ± 1.8 | 17.1 ± 1.8 |
| 22:5n-3 | 18.5 ± 4.5 | 14.2 ± 1.0 | 10.2 ± 0.7 | 9.0 ± 1.9 | 7.3 ± 2.0 |
| 22:6n-3 | 144.9 ± 23.9 | 129.0 ± 30.0 | 115.7 ± 11.1 | 101.0 ± 1.5 | 107.5 ± 2.5 |
| ∑ n-3 fatty acids | 249.6 ± 0.2 | 221.2 ± 2.6 | 174.3 ± 6.1 | 149.4 ± 2.3 | 146.5 ± 0.5 |
| ∑P UFA | 307.4 ± 12.4 | 285.6 ± 21.0 | 256.2 ± 19.2 | 239.8 ± 21.7 | 253. ± 46.2 |
| ∑-3HUFA | 234.4 ± 1.7 | 208.1 ± 16.2 | 161.4 ± 8.7 | 139.3 ± 0.4 | 133.6 ± 0.4 |
| ∑ n-3/n-6 ratio | 4.3 | 3.4 | 2.1 | 1.7 | 1.4 |
| ∑ EPA+DHA | 203 ± 21.1 | 186.1 ± 11.4 | 146.7 ± 2.1 | 122.3 ± 2.5 | 124.6 ± 1.2 |
| Parameters | Test Diets | ||||
|---|---|---|---|---|---|
| 6F0C | 5F1C | 4F2C | 2F4C | 0F6C | |
| BW0 (g/fish) | 1.8 | 1.9 | 1.8 | 1.9 | 1.9 |
| BW56d (g/fish) | 26.2 ± 0.1 | 27.1 ± 0.2 | 27.3 ± 0.6 | 28.3 ± 0.1 | 26.5 ± 0.1 |
| BWG (%) | 1306.2 ± 47.9 | 1326.3 ± 6.1 | 1364.4 ± 27.2 | 1365.3 ± 49.1 | 1275.5 ± 25.4 |
| SGR | 4.7 ± 0.1 | 4.7 ± 0.0 | 4.7 ± 0.0 | 4.7 ± 0.1 | 4.6 ± 0.0 |
| FI | 25.6 ± 0.2 | 26.7 ± 0.4 | 27.8 ± 1.2 | 27.0 ± 0.8 | 27.2 ± 1.1 |
| FCR | 1.1 ± 0.0 | 1.1 ± 0.0 | 1.1 ± 0.1 | 1.0 ± 0.0 | 1.1 ± 0.0 |
| HSI | 1.1 ± 0.1 | 1.0 ± 0.1 | 0.9 ± 0.1 | 1.1 ± 0.4 | 1.1 ± 0.1 |
| VSI | 9.1 ± 0.7 | 9.5 ± 0.3 | 7.5 ± 0.6 | 8.5 ± 0.7 | 8.9 ± 0.7 |
| CF | 2.3 ± 0.0 | 2.3 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.0 | 2.5 ± 0.2 |
| SR | 82.2 ± 5.8 | 97.7 ± 2.2 | 88.8 ± 8.0 | 100 ± 0.0 | 86.6 ± 6.62 |
| Parameter | Test Diets | ||||
|---|---|---|---|---|---|
| FO6CO0 | FO5CO1 | FO4CO2 | FO2CO4 | FO0CO6 | |
| Moisture | 704 ± 4.8 | 694 ± 4.5 | 702 ± 11.8 | 697 ± 6.9 | 700 ± 4.7 |
| Crude Protein | 546 ± 6 | 512 ± 00 | 544 ± 45 | 539 ± 4 | 531 ± 2 |
| Crude Lipid | 291 ± 3 a | 232 ± 1 b | 212 ± 4 bc | 232 ± 1 b | 244 ± 9 b |
| Ash | 157 ± 2 | 154 ± 3 | 161 ± 2 | 155 ± 1 | 155 ± 1 |
| Fatty Acid Type | Test Diet | ||||
