
Effects of Dietary Valine Levels on Production Performance, Egg Quality, Antioxidant Capacity, Immunity, and Intestinal Amino Acid Absorption of Laying Hens during the Peak Lay Period

 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diets, Birds, and Management
2.2. Productive Performance and Determination of Egg Quality
2.3. Sample Collection and Processing
2.4. Serum Index and Digestive Enzyme Assays
2.5. Determination of Serum Free Amino Acids
2.6. Total RNA Extraction and Real-Time PCR
2.7. Statistical Analysis
3. Results
3.1. Laying Hens Performance
3.2. Egg Quality
3.3. Serum Antioxidant Capacity
3.4. Serum Immunoglobulins Levels
3.5. Digestive Enzymes Activities
3.6. Serum Free Amino Acids
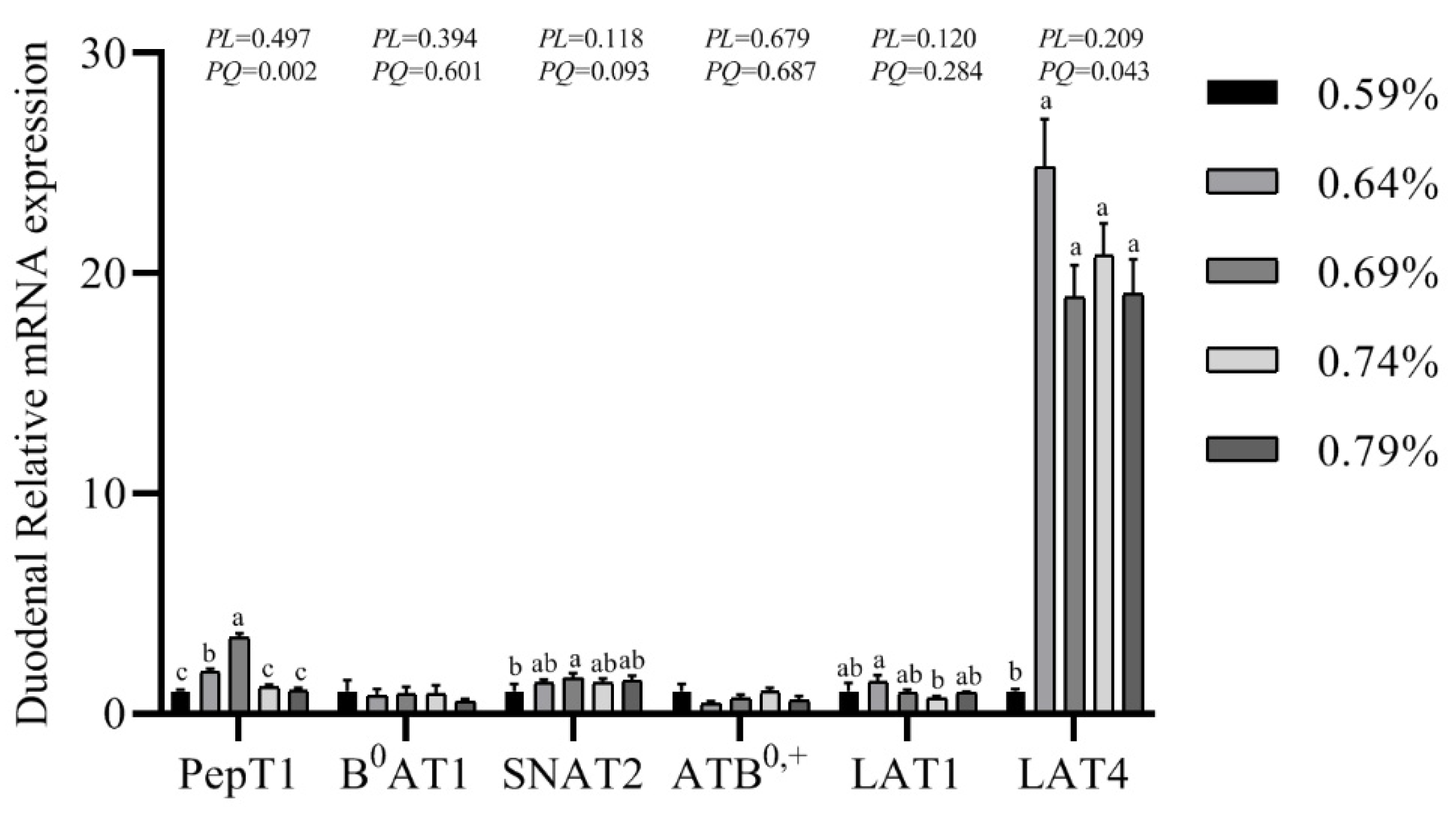
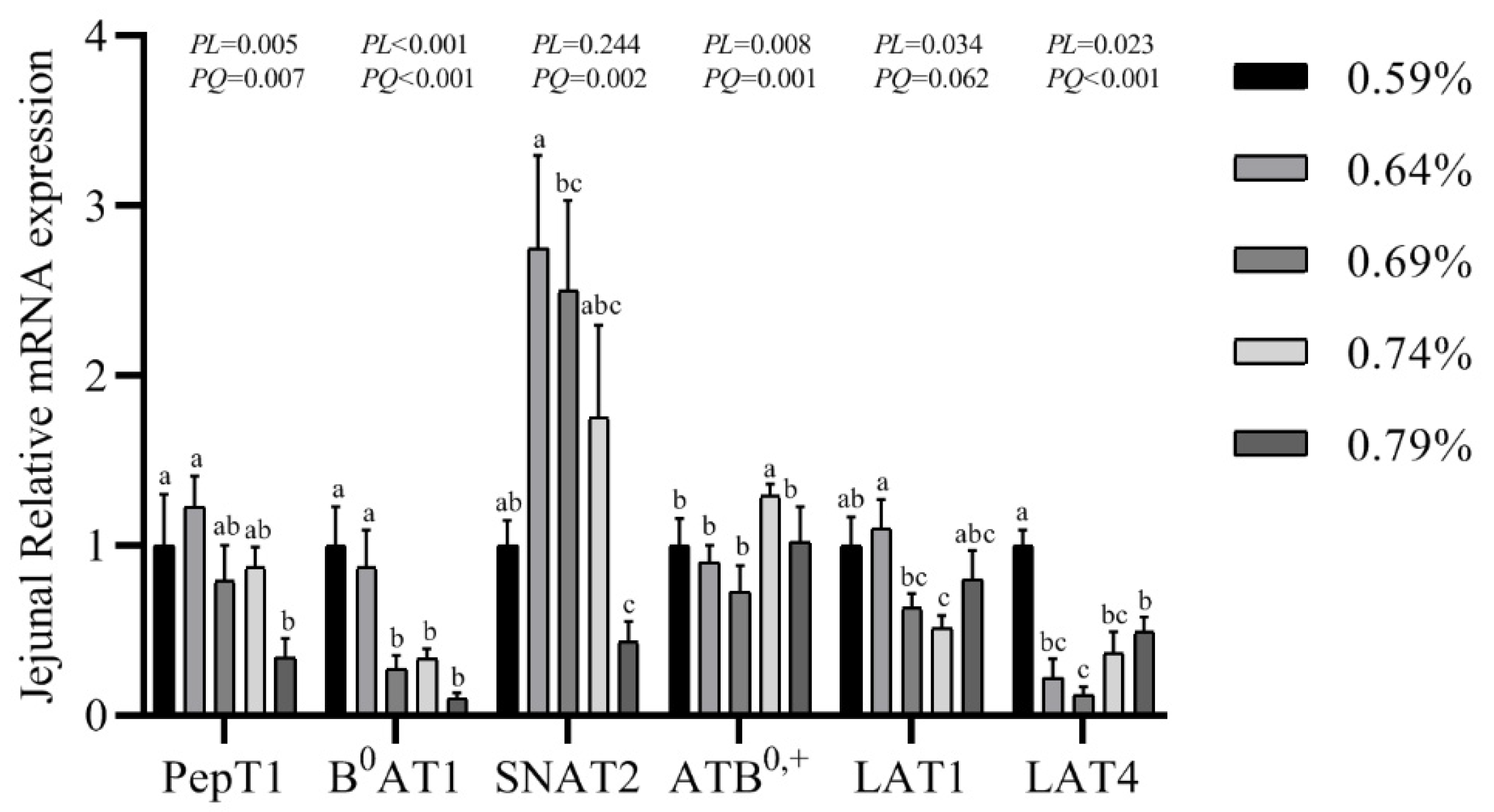
3.7. Gene mRNA Expression Levels of PepT1, B0AT1, SNAT2, ATB0,+, LAT1 and LAT4
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbate, J.M.; Macrì, F.; Capparucci, F.; Iaria, C.; Briguglio, G.; Cicero, L.; Salvo, A.; Arfuso, F.; Ieni, A.; Piccione, G.; et al. Administration of Protein Hydrolysates from Anchovy (Engraulis encrasicolus) Waste for Twelve Weeks Decreases Metabolic Dysfunction-Associated Fatty Liver Disease Severity in ApoE−/− Mice. Animals 2020, 10, 2303. [Google Scholar] [CrossRef]
- Avondo, M.; Pagano, R.I.; Guastella, A.M.; Criscione, A.; Di Gloria, M.; Valenti, B.; Piccione, G.; Pennisi, P. Diet Selection and Milk Production and Composition in Girgentana Goats with Different Alpha s1-casein Genotype. J. Dairy Res. 2009, 76, 202–209. [Google Scholar] [CrossRef]
- Monteverde, V.; Congiu, F.; Vazzana, I.; Dara, S.; Di Pietro, S.; Piccione, G. Serum Lipid Profile Modification Related to Polyunsaturated Fatty Acid Supplementation in Thoroughbred Horses. J. Appl. Anim. Res. 2017, 45, 615–618. [Google Scholar] [CrossRef]
- Armato, L.; Gianesella, M.; Morgante, M.; Fiore, E.; Rizzo, M.; Giudice, E.; Piccione, G. Rumen Volatile Fatty Acids × Dietary Supplementation with Live Yeast and Yeast Cell Wall in Feedlot Beef Cattle. Acta Agric. Scand. Sect. A Anim. Sci. 2016, 66, 119–124. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Brosnan, M.E. Branched-Chain Amino Acids: Enzyme and Substrate Regulation. J. Nutr. 2006, 136, 207S–211S. [Google Scholar] [CrossRef]
- Ferrando, A.A.; Williams, B.D.; Stuart, C.A.; Lane, H.W.; Wolfe, R.R. Oral Branched-Chain Amino Acids Decrease Whole-Body Proteolysis. Jpn. J. Parenter. Enter. Nutr. 1995, 19, 47–54. [Google Scholar] [CrossRef]
- Wu, G. Amino Acids: Metabolism, Functions, and Nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [PubMed]
- Lelis, G.R.; Albino, F.T.; Tavernari, F.C.; Calderano, A.A.; Rostagno, H.S.; Barros, V.R.S.M.; Maia, R.C. Digestible Valine-to-Digestible Lysine Ratios in Brown Commercial Layer Diets. J. App. Poult. Res. 2014, 23, 683–690. [Google Scholar] [CrossRef]
