
Overview of the Genus Boleodorus and First Reports of Boleodorus thylactus and B. volutus from Southern Alberta, Canada



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Studies
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogenetic Analyses
3. Results
3.1. Systematics
3.1.1. Morphological and Morphometrical Observations on Boleodorus Species

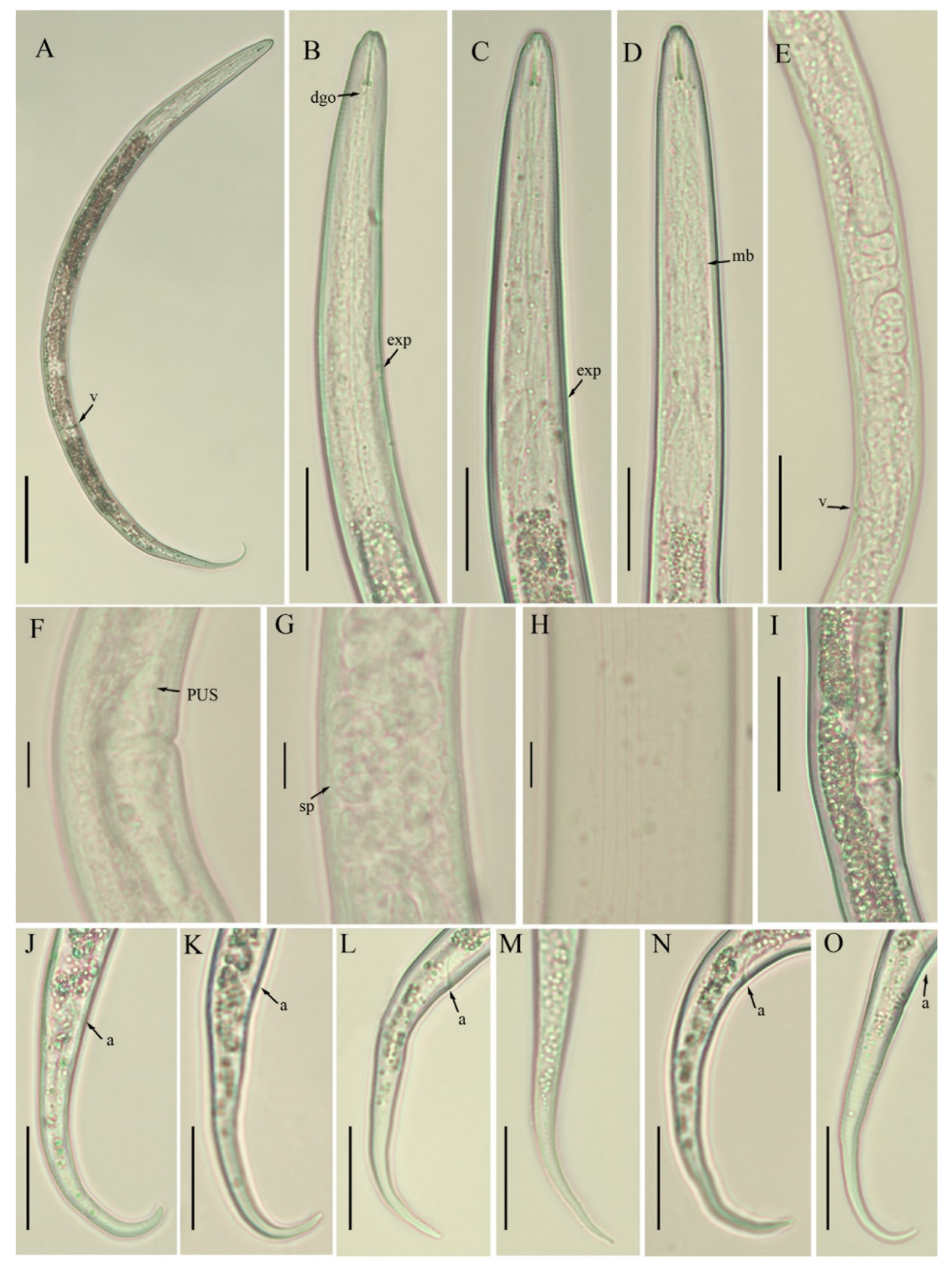{kind=link}
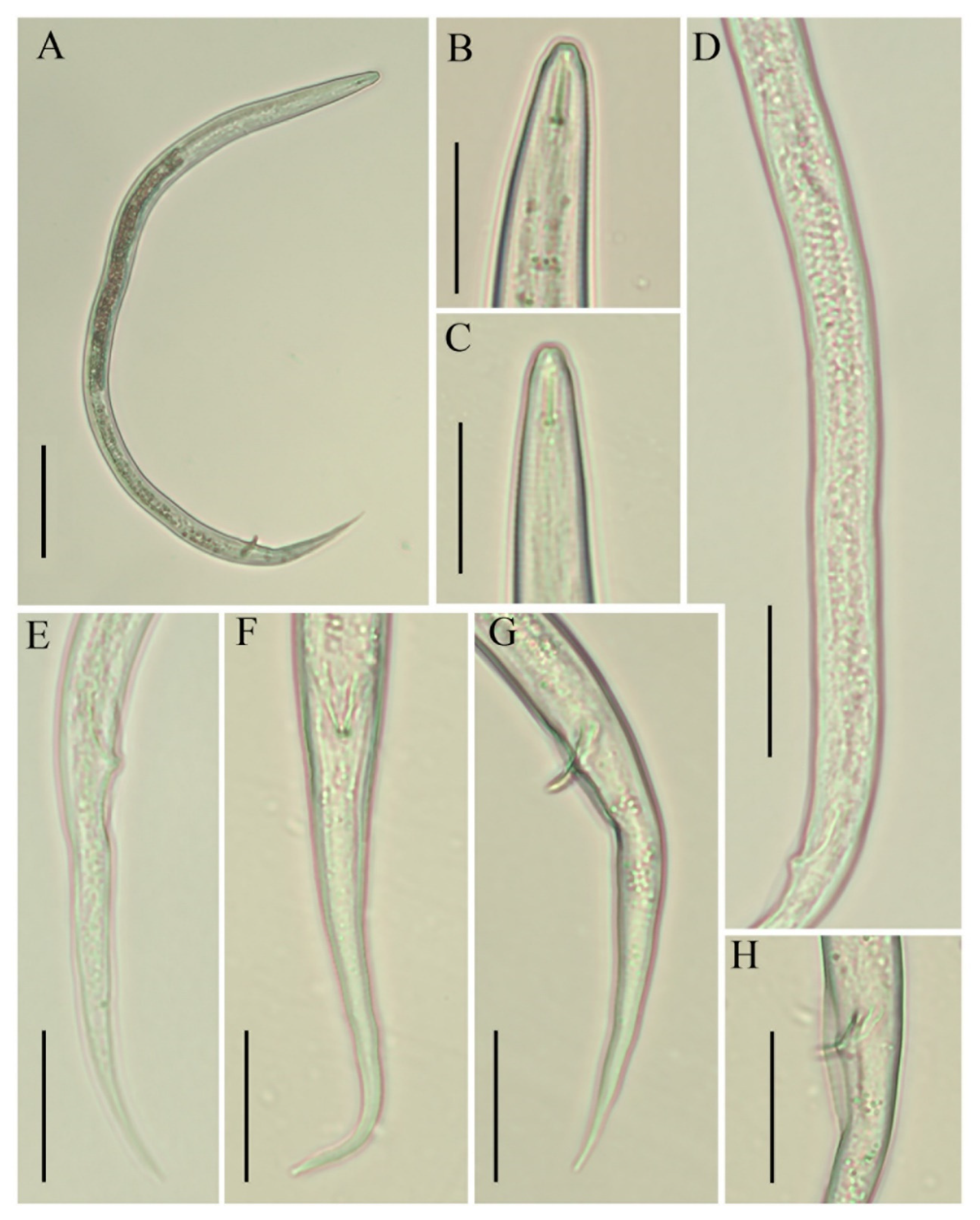{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Body Habitus | Lip Shape | Tail Shape | Tail Length | L | a | b | c | c′ | V | Stylet Length | DGO | 2 SE Pore Dist. | 3 LL | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 B. acurvus | Almost straight | Continuous, narrow conical | Filiform, terminus clavate | 75–90 | 470–540 | 27–37 | 4.2–4.7 | 6–9 | 7–9 | 60–63 | 10–11 | 4–6 | 80–90 | 4 | Jairajpuri [44] |
| Almost straight | Continuous, conical, depression at OA 1 | Filiform, terminus rounded | 77–93 | 475–605 | 27–39 | 4.1–5.1 | 5.7–6.9 | 7.2–8.5 | 62–64 | 10.5–11.5 | – | 79–95 | 4 | Zeidan and Geraert [50] | |
| 2 B. acutus | Arcuate to open C-shape | Continuous, conical, elevated at perioral region | Uniformly conoid | 63 | 500 | 22 | – | 8.0 | – | 67 | 13 | – | – | 4 | Thorne and Malek [51] |
| 3 B. azadkashmirensis | Open C-shape | Continuous, elevated, anteriorly truncated | Elongate-conoid, terminus hooked | 60 | 400–500 | 23–33 | 4.0–5.0 | 7.0–8.0 | 4.6–5.8 | 63.5–67.6 | 10.4–12 | 1.5–2.0 | 67–80 | 4 | Maqbool et al. [46] |
| 4 B. bambosus | Arcuate | Slightly offset, raised, cupolate | Elongate-conoid, terminus rounded and curved | 46–59 | 410–450 | 30–35 | 4.4–4.8 | 7.5–9.4 | 6.0–12 | 65.3–68.1 | 9.6–12.8 | 3.2 | 62.4–76.8 | 4 | Mohilal et al. [43] |
| 5 B. citri | Spiral | Continuous, cupolate, concave | Hooked, terminus subacute | 46 | 280–310 | 21–23 | – | 7.3–12 | – | 64–68 | 9–10.5 | – | 55–61 | 4 | Edward and Rai [42] |