|---|---|---|---|---|---|
| 6F0C | 5F1C | 4F2C | 2F4C | 0F6C | |
| 14:0 | 27.1 ± 0.3 | 29.1 ± 0.2 | 33.5 ± 0.6 | 32.0 ± 0.1 | 26.1 ± 0.2 |
| 16:0 | 123.2 ± 0.1 a | 109.5 ± 0.1 a | 102.5 ± 0.7 b | 96.0 ± 0.3 b | 85.9 ± 0.1 b |
| 18:0 | 67.5 ± 0.0 a | 57.5 ± 0.2 a | 46.1 ± 0.6 ab | 36.0 ± 0.6 bc | 26.3 ± 0.5 bc |
| ∑Saturated | 217.8 ± 0.0 a | 195.6 ± 0.3 a | 182.1 ± 0.3 ab | 164.0 ± 0.9 b | 138.3 ± 0.1 bc |
| 16:1n-9 | 61.5 ± 0.5 a | 44.8 ± 0.2 a | 37.7 ± 0.9 a | 32.9 ± 0.7 ab | 27.0 ± 0.7 b |
| 18:1n-5 | 4.9 ± 0.2 a | 2.0 ± 0.1 a | 3.0 ± 0.1 a | 1.7 ± 0.01 b | 0.0 ± 0.7 c |
| 18:1n-9 | 70.1 ± 0.01 a | 98.3 ± 0.5 b | 124.6 ± 0.8 c | 148.4 ± 0.1 c | 168.6 ± 1.0 c |
| 20:1n-9 | 51.0 ± 0.2 a | 44.2 ± 0.2 a | 36.8 ± 0.5 b | 37.5 ± 0.0 b | 31.2 ± 2.1 c |
| 22:1n-9 | 34.5 ± 0.3 a | 31.0 ± 0.01 a | 23.0 ± 0.0 a | 20.6 ± 0.5 ab | 13.6 ± 0.1 b |
| ∑MUFA | 222.0 ± 0.6 | 219.5 ± 0.1 | 223.1 ± 0.2 | 241.1 ± 0.6 | 240.2 ± 2.1 |
| 18:2n-6 | 84.0 ± 0.1 | 86.1 ± 0.2 | 99.2 ± 2.7 | 108.9 ± 1.5 | 113.2 ± 4.6 |
| 18:3n-6 | 5.8 ± 0.2 a | 3.7 ± 0.1 a | 3.1 ± 0.2 a | 2.5 ± 0.8 ab | 1.5 ± 0.0 b |
| 20:4n-6 | 9.6 ± 0.5 a | 5.3 ± 0.6 ab | 3.6 ± 0.0 b | 3.0 ± 0.0 b | 3.2 ± 0.0 b |
| ∑n-6 fatty acids | 99.6 ± 0.2 | 95.1 ± 0.0 | 105.3 ± 0.2 | 114.4 ± 1.3 | 117.7 ± 3.2 |
| 18:3n-3 | 15.5 ± 0.4 a | 18.6 ± 0.4 ab | 20.2 ± 0.6 b | 25.1 ± 2.1 bc | 27.9 ± 0.0 bc |
| 18:4n-3 | 12.5 ± 0.1 a | 9.1 ± 0.2 ab | 9.6 ± 0.2 ab | 7.9 ± 0.0 b | 3.9 ± 0.0 c |
| 20:3n-3 | 7.4 ± 0.3 a | 3.6 ± 0.1 b | 1.7 ± 0.0 bc | 0.6 ± 0.0 c | 0.9 ± 0.4 c |
| 20:4n-3 | 6.3 ± 0.5 a | 3.7 ± 0.02 b | 2.3 ± 0.01 b | 2.6 ± 0.00 b | 0.0 ± 0.0 c |
| 20:5n-3 | 80.5 ± 1.6 a | 59.6 ± 0.2 b | 55.7 ± 0.3 b | 47.4 ± 0.8 b | 52.5 ± 2.4 b |
| 22:5n-3 | 23.0 ± 0.6 a | 14.5.0 ± 0.2 b | 7.1 ± 0.5 c | 6.7 ± 2.0 c | 6.5 ± 0.1 c |
| 22:6n-3 | 159.0 ± 0.1 | 145 ± 0.8 | 142.2 ± 0.5 | 128.4 ± 0.7 | 120.2 ± 0.0 |