- Harper, A.E.; Miller, R.H.; Block, K.P. Branched-Chain Amino Acid Metabolism. Annu. Rev. Nutr. 1984, 4, 409–454. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Poultry, 9th ed.; National Academic Press: Washington, DC, USA, 1994. [Google Scholar]
- The Agricultural Industry Standard of the People’s Republic of China-Chicken Feeding Standard (NY/T33-2004); Ministry of Agriculture of the PRC: Beijing, China, 2006; Volume 4, pp. 19–27. (In Chinese)
- Harms, R.H.; Russell, G.B. Evaluation of Valine Requirement of the Commercial Layer Using a Corn-Soybean Meal Basal Diet. Poult. Sci. 2001, 80, 215–218. [Google Scholar] [CrossRef]
- Bregendahl, K.; Roberts, S.A.; Kerr, B.; Hoehler, D. Ideal Ratios of Isoleucine, Methionine, Methionine Plus Cystine, Threonine, Tryptophan, and Valine Relative to Lysine for White Leghorn-type Laying Hens of Twenty-Eight to Thirty-Four Weeks of Age. Poult. Sci. 2008, 87, 744–758. [Google Scholar] [CrossRef]
- Eder, K.; Peganova, S. Studies on the Requirement and Excess of Valine in Laying Hens. Archiv. Gefl. 2002, 66, 250–251. [Google Scholar]
- Wen, J.; Helmbrecht, A.; Elliot, M.A.; Thomson, J.; Persia, M.E. Evaluation of the Valine Requirement of Small-Framed First Cycle Laying Hens. Poult. Sci. 2019, 98, 1272–1279. [Google Scholar] [CrossRef]
- Fisher, H.; Johnson, D., Jr. The Amino Acid Requirement of the Laying Hen. II. Classification of the Essential Amino Acids Required for Egg Production. J. Nutr. 1956, 60, 275–282. [Google Scholar] [PubMed]
- Zhang, S.; Saremi, B.; Gilbert, E.R.; Wong, E.A. Physiological and Biochemical Aspects of Methionine Isomers and a Methionine Analogue in Broilers. Poult. Sci. 2017, 96, 425–439. [Google Scholar] [CrossRef]
- Daniel, H. Molecular and Integrative Physiology of Intestinal Peptide Transport. Annu. Rev. Physiol. 2004, 66, 361–384. [Google Scholar] [CrossRef] [PubMed]
- Bröer, A.; Klingel, K.; Kowalczuk, S.; Rasko, J.E.; Cavanaugh, J.; Bröer, S. Molecular Cloning of Mouse Amino Acid Transport System B0, a Neutral Amino Acid Transporter Related to Hartnup Disorder. J. Biol. Chem. 2004, 279, 24467–24476. [Google Scholar] [CrossRef]
- Hundal, H.S.; Taylor, P.M. Amino Acid Transceptors: Gate Keepers of Nutrient Exchange and Regulators of Nutrient Signaling. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E603–E613. [Google Scholar] [CrossRef]
- Van Winkle, L.J.; Christensen, H.N.; Campione, A.L. Na+-Dependent Transport of Basic, Zwitterionic, and Bicyclic Amino Acids by a Broad-Scope System in Mouse Blastocysts. J. Biol. Chem. 1985, 260, 12118–12123. [Google Scholar] [CrossRef]
- Wang, Q.; Holst, J. L-type Amino Acid Transport and Cancer: Targeting the mTORC1 Pathway to Inhibit Neoplasia. Am. J. Cancer Res. 2015, 5, 1281–1294. [Google Scholar]