| 6 B. clavicaudatus | – | Continuous, conical | Conoid, terminus clavate | – | 700 | 31 | 5.7 | 8.5 | – | 60 | 13 | – | – | 4 | Thorne [5] |
| – | Conical, anteriorly flattened | – | 61–74 | 625–740 | 37–49 | 5.2–6.1 | 8.5–12 | 5.1–7.2 | 54.4–62.6 | 11–12.3 | – | – | 4 | Sturhan and Hohberg [52] | |
| 7 B. constrictus | Spiral | Continuous, conical, depression at OA | Elongate-conoid, terminus hooked | 68–85 | 490–550 | 32–37 | 4.6–5.4 | 6.4–8 | 6.4–9.0 | 63.3–67.2 | 11 | 2.5 | – | 4 | Rahman and Ahmad [32] |
| 8 B. cylindricus | – | Offset, elevated | Filiform, terminus rounded | 88–101 | 480–610 | 37–47 | – | 6–7 | 9–13 | 63–64 | 6–8 | 7 | 4 | Dhanachand et al. [36] | |
| 9 B. cynodoni | Close C-shape | Continuous, low, rounded | Elongate, terminus hooked | 54 | 420–490 | 26–30 | 4.5–6.0 | 7–9 | – | 62–65 | 8 | – | 64 | 4 | Fotedar and Mahajan [47] |
| 10 B. filiformis | Arcuate | Slightly offset, cupolate, anteriorly flattened | Elongate, terminus acute | 62 | 460–550 | 24–31 | 5.0–5.6 | 7–9 | – | 63–69 | 9 | 1.5 | 75 | 4 | Husain and Khan [33] |
| 11 B. flexuosus | Spiral | Slightly offset, high, not annulated | Elongate-conoid, curved, terminus pointed | 91–96 | 550–570 | 25–28 | – | 5.7–6.1 | – | 61–65 | 10.5 | 4 | – | 4 | Eroshenko [49] |
| 12 B. hyderi | C-shape | Continuous, cupolate, flat | Elongate-conoid, terminus rounded | – | 440–500 | 23–27 | 4.3–5.0 | 6.8–7.8 | – | 63.6–68.5 | 10–11 | – | 75–80 | 4 | Husain and Khan [53] |
| 13 B. impar | Close C-shape | Continuous, conoid, elevated | Elongate, ventrally arcuate | 96 | 504–600 | 25–32 | 4.8–6.2 | 5–7 | – | 63–66 | 13–14 | 3 | 95 | 4 | Khan and Basir [34] |
| 14 B. innuptus | – | Offset, conical, not annulated | Ventrally arcuate, terminus hooked | 59 | 470–490 | 25–29 | – | 7.3–8.3 | – | 64–66 | 11.5–12.5 | – | – | 4 | Andrassy [37] |
| 15 B. longicaudatus | Slightly ventrally arcuate | Continuous, conical, depression at OA | Elongate, terminus pointed | 88–92 | 430–720 | 25–28 | 7.5–9.2 | 4.8–6.2 | 8.6–12 | 65–74 | 8.5 | 3.4 | 71–74 | 4 | Bina et al. [45] |