| ∑n-3 fatty acids | 304.2 ± 2.6 a | 254.9 ± 2.9 ab | 238.8 ± 0.0 ab | 218.7 ± 0.9 ab | 211.9 ± 2.1 ab |
| ∑PUFA 1 | 403.8 ± 0.9 | 350 ± 5.3 | 344.1 ± 0.0 | 333.1 ± 1.1 | 329.6 ± 0 |
| ∑-3HUFA 2 | 276.2 ± 5.9 | 226.4 ± 2.5 | 209.0 ± 2.9 | 185.7 ± 4.30 | 180.1 ± 5.2 |
| ∑n-3/n-6 ratio 3 | 3.1 | 2.7 | 2.3 | 2.0 | 1.8 |
| ∑EPA + DHA 4 | 239.5 ± 0.1 a | 204.6 ± 0.0 a | 197.9 ± 0.0 a | 175.8 ± 0.0 ab | 172.7 ± 0.3 ab |
| Fatty Acid Type | Test Diet | ||||
|---|---|---|---|---|---|
| 6F0C | 5F1C | 4F2C | 2F4C | 0F6C | |
| 14:0 | 26.8 ± 0.1 a | 17.1 ± 0.2 a | 12.4 ± 0.2 ab | 11.6 ± 0.4 abc | 8.6 ± 0.1 c |
| 16:0 | 73.0 ± 0.2 | 75.2 ± 0.1 | 82.4 ± 0.7 | 99.6 ± 0.3 | 108.2 ± 0.1 |
| 18:0 | 96.3 ± 0.0 a | 53.2 ± 0.2 ab | 45.5 ± 0.6 ab | 30.1 ± 0.6 bc | 27.9 ± 0.5 bc |
| ∑Saturated | 196.1 ± 5.6 | 145.5 ± 0.3 | 140.3 ± 0.3 | 141.3.4 ± 0.9 | 144.7 ± 0.1 |
| 16:1n-9 | 83.6 ± 0.4 a | 65.7 ± 0.2 ab | 44.0 ± 0.9 ab | 31.4 ± 0.7 ab | 29.9 ± 0.5 b |
| 18:1n-5 | 1.4 ± 0.2 | 0.2 ± 0.1 | 0.2 ± 0.1 | - ± - | - ± - |
| 18:1n-9 | 76.6 ± 2.5 a | 101.3 ± 0.5 a | 126.2 ± 0.8 ab | 145.8 ± 0.1 ab | 164.2 ± 1.0 b |
| 20:1n-9 | 21.2 ± 0.0 a | 13.5 ± 0.2 b | 14.2 ± 0.5 b | 13.6 ± 0.0 b | 11.0 ± 1.2 b |
| 22:1n-9 | 35.5 ± 0.5 | 26.0 ± 0.01 | 23.5 ± 0.0 | 20.3 ± 0.5 | 20.9 ± 0.1 |
| ∑ MUFA | 218.3 ± 1.6 | 206.7 ± 0.1 | 208.1 ± 0.2 | 211.1 ± 0.6 | 226 ± 2.1 |
| 18:2n-6 | 63.2 ± 0.1 a | 80.9 ± 0.2 ab | 118.4 ± 2.7 ab | 128.8 ± 1.5 ab | 144.0 ± 1.3 b |
| 18:3n-6 | 6.1 ± 0.2 a | 1.7 ± 0.1 b | 0.1 ± 0.2 b | 0.9 ± 0.8 b | 0.0 ± 0.0 c |
| 20:4n-6 | 8.6 ± 0.5 a | 6.2 ± 0.6 a | 4.3 ± 0.0 a | 1.0 ± 0.0 b | 0.0 ± 0.0 c |
| ∑ n-6 fatty acids | 77.9 ± 0.2 | 88.8 ± 2.1 | 122.8 ± 0.2 | 130.7 ± 1.3 | 144.0 ± 3.2 |
| 18:3n-3 | 10.6 ± 0 | 10.1 ± 0.4 | 12.4 ± 0.6 | 13.7 ± 2.1 | 14.7 ± 0.0 |
| 18:4n-3 | 4.1 ± 0.02 a | 1.3 ± 0.2 b | 1.2 ± 0.2 b | 1.2 ± 0.0 b | 1.2 ± 0.0 b |