- Bodoy, S.; Martín, L.; Zorzano, A.; Palacín, M.; Estévez, R.; Bertran, J. Identification of LAT4, a Novel Amino Acid Transporter with System L Activity. J. Biol. Chem. 2005, 280, 12002–12011. [Google Scholar] [CrossRef]
- Gilbert, E.R.; Li, H.; Emmerson, D.A.; Webb, K.E.; Wong, E.A. Developmental Regulation of Nutrient Transporter and Enzyme mRNA Abundance in the Small Intestine of Broilers. Poult. Sci. 2007, 86, 1739–1753. [Google Scholar] [CrossRef]
- Baird, F.E.; Bett, K.J.; MacLean, C.; Tee, A.R.; Hundal, H.S.; Taylor, P.M. Tertiary Active Transport of Amino Acids Reconstituted by Coexpression of System A and L Transporters in Xenopus oocytes. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E822–E829. [Google Scholar] [CrossRef]
- Mehta, A.; Singh, S.; Dhawan, V.; Ganguly, N.K. Intestinal Mucosal Lipid Peroxidation and Absorptive Function in Salmonella Typhimurium Mediated Intestinal Infection. Mol. Cell Biochem. 1998, 178, 345–352. [Google Scholar] [CrossRef]
- Alteheld, B.; Evans, M.E.; Gu, L.H.; Ganapathy, V.; Leibach, F.H.; Jones, D.P.; Ziegler, T.R. Alanylglutamine Dipeptide and Growth Hormone Maintain PepT1-mediated Transport in Oxidatively Stressed Caco-2 Cells. J. Nutr. 2005, 135, 19–26. [Google Scholar] [CrossRef][Green Version]
- Xu, Q.Q.; Azzam, M.M.; Zou, X.T.; Dong, X.Y. Effects of Chitooligosaccharide Supplementation on Laying Performance, Egg Quality, Blood Biochemistry, Antioxidant Capacity and Immunity of Laying Hens During the Late Laying Period. Ital. J. Anim. Sci. 2020, 19, 1181–1188. [Google Scholar] [CrossRef]
- Fang, Q. Determination of Production Performance and Studies of the Profile of Growth and Laying Rate in Fengda-1 Layer. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2017. [Google Scholar]
- Liu, X.; Zhao, H.L.; Thiessen, S.; House, J.D.; Jones, P.J. Relationship of Rate of Egg Production as Affected by Feed to Haugh Units of Eggs. Poult. Sci. 1960, 39, 75–76. [Google Scholar]
- Scott, M.L.; Hull, S.J.; Mullenhoff, P.A. The Calcium Requirements of Laying Hens and Effects of Dietary Oyster Shell Upon Egg Shell Quality. Poult. Sci. 1971, 50, 1055–1063. [Google Scholar] [CrossRef]
- Götz, M.E.; Künig, G.; Riederer, P.; Youdim, M.B. Oxidative Stress: Free Radical Production in Neural Degeneration. Pharmacol. Ther. 1994, 63, 37–122. [Google Scholar] [CrossRef]
- Dong, X.Y.; Azzam, M.M.; Rao, W.; Yu, D.Y.; Zou, X.T. Evaluating the Impact of Excess Dietary Tryptophan on Laying Performance and Immune Function of Laying Hens Reared Under Hot and Humid Summer Conditions. Brit. Poult. Sci. 2012, 53, 491–496. [Google Scholar] [CrossRef]