| 16 B. minustylus | Slightly curved ventrally | Continuous, rounded | Short, terminus rounded | 59–72 | 960–830 | 48–52 | 5.6–7.4 | 10–14 | 5.3–7.5 | 79–81 | 4.0–4.8 | 3–5 | 99–109 | 4 | Mohilal et al. [43] |
| 17 B. mirus | Ventrally arcuate | Continuous, elevated | Elongate- filiform, terminus rounded | – | 540–572 | 25–32 | 4.8–5.3 | 6.2–7.8 | – | 63–66 | 11–12 | 2 | 100 | 4 | Khan [35] |
| 18 B. modicus | Spiral | Slightly offset, raised, anteriorly-flattened, depression at OA | Hooked, terminus striated and rounded | 51 | 429–487 | 24–31 | 4.2–5.0 | 7.7–9.8 | 4.0–5.5 | 66.5–71 | 9.8–10.2 | 3 | 77–78 | 4 | Lal and Khan [38] |
| 19 B. neosimilis | – | Continuous, narrow, truncate | Uniform, terminus blunt | 51 | 460 | 23 | – | 9 | – | 68 | 10 | – | – | 4 | Geraert [54] |
| 20 B. pakistanensis | C-shape | Slightly offset, elevated, conoid, anteriorly truncated | Elongate-conoid, terminus rounded | 73 | 540–580 | 31–34 | 5.0–5.2 | 7.5–8.0 | – | 67–68 | 11–12 | 4 | – | 4 | Siddiqi [55] |
| 21 B. punici | Slightly ventrally arcuate | Setoff, elevated, wide | Long, filiform, terminus rounded | 73–82 | 450–510 | 37–43 | 4.9–6.6 | 5–7 | 9.6–12 | 58–64 | 7–8 | 7 | – | 4 | Gambhir and Dhanachand [39] |
| 22 B. rafiqi | C-shape | Continuous, cupolate, flat | Elongate, arcuate, terminus rounded | – | 500–600 | 22–35 | 4.7–5.0 | 7.0–10 | – | 65–68 | 8–11 | – | 80–90 | 4 | Husain and Khan [53] |
| 23 B. seshadrii | Open C-shape | Continuous, conoid | Dorsally curved, terminus rounded | – | 410–510 | 28–29 | 4.2–4.9 | 6.0–8.1 | – | 67–68 | 12–13 | 3 | – | 4 | Handoo et al. [56] |
| 24 B. similis | C-shape | Continuous, elevated | Elongate, terminus hooked | 63 | 390–440 | 19–24 | 3.0–4.7 | 5–7 | – | 65–68 | 10–11 | 2 | 75 | 4 | Khan and Basir [48] |
| 25 B. spinocaudatus | Almost straight | Continuous, high and wide | Elongate, terminus rounded or pointed | 49–80 | 600–690 | 25–36 | 6.6–11 | 7.5–13 | 4.6–6.7 | 76–83 | 8.5–10.2 | 5.1 | 62–88 | 4 | Bina et al. [45] |