| 20:3n-3 | 4.5 ± 0.2 a | 1.2 ± 0.1 b | 0.6 ± 0.0 c | 0.6 ± 0.0 c | 1.1 ± 0 b |
| 20:4n-3 | 5.3 ± 0.3 a | 2.1 ± 0.02 b | 2.2 ± 0.2 b | 2.2 ± 0.0 b | 0.0 ± 0.0 c |
| 20:5n-3 | 64.4 ± 1.1 a | 60.0 ± 1.6 a | 24.0 ± 3.1 b | 24.9 ± 0.8 b | 19.8 ± 0.8 b |
| 22:5n-3 | 4.0 ± 0.0 a | 0.2 ± 0.2 b | 0.1 ± 0.5 b | 0.3 ± 0.03 b | 0.1 ± 0.01 b |
| 22:6n-3 | 147.2 ± 5.3 | 133.7 ± 2.8 | 123.3 ± 1.5 | 112.4 ± 7 | 100.0 ± 0.0 |
| ∑ n-3 fatty acids | 237.8 ± 2.6 | 209.6 ± 1.7 | 161.8 ± 3.0 | 152.3 ± 1.1 | 131.9 ± 3.1 |
| ∑P UFA 1 | 315.7 ± 2.1 | 298.4 ± 5.3 | 284.6 ± 0.0 | 283.3 ± 1.1 | 275 ± 2.3 |
| ∑-3HUFA 2 | 225.4 ± 0.5 | 197 ± 2.5 | 150.2 ± 2.9 | 140.4 ± 4.30 | 121.0 ± 5.2 |
| ∑ n-3/n-6 ratio 3 | 3.0 | 2.3 | 1.3 | 1.2 | 1.0 |
| ∑ EPA + DHA 4 | 211.6 ± 1.3 a | 193.7 ± 0.0 a | 147.3 ± 0.0 a | 137.3 ± 0.0 ab | 119.8 ± 0.3 b |
| Fatty Acid Type | Test Diet | ||||
|---|---|---|---|---|---|
| 6F0C | 5F1C | 4F2C | 2F4C | 0F6C | |
| 14:0 | 91.2 ± 3 | 92.5 ± 2 | 94.1 ± 6 | 96.0 ± 1 | 96.6 ± 2 |
| 16:0 | 94.3 ± 1 | 86.4 ± 1 | 91.7 ± 07 | 92.3 ± 3 | 90.7 ± 1 |
| 18:0 | 95.3 ± 0 | 85.1 ± 2 | 95.2 ± 6 | 88.6 ± 6 | 85.5 ± 5 |
| ∑Saturated | 93.6 ± 0 | 88.0 ± 3 | 93.6 ± 3 | 92.3 ± 9 | 90.9 ± 1 |
| 16:1n-9 | 93.5 ± 1 | 93.7 ± 1 | 98.2 ± 1 | 96.9 ± 13 | 94.1 ± 0.1 |
| 18:1n-5 | 95.0 ± 2 | 97.5 ± 1 | 92.3 ± 1 | 97.7 ± 0.1 | 95.9 ± 7 |
| 18:1n-9 | 92.7 ± 0.1 | 89.6 ± 5 | 98.2 ± 8 | 96.8 ± 1 | 95.1 ± 1 |
| 20:1n-9 | 92.1 ± 2 | 92.1 ± 2 | 98.2 ± 5 | 96.1 ± 0 | 94.6 ± 21 |
| 22:1n-9 | 99.1 ± 2 | 99.4 ± 1 | 98.2 ± 2 | 95.3 ± 2 | 97.0 ± 1 |
| ∑MUFA | 94.5 ± 6 | 94.5 ± 1 | 97.0 ± 2 | 96.6 ± 6 | 95.3 ± 21 |
| 18:2n-6 | 93.6 ± 3 | 84.4 ± 3 | 98.5 ± 6 | 97.3 ± 3 | 96.1 ± 1 |
| 18:3n-6 | 94.1 ± 2 | 97.5 ± 4 | 94.6 ± 0.1 | 97.7 ± 2 | 96.3 ± 0 |
| 20:4n-6 | 93.6 ± 5 | 83.0 ± 6 | 99.0 ± 0 | 97.0 ± 0 | 96.0 ± 00 |