- Azzam, M.M.; Dong, X.Y.; Xie, P.; Zou, X.T. Influence of L-threonine Supplementation on Goblet Cell Numbers, Histological Structure and Antioxidant Enzyme Activities of Laying Hens Reared in a Hot and Humid Climate. Brit. Poult. Sci. 2012, 53, 640–645. [Google Scholar] [CrossRef]
- Azzam, M.M.; Dong, X.Y.; Dai, L.; Zou, X.T. Effect of Excess Dietary L-valine on Laying Hen Performance, Egg quality, Serum Free Amino Acids, Immune Function and Antioxidant Enzyme Activity. Brit. Poult. Sci. 2015, 56, 72–78. [Google Scholar] [CrossRef]
- Dong, X.Y.; Azzam, M.M.; Zou, X.T. Effects of Dietary L-isoleucine on Laying Performance and Immunomodulation of Laying Hens. Poult. Sci. 2016, 95, 2297–2305. [Google Scholar] [CrossRef]
- Hale, L.L.; Pharr, G.T.; Burgess, S.C.; Corzo, A.; Kidd, M.T. Isoleucine Needs of Thirty- to Forty-day-old Female Chickens: Immunity. Poult. Sci. 2004, 83, 1979–1985. [Google Scholar] [CrossRef]
- Rideau, N.; Nitzan, Z.; Mongin, P. Activities of Amylase, Trypsin and Lipase in the Pancreas and Small Intestine of the Laying Hen During Egg Formation. Brit. Poult. Sci. 1983, 24, 1–9. [Google Scholar] [CrossRef]
- Kadam, M.M.; Bhanja, S.K.; Mandal, A.B.; Thakur, R.; Vasan, P.; Bhattacharyya, A.; Tyagi, J.S. Effect of in ovo Threonine Supplementation on Early Growth, Immunological Responses and Digestive Enzyme Activities in Broiler Chickens. Brit. Poult. Sci. 2008, 49, 736–741. [Google Scholar] [CrossRef]
- Zhang, J.C.; Chen, P.; Zhang, C.; Khalil, M.M.; Zhang, N.Y.; Qi, D.S.; Wang, Y.W.; Sun, L.H. Yeast Culture Promotes the Production of Aged Laying Hens by Improving Intestinal Digestive Enzyme activities and the Intestinal Health Status. Poult. Sci. 2020, 99, 2026–2032. [Google Scholar] [CrossRef]
- Ishibashi, T.; Ogawa, Y.; Itoh, T.; Fujimura, S.; Koide, K.; Watanabe, R. Threonine Requirements of Laying Hens. Poult. Sci. 1998, 77, 998–1002. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, S.; Bao, J.; He, X.; Tong, D.; Chen, C.; Ying, Q.; Zhang, Q.; Zhang, C.; Li, J. Melatonin Improves Laying Performance by Enhancing Intestinal Amino Acids Transport in Hens. Front. Endocrinol. 2018, 9, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Azzam, M.M.; Yuan, C.; Liu, G.H.; Zou, X.T. Effect of Excess Dietary Threonine on Laying Performance, Egg quality, Serum free Amino Acids, and Digestive Enzymes Activities of Laying Hens During the Postpeak Period. J. App. Poult. Res. 2014, 23, 605–613. [Google Scholar] [CrossRef]
- Silk, D.B.; Grimble, G.K.; Rees, R.G. Protein Digestion and Amino Acid and Peptide Absorption. Proc. Nutr. Soc. 1985, 44, 63–72. [Google Scholar] [CrossRef]
- Chen, H.; Pan, Y.; Wong, E.A.; Bloomquist, J.R.; Webb, K.E. Molecular Cloning and Functional Expression of a Chicken Intestinal Peptide Pransporter (cPepT1) in Xenopus Oocytes and Chinese Hamster Ovary Cells. J. Nutr. 2002, 132, 387–393. [Google Scholar] [CrossRef]