| 26 B. solomonensis | Almost straight | Slightly offset, narrow, conical, depression at OA | Elongate, terminus rounded or clavate | 80–93 | 550–650 | 24–39 | 4.5–6.0 | 6.3–7.3 | 6.1–9.3 | 53.4–58 | 12–14.5 | – | 85–98 | 4 | Ye and Geraert [40] |
| 27 B. spiralis | Spiral | Slightly offset, truncate | Short, terminus rounded | 45 | 400–490 | 22–27 | 3.4–4.4 | 8.1–10 | – | 62.4–73.2 | 10–12 | – | – | 4 | Egunjobi [41] |
| 28 B. thylactus | Ventrally arcuate | Offset, convex conoid | Hooked, terminus acute | – | 600 | 31 | 5.5 | 8.0 | – | 12.0 | – | – | 4 | Thorne [5] | |
| 29 B. tenuis | Open C-shape | Slightly offset, raised, depression at OA | Hooked, terminus unstriated and rounded | 71 | 480–550 | 25–30 | 4.6–5.2 | 6.3–7.6 | 6.0–7.7 | 65–70 | 11.3–12.5 | 2.5 | 75–84.6 | 4 | Lal and Khan [38] |
| 30 B. teres | Ventrally arcuate | Continuous, conoid-truncate | Ventrally curved, terminus rounded | 71 | 420–560 | 23–35 | – | 6.0–8.5 | 6.5–10 | 54–63 | 10–12 | 5 | 90 | 4 | Nanjappa and Khan [57] |
| 31 B. typicus | Open C-shape | Continuous, truncated, cupolate | Elongate-conoid, terminus rounded | – | 440–580 | 23–27 | 3.9–5.3 | 6–9 | – | 62–68 | 12–14 | – | 90–100 | 6 | Husain and Khan [30] |
| 32 B. volutus | Spiral | Offset, raised, cupolate, anteriorly flattened | Hooked, terminus unstriated and rounded | 35–60 | 450–510 | 20–28 | 4.9–5.7 | 8.7–12 | – | 67–72 | 8.3–9.2 | – | 78 | 4 | Lima and Siddiqi [6] |
| 33 B. zaini | Spiral | Continuous, high, smooth, truncated | Conoid, terminus rounded | 87 | 640–670 | 27–30 | 5.7–7.2 | 7.6–9.5 | 6.5–7.3 | 65.2–67.3 | 12.6 | 4 | – | 6 | Maqbool [31] |
| Species Name | n | L | a | b | c | Stylet Length | Spicule Length | Spicule Morphology | Gubernaculum | Bursa | References | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | B. acurvus | 1 | 460 | 38 | 4.2 | 5 | – | 16 | Cephalated, arcuate | – | Adanal | Jairajpuri [44] |