| 22:4n-6 | 94.2 ± 0.1 | 95.5 ± 1.0 | 98.6 ± 0.1 | 97.6 ± 0.0 | 94.7 ± 0.2 |
| ∑n-6 fatty acids | 93.9 ± 0.1 | 90.1 ± 2.1 | 97.7 ± 0.2 | 97.4 ± 2.6 | 95.8 ± 0.3 |
| 18:3n-3 | 95.1 ± 0.6 | 81.2 ± 0.9 | 99.2 ± 0.1 | 98.5 ± 4.1 | 97.9 ± 0.0 |
| 18:4n-3 | 95.5 ± 0.01 | 97.0 ± 0.2 | 99.4 ± 0.0 | 99.0 ± 0.0 | 97.1 ± 0.0 |
| 20:3n-3 | 95.3 ± 0.2 | 88.6 ± 0.8 | 99.4 ± 0.5 | 98.8 ± 2.0 | 96.7 ± 0.3 |
| 20:4n-3 | 95.1 ± 0.01 | 94.4 ± 0.1 | 98.6 ± 0.3 | 98.6 ± 0.1 | 97.4 ± 0.5 |
| 20:5n-3 | 95.7 ± 0.3 | 95.2 ± 0.5 | 98.4 ± 0.4 | 98.5 ± 0.1 | 97.8 ± 0.1 |
| 22:5n-3 | 98.7 ± 0.1 | 94.8 ± 0.7 | 99.8 ± 0.1 | 98.9 ± 0.3 | 97.2 ± 0.4 |
| 22:6n-3 | 94.6 ± 0.2 | 93.5 ± 1.5 | 99.6 ± 0.0 | 97.6 ± 0.9 | 96.6 ± 0.1 |
| ∑n-3 fatty acids | 95.7 ± 0.01 | 92.1 ± 1.7 | 99.2 ± 0.0 | 98.6 ± 0.5 | 97.2 ± 0.4 |
| ∑P UFA 1 | 94.8 ± 0.2 | 91.1 ± 3.0 | 98.4 ± 0.0 | 98.0 ± 0.2 | 96.5 ± 0.4 |
| ∑-3HUFA 2 | 95.9 ± 0.4 | 93.3 ± 0.1 | 99.2 ± 0.04 | 98.5 ± 0.2 | 97.1 ± 0.1 |
| ∑ n-3/n-6 ratio 3 | 1.02 | 1.02 | 1.01 | 1.0 | 1.04 |
| ∑ EPA + DHA 4 | 95.2 ± 0.2 | 94.3 ± 0.1 | 99.0 ± 0.2 | 98.0 ± 0.1 | 97.2 ± 0.2 |
| Parameters | Test Diets | ||||
|---|---|---|---|---|---|
| 6F0C | 5F1C | 4F2C | 2F4C | 0F6C | |
| Haematocrit (%) | 36.0 ± 1.1 | 37.3 ± 3.1 | 36.3 ± 3.1 | 36.6 ± 1.2 | 33.0 ± 4.2 |
| Glucose (mg/dL) | 72.3 ± 4.0 | 71.3 ± 14.3 | 78.3 ± 10.5 | 72.3 ± 19.0 | 88 ± 5.5 |
| Serum total protein (g/dL) | 3.1 ± 0.1 | 3.4 ± 0.2 | 3.8 ± 0.4 | 3.3 ± 0.0 | 3.4 ± 0.2 |
| Total Cholesterol (mg/dL) | 213.6 ± 8.9 | 238.0 ± 24.2 | 241.6 ± 6.9 | 196.0 ± 8.0 | 229.6 ± 21.6 |
| BUN (mg/dL) | 11.3 ± 2.0 | 8.3 ± 2.0 | 13.0 ± 3.2 | 14.6 ± 1.2 | 9.0 ± 1.1 |
| T-Bil (mg/dL) | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.4 ± 0.0 | 0.5 ± 0.0 | 0.5 ± 0.0 |
| Triglyceride (mg/dL) | 393.3 ± 62.6 | 476.6 ± 23.3 | 437.6 ± 62.3 | 415.0 ± 58.0 | 497.6 ± 2.3 |