- Chen, H.; Wong, E.A.; Webb, K.E. Tissue Distribution of a Peptide Transporter mRNA in Sheep, Dairy cows, Pigs, and Chickens. J. Anim. Sci. 1999, 77, 1277–1283. [Google Scholar] [CrossRef]
- Jiang, S.; El-Senousey, H.K.; Fan, Q.; Lin, X.; Gou, Z.; Li, L.; Wang, Y.; Fouad, A.M.; Jiang, Z. Effects of Dietary Threonine Supplementation on Productivity and Expression of Genes Related to Protein Deposition and Amino Acid Transportation in Breeder Hens of Yellow-Feathered Chicken and Their Offspring. Poult. Sci. 2019, 98, 6826–6836. [Google Scholar] [CrossRef]
- Moreira Filho, A.; Ferket, P.R.; Malheiros, R.D.; Oliveira, C.; Aristimunha, P.C.; Wilsmann, D.E.; Givisiez, P. Enrichment of the Amnion with Threonine in Chicken Embryos Affects the Small Intestine Development, Ileal Gene Expression and Performance of Broilers Between 1 and 21 Days of Age. Poult. Sci. 2019, 98, 1363–1370. [Google Scholar] [CrossRef]
- Terada, T.; Shimada, Y.; Pan, X.; Kishimoto, K.; Sakurai, T.; Doi, R.; Onodera, H.; Katsura, T.; Imamura, M.; Inui, K. Expression Profiles of Various Transporters for Oligopeptides, Amino Acids and Organic Ions Along the Human Digestive Tract. Biochem. Pharm. 2005, 70, 1756–1763. [Google Scholar] [CrossRef]
- Romeo, E.; Dave, M.H.; Bacic, D.; Ristic, Z.; Camargo, S.M.; Loffing, J.; Wagner, C.A.; Verrey, F. Luminal Kidney and Intestine SLC6 Amino Acid Transporters of B0AT-cluster and Their Tissue Distribution in Mus Musculus. Am. J. Phys. Ren. Phys. 2006, 290, F376–F383. [Google Scholar] [CrossRef]
- Jiang, S.Q.; Gou, Z.Y.; Lin, X.J.; Li, L. Effects of Dietary Tryptophan Levels on Performance and Biochemical Variables of Plasma and Intestinal Mucosa in Yellow-Feathered Broiler Breeders. J. Anim. Phys. Anim. Nutr. 2018, 102, e387–e394. [Google Scholar] [CrossRef] [PubMed]
- Franchi-Gazzola, R.; Dall’Asta, V.; Sala, R.; Visigalli, R.; Bevilacqua, E.; Gaccioli, F.; Gazzola, G.C.; Bussolati, O. The Role of the Neutral Amino Acid Transporter SNAT2 in Cell Volume Regulation. Acta Phys. 2006, 187, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, T.; Huang, W.; Nakanishi, T.; Bridges, C.C.; Smith, S.B.; Prasad, P.D.; Ganapathy, M.E.; Ganapathy, V. Transport of D-Serine via the Amino Acid Transporter ATB(0,+) Expressed in the Colon. Biochem. Biophys. Res. Commun. 2002, 291, 291–295. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, J.K.; Kim, I.J.; Choi, B.K.; Jung, K.Y.; Lee, S.; Park, K.J.; Chairoungdua, A.; Kanai, Y.; Endou, H.; et al. Reabsorption of Neutral Amino Acids Mediated by Amino Acid Transporter LAT2 and TAT1 in the Basolateral Membrane of Proximal Tubule. Arch. Pharm. Res. 2005, 28, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Sumners, L.H.; Dalloul, R.A.; Miska, K.B.; Fetterer, R.H.; Jenkins, M.C.; Zhu, Q.; Wong, E.A. Changes in Expression of an Antimicrobial Peptide, Digestive Enzymes, and Nutrient Transporters in the Intestine of E. praecox-Infected Chickens. Poult. Sci. 2015, 94, 1521–1526. [Google Scholar] [CrossRef] [PubMed]