| 1 | 520 | 37 | 4.4 | 6.1 | 10.5 | 12.5 | – | 5.0 | Adanal | Zeidan and Geraert [50] | ||
| 2 | B. azadkashmirensis | 10 | 400–440 | 25–44 | 4.0–4.6 | 6.6–7.8 | 10.4–11.2 | 12.8–16 | Cephalated, arcuate | 4.0–4.8 | Crenated, adanal | Maqbool, Shahina, and Firoza [46] |
| 3 | B. clavicaudatus | 1 | 635 | 39 | 5.6 | 9.6 | 11.5 | 14 | – | – | – | Sturhan and Hohberg [52] |
| 4 | B. cylindricus | – | – | – | – | – | – | 16 | – | 3–6 | – | Dhanachand, Renubala, and Anandi [36] |
| 5 | B. cynodoni | 4 | 390–450 | 32–35 | 4.5–6.0 | 6–7 | 8 | 10–11 | – | 4 | Crenate, short | Fotedar and Mahajan [47] |
| 6 | B. filiformis | 1 | 460 | 38 | 4 | 6 | 9 | 13 | Paired, cephalated, arcuate | 3 | Crenate, adanal | Husain and Khan [33] |
| 7 | B. flexuosus | – | – | – | – | – | – | 14 | – | – | – | Eroshenko [49] |
| 8 | B. hyderi | 1 | 380 | 22 | 4.1 | 7.0 | 10 | 12 | – | 5 | – | Husain and Khan [53] |
| 9 | B. impar | 6 | 520–560 | 37–47 | 4–6 | 5–6 | 12–14 | 18–20 | Paired, cephalated, arcuate | 7–8 | Adanal | Khan and Basir [34] |
| 10 | B. minustylus | 2 | 660–670 | 52–53 | 5.3–5.8 | 11–15 | 3–4 | 14–16 | Sclerotized and arcuate | 4.8 | – | Mohilal, Anandi, and Dhanachand [43] |
| 11 | B. mirus | 5 | 525–590 | 37–47 | 5–6 | 4–4.8 | 10–11 | 19–22 | Paired, cephalated, arcuate | 9–10 | Crenated | Khan [35] |
| 12 | B. neosimilis | – | – | – | – | – | – | – | Cephalated | 14 | – | Geraert [54] |
| 13 | B. punici | 2 | 470–500 | 34–35 | 5.2–5.8 | 5.0–5.7 | 7–9 | 13–16 | – | 2–3 | – | Gambhir and Dhanachand [39] |
| 14 | B. similis | 1 | 380 | 26 | 4.2 | 5 | 11 | 15 | Cephalated, arcuate | 3 | Crenated | Khan and Basir [48] |
| 15 | B. solomonensis | 6 | 505–590 | 25.5–35.0 | 4.2–5.4 | 5.7–6.6 | 12–13 | 14–17 | Paired, cephalated | – | Adanal | Ye and Geraert [40] |
| 16 | B. thylactus | 1 | 500 | 33 | 5.0 | 7.2 | – | – | – | – | – | Thorne [5] |
| 14 | 535–662 | 30.2–42.3 | 4.8–6.3 | 8.2–11.5 | 10.9–12.4 | 8.8–11.1 | – | – | – | Deimi and Mitkowski [58] | ||