| ALT (UI/L) | 52.3 ± 11.3 | 65.5 ± 15.4 | 64.3 ± 7.6 | 50 ± 1.3 | 53.0 ± 3 |
| AST (UI/L) | 43.0 ± 10.5 | 44.0 ± 29.1 | 38.0 ± 4.8 | 42.3 ± 9.5 | 69.3 ± 16.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mzengereza, K.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Yukun, Z.; Shadrack, R.S.; Seo, S.; Duy Khoa, T.N.; Moss, A.; Dossou, S.; et al. Effect of Substituting Fish Oil with Camelina Oil on Growth Performance, Fatty Acid Profile, Digestibility, Liver Histology, and Antioxidative Status of Red Seabream (Pagrus major). Animals 2021, 11, 1990. https://doi.org/10.3390/ani11071990
Mzengereza K, Ishikawa M, Koshio S, Yokoyama S, Yukun Z, Shadrack RS, Seo S, Duy Khoa TN, Moss A, Dossou S, et al. Effect of Substituting Fish Oil with Camelina Oil on Growth Performance, Fatty Acid Profile, Digestibility, Liver Histology, and Antioxidative Status of Red Seabream (Pagrus major). Animals. 2021; 11(7):1990. https://doi.org/10.3390/ani11071990
Chicago/Turabian StyleMzengereza, Kumbukani, Manabu Ishikawa, Shunsuke Koshio, Saichiro Yokoyama, Zhang Yukun, Ronick S. Shadrack, Seok Seo, Tran Nguyen Duy Khoa, Amina Moss, Serge Dossou, and et al. 2021. "Effect of Substituting Fish Oil with Camelina Oil on Growth Performance, Fatty Acid Profile, Digestibility, Liver Histology, and Antioxidative Status of Red Seabream (Pagrus major)" Animals 11, no. 7: 1990. https://doi.org/10.3390/ani11071990
APA StyleMzengereza, K., Ishikawa, M., Koshio, S., Yokoyama, S., Yukun, Z., Shadrack, R. S., Seo, S., Duy Khoa, T. N., Moss, A., Dossou, S., Basuini, M. F. E., & Dawood, M. A. O. (2021). Effect of Substituting Fish Oil with Camelina Oil on Growth Performance, Fatty Acid Profile, Digestibility, Liver Histology, and Antioxidative Status of Red Seabream (Pagrus major). Animals, 11(7), 1990. https://doi.org/10.3390/ani11071990