- Bröer, S.; Fairweather, S.J. Amino Acid Transport Across the Mammalian Intestine. Compr. Physiol. 2018, 13, 343–373. [Google Scholar]
- Guetg, A.; Mariotta, L.; Bock, L.; Herzog, B.; Fingerhut, R.; Camargo, S.M.; Verrey, F. Essential Amino Acid Transporter Lat4 (Slc43a2) is Required for Mouse Development. J. Phys. 2015, 593, 1273–1289. [Google Scholar] [CrossRef] [PubMed]
- Geier, E.G.; Schlessinger, A.; Fan, H.; Gable, J.E.; Irwin, J.J.; Sali, A.; Giacomini, K.M. Structure-Based Ligand Discovery for the Large-neutral Amino Acid Transporter 1, LAT-1. Proc. Natl. Acad. Sci. USA 2013, 110, 5480–5485. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Dietary Valine Levels (%) a | ||||
|---|---|---|---|---|---|
| 0.59 | 0.64 | 0.69 | 0.74 | 0.79 | |
| Corn | 66.6 | 66.6 | 66.6 | 66.6 | 66.6 |
| Soybean meal | 10.5 | 10.65 | 11.2 | 11.55 | 11.8 |
| Wheat bran | 2.9 | 2.9 | 2.91 | 2.92 | 2.92 |
| Peanut meal | 8.7 | 8.5 | 7.9 | 7.5 | 7.2 |
| Limestone | 9.3 | 9.3 | 9.3 | 9.3 | 9.3 |
| Soybean oil | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| DL-Methionine (98%) | 0.16 | 0.16 | 0.15 | 0.15 | 0.15 |
| Lysine (78%) | 0.11 | 0.11 | 0.11 | 0.1 | 0.1 |
| Valine (98%) | 0 | 0.0508 | 0.1016 | 0.1523 | 0.2031 |
| CaHPO4 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 |
| Salt | 0.36 | 0.36 | 0.36 | 0.36 | 0.36 |
| Choline chloride, 60% | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Mineral and vitamin premix b | 0.27 | 0.27 | 0.27 | 0.27 | 0.27 |
| Calculated nutritional level, % | |||||
| Crude protein (CP) | 14.7 | 14.7 | 14.7 | 14.7 | 14.7 |
| ME, MJ/Kg | 2.68 | 2.68 | 2.68 | 2.68 | 2.68 |
| Analyzed nutrients | |||||
| Crude protein (CP) | 14.65 | 14.72 | 14.75 | 14.74 | 14.78 |
| Calcium | 3.58 | 3.58 | 3.59 | 3.59 | 3.59 |
| Total phosphorus | 0.46 | 0.46 | 0.46 | 0.46 | 0.46 |
| Methionine | 0.36 | 0.36 | 0.36 | 0.37 | 0.37 |
| Lysine | 0.66 | 0.66 | 0.66 | 0.67 | 0.67 |
| Threonine | 0.48 | 0.48 | 0.48 | 0.49 | 0.49 |
| Tryptophan | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 |
| Arginine | 1.05 | 1.04 | 1.03 | 1.03 | 1.02 |
| Valine | 0.59 | 0.64 | 0.69 | 0.74 | 0.79 |
| Gene Name | Primer | Primer Sequence (5′-3′) | Accession No. |
|---|---|---|---|
| β-Actin | Forward | TCCCTGGAGAAGAGCTATGAA | NM_205518.1 |
| Reverse | CAGGACTCCATACCCAAGAAAG | ||
| PepT1 | Forward | CTTGGCAGATCCCTCAGTATTT | XM_034074354.1 |
| Reverse | GTTGGGCTTCAACCTCATTTG | ||
| B0AT1 | Forward | CATGATCGGACACAAGCCCA | XM_419056.6 |
| Reverse | AGCATAGACCCAGCCAGGATA | ||