| 17 | B. tenuis | 10 | 440–530 | 25.0–31.6 | 4.5–5.0 | 6.5–6.9 | 10.8–12.0 | 12–14.2 | Arcuate | 5.0 | – | Lal and Khan [38] |
| 18 | B. teres | – | – | – | – | – | – | 13 | Cephalated | 6 | Crenate | Nanjappa and Khan [57] |
| 19 | B. typicus | 5 | 520–560 | 33–47 | 4.5–5.3 | 7 | 13–14 | 18–20 | Paired, cephalated | 3–5 | Adanal | Husain and Khan [30] |
| 20 | B. volutus | 1 | 440 | 28 | 4.9 | 9.6 | 9.0 | 14–15 | Cephalated, arcuate | 4.8–5.1 | Adanal | Lima and Siddiqi [6] |
| 21 | B. zaini | 9 | 480–600 | 30–33 | 5.3–6.2 | 7.5–9 | 12.6 | 16–18 | Paired, arcuate | 6.7 | Short, adanal | Maqbool [31] |
3.1.2. Description of B. thylactus Thorne, 1941
| Canada (Present Study) | Thorne [5] | Geraert [59] | Rashid et al. [60] | Lal and Khan [38] | Deimi and Mitkowski [58] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Populations | 61 | 40 | 50 | 62 | USA 1 | Belgium | Brazil | India | Iran | ||
| Characters | Females | Males | Females | Females | Females | Female | Females 2 | Female | Females 3 | Females | Male |
| n | 15 | 5 | 10 | 10 | 10 | – | – | 1 | – | 12 | 14 |
| Body length | 525.3 ± 31.5 (483–602.0) | 477.3 ± 23.7 (450.0–500.0) | 501.3 ± 32.7 (446–552) | 527.1 ± 28.9 (490.0–576.0) | 507.5 ± 34.3 (450.0–558.0) | 600 | 380–590 | 390 | 430–580 | 598 (535–662) | 544 (501–614) |
| a | 32.5 ± 2.3 (28.5–35.9) | 38.5 ± 1.9 (36.9–41.2) | 33.0 ± 22.1 (37.2–31.5) | 31.8 ± 2.5 (27.3–34.4) | 32.8 ± 2.8 (27.8–37.9) | 31 | 21–39 | 25 | 23–29 | 35.4 (30.2–42.3) | 41.3 (37.6–47.2) |
| b | 4.8 ± 0.3 (4.3–5.5) | 4.4 ± 0.2 (4.2–4.6) | 4.7 ± 7.5 (4.5–4.9) | 4.9 ± 0.3 (4.6–5.5) | 4.8 ± 0.3 (4.3–5.3) | 5.5 | 4–6.4 | 4.4 | 4.8–6.2 | 5.2 (4.8–6.3) | 5.6 (4.6–5.8) |
| c | 9.0 ± 1.1 (7.0–10.2) | 6.8 ± 0.2 (6.6–7.0) | 8.1 ± 5.1 (8.1–7.7) | 8.7 ± 1.1 (7.0–10.1) | 8.0 ± 0.9 (6.6–9.1) | 8 | 5.6–8.7 | 7.4 | 6.7–8.2 | 9.7 (8.2–11.5) | 8.8 (7.2–9.4) |
| c′ | 5.9 ± 0.8 (5.0–8.0) | 7.9 ± 0.7 (6.9–8.4) | 6.4 ± 9.2 (6.1–6.7) | 6.2 ± 0.8 (5.2–7.7) | 6.5 ± 1.0 (5.0–8.1) | – | – | 6.4 | 5.9–7.8 | 6.4 (4.6–7.9) | 7.4 (6.4–9.1) |