| ATB0,+ | Forward | TAAACCAGTGCAATTTCCCA | XM_001199603.1 |
| Reverse | CGATGTTGCCAGTCTCATC | ||
| SNAT2 | Forward | GAAGGAGTTCAGTTGGTGGCG | NM_001305439.1 |
| Reverse | CGGATAGTAGGGACAAAGATAACGAG | ||
| LAT1 | Forward | TGGCCTTGTAC AGTGGTCTT | NM_001030579.2 |
| Reverse | GCTTCGGACTTC AGC ATCTG | ||
| LAT4 | Forward | ACAACTGTGGGACGCCGACTGA | XM_415803.6 |
| Reverse | GGCATTGGTGGCATTGGTGATTT |
| Item | Dietary Valine Levels (%) | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0.59 | 0.64 | 0.69 | 0.74 | 0.79 | L | Q | ||
| Egg mass (a/repeat/d) | 25.99 ab | 25.87 b | 26.23 ab | 26.31 ab | 26.43 a | 0.14 | 0.003 | 0.012 |
| Laying rate (%) 1 | 81.63 | 81.28 | 82.46 | 82.52 | 82.86 | 0.43 | 0.007 | 0.026 |
| Egg weight (g) | 48.50 a | 48.50 a | 48.28 b | 48.31 ab | 48.27 b | 0.08 | 0.011 | 0.034 |
| Broken egg rate (%) | 0.39 a | 0.24 ab | 0.24 ab | 0.24 ab | 0.13 b | 0.05 | 0.003 | 0.011 |
| Feed intake (g/hen/d) | 100.90 | 101.20 | 100.45 | 100.83 | 100.34 | 2.30 | 0.835 | 0.977 |
| FCR (kg feed/kg egg) | 2.55 a | 2.56 a | 2.52 bc | 2.53 b | 2.51 c | 0.02 | 0.001 | 0.003 |
| Item | Dietary Valine Levels (%) | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0.59 | 0.64 | 0.69 | 0.74 | 0.79 | L | Q | |||
| Egg weight, g | 50.32 | 51.03 | 50.34 | 49.55 | 49.28 | 0.67 | 0.095 | 0.180 | |
| Eggshell strength (N) | 4.88 | 4.58 | 4.65 | 4.75 | 4.80 | 0.15 | 0.944 | 0.417 | |
| Eggshell thickness (mm−2) | Sharp end | 0.42 ab | 0.42 ab | 0.41 ab | 0.43 a | 0.40 b | 0.005 | 0.024 | 0.048 |
| Equator | 0.40 | 0.40 | 0.39 | 0.41 | 0.38 | 0.005 | 0.370 | 0.143 | |
| Blunt end | 0.39 | 0.38 | 0.38 | 0.39 | 0.38 | 0.006 | 0.473 | 0.774 | |
| Albumen height (mm) | 5.19 ab | 5.78 a | 5.18 ab | 5.09 ab | 4.67 b | 0.20 | 0.010 | 0.006 | |
| Haugh unit | 74.45 ab | 77.23 a | 74.73 ab | 71.45 ab | 69.40 b | 1.85 | 0.011 | 0.014 | |
| Egg yolk color (points) | 10.34 a | 10.43 a | 10.53 a | 10.43 a | 9.07 b | 0.13 | <0.001 | <0.001 | |
| Serum | Dietary Valine Levels (%) | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0.59 | 0.64 | 0.69 | 0.74 | 0.79 | L | Q | ||
| CAT, U/mL | 8.19 ab | 6.59 ab | 11.55 a | 7.24 ab | 5.74 b | 1.71 | 0.229 | 0.041 |
| T-SOD, U/mL | 175.79 | 175.33 | 187.55 | 175.85 | 166.15 | 7.46 | 0.544 | 0.255 |
| T-AOC, U/mL | 0.90 ab | 0.92 ab | 0.76 b | 0.85 ab | 1.02 a | 0.05 | 0.312 | 0.025 |
| GSH-Px, U/mL | 274.29 | 180.57 | 278.79 | 278.06 | 259.71 | 15.19 | 0.229 | 0.488 |
| MDA, nmol/mL | 2.97 ab | 2.90 ab | 2.49 b | 3.14 ab | 4.80 a | 0.40 | 0.029 | 0.002 |
| Amino Acids, mg/L | Dietary Valine Levels (%) | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0.59 | 0.64 | 0.69 | 0.74 | 0.79 | L | Q | ||
| Essential AA | ||||||||
| Arginine | 102.01 b | 99.83 c | 107.65 a | 103.65 b | 96.40 d | 0.41 | 0.244 | 0.001 |
| Histidine | 2.09 | 2.09 | 1.91 | 1.85 | 1.87 | 0.05 | 0.002 | 0.008 |
| Isoleucine | 10.90 d | 11.73 c | 12.67 b | 13.33 ab | 13.86 a | 0.18 | <0.001 | <0.001 |
| Leucine | 19.29 ab | 18.90 b | 19.75 a | 17.76 c | 18.71 b | 0.14 | 0.042 | 0.134 |
| Lysine | 32.81 b | 32.98 b | 33.69 b | 35.15 a | 35.75 a | 0.28 | <0.001 | <0.001 |
| Methionine | 8.89 a | 8.30 b | 7.98 c | 7.45 d | 7.17 e | 0.05 | <0.001 | <0.001 |
| Phenylalanine | 17.53 b | 17.17 b | 18.52 a | 17.46 b | 18.77 a | 0.18 | 0.011 | 0.032 |
| Threonine | 50.65 a | 48.80 c | 49.93 a | 46.96 b | 50.06 a | 0.15 | 0.169 | 0.019 |
| Valine | 13.02 e | 13.90 d | 15.06 c | 16.68 b | 18.14 a | 0.17 | <0.001 | <0.001 |
| Nonessential AA | ||||||||
| Alanine | 81.67 a | 78.50 b | 76.12 c | 75.08 c | 78.40 b | 0.32 | 0.005 | <0.001 |
| Aspartic acid | 2.58 a | 2.41 d | 2.45 cd | 2.49 bc | 2.52 ab | 0.01 | 0.169 | 0.019 |
| Cystine | 24.56 b | 22.31 c | 26.36 a | 21.03 d | 23.60 b | 0.22 | 0.304 | 0.598 |
| Glutamic acid | 31.00 b | 33.59 a | 32.41 ab | 31.47 b | 30.57 b | 0.29 | 0.165 | 0.004 |
| Glycine | 56.67 a | 51.47 c | 53.74 b | 49.39 d | 52.04 c | 0.26 | 0.002 | <0.001 |
| Proline | 16.61 | 16.30 | 17.01 | 16.40 | 16.58 | 0.30 | 0.966 | 0.962 |
| Serine | 54.02 a | 52.98 b | 51.31 c | 50.70 c | 50.32 c | 0.26 | 0.165 | 0.004 |
| Tyrosine | 34.17 a | 30.20 c | 32.30 b | 33.26 ab | 33.49 a | 0.25 | 0.480 | 0.015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jian, H.; Miao, S.; Liu, Y.; Li, H.; Zhou, W.; Wang, X.; Dong, X.; Zou, X. Effects of Dietary Valine Levels on Production Performance, Egg Quality, Antioxidant Capacity, Immunity, and Intestinal Amino Acid Absorption of Laying Hens during the Peak Lay Period. Animals 2021, 11, 1972. https://doi.org/10.3390/ani11071972
Jian H, Miao S, Liu Y, Li H, Zhou W, Wang X, Dong X, Zou X. Effects of Dietary Valine Levels on Production Performance, Egg Quality, Antioxidant Capacity, Immunity, and Intestinal Amino Acid Absorption of Laying Hens during the Peak Lay Period. Animals. 2021; 11(7):1972. https://doi.org/10.3390/ani11071972
Chicago/Turabian StyleJian, Huafeng, Sasa Miao, Yating Liu, Huaiyu Li, Wenting Zhou, Xiaoming Wang, Xinyang Dong, and Xiaoting Zou. 2021. "Effects of Dietary Valine Levels on Production Performance, Egg Quality, Antioxidant Capacity, Immunity, and Intestinal Amino Acid Absorption of Laying Hens during the Peak Lay Period" Animals 11, no. 7: 1972. https://doi.org/10.3390/ani11071972
APA StyleJian, H., Miao, S., Liu, Y., Li, H., Zhou, W., Wang, X., Dong, X., & Zou, X. (2021). Effects of Dietary Valine Levels on Production Performance, Egg Quality, Antioxidant Capacity, Immunity, and Intestinal Amino Acid Absorption of Laying Hens during the Peak Lay Period. Animals, 11(7), 1972. https://doi.org/10.3390/ani11071972