| V | 67.0 ± 1.3 (65.0–69.0) | 31.2 ± 1.7 (29.2–32.4) | 66.2 ± 1.1 (65.0–68.0) | 66.8 ± 1.5 (65.0–69.0) | 66.5 ± 1.4 (65.0–69.0) | – | 62–71.5 | 65 | 64–68 | 64.1 (61.8–65.9) | – |
| MB | 41.9 ± 4.2 (37.5–48.6) | 56 | – | – | – | – | – | – | – | – | – |
| G1 | 25.7 ± 2.2 (23.1–30.6) | – | 22.0 ± 24.8 (22.2–21.2) | 22.4 ± 1.9 (21.1–23.8) | 21.5 ± 1.7 (19.5–22.6) | – | – | – | – | – | – |
| Lip height | 2.8 ± 0.2 (2.4–3.0) | 3.0 ± 0.1 (3.0–3.1) | 2.9 ± 0.2 (2.6–3.3) | 2.7 ± 0.2 (2.4–2.9) | 3.0 ± 0.2 (2.5–3.3) | – | – | – | – | – | – |
| Lip width | 5.5 ± 0.3 (5.1–6.0) | 5.3 ± 0.3 (5.0–5.7) | 5.9 ± 0.2 (5.4–6.0) | 5.4 ± 0.3 (5.1–5.9) | 5.6 ± 0.3 (5.0–5.9) | – | – | – | – | – | – |
| Stylet length | 9.3 ± 0.8 (8.2–10.7) | 9.0 ± 1.2 (7.5–10.3) | 9.4 ± 0.5 (8.5–10.6) | 9.6 ± 0.8 (8.6–10.8) | 9.6 ± 0.3 (9.2–10.0) | 12 | 8.5–12 | 10 | 10.5–12 | 10.9 (10.1–12.3) | 11.1 (10.9–12.4) |
| Anterior end to excretory pore | 84.8 ± 5.1 (74.0–92.0) | 82.3 ± 3.9 (77.0–86.0) | 85.1 ± 4.5 (79.0–92.0) | 88.3 ± 4.2 (85.0–96.0) | 82.2 ± 3.1 (79.0–87.0) | – | – | – | – | 82.6 (80.7–87.8) | 80.2 (78.7–84.1) |
| Pharynx length | 109.1 ± 3.9 (103.0–117.0) | 109.5 ± 1.7 (108.0–111.0) | 107.3 ± 4.4 (99.0–113.0) | 107.2 ± 6.0 (99.0–117.0) | 105.0 ± 5.1 (99.0–113.0) | – | – | 88.5 | – | 108 (100–119) | 101 (99–107) |
| Maximum body width | 16.3 ± 1.9 (13.8–20.0) | 12.4 ± 0.6 (12.0–13.3) | 15.2 ± 1.5 (12.0–17.5) | 16.6 ± 1.2 (15.0–18.0) | 15.6 ± 1.4 (13.5–18.0) | – | – | 16 | – | 15.7 (13.4–17.5) | 12.6 (12.1–14.3) |
| Vulva body width | 14.8 ± 1.6 (13.0–18.5) | – | 14.4 ± 1.3 (12.0–17.0) | 15.3 ± 1.1 (14.0–17.0) | 14.4 ± 1.1 (13.0–16.0) | – | – | – | – | – | – |
| Post uterine sac length | 11.1 ± 0.9 (9.7–13.0) | – | 10.2 ± 1.1 (9.0–12.0) | 9.7 ± 1.1 (8.2–11.6) | 9.4 ± 0.8 (8.2–10.8) | – | – | – | – | 10.7 (8.5–14.6) | – |
| Distance from vulva to anus | 111.6 ± 9.0 (102.0–131.0) | – | 106.9 ± 4.2 (101.0–115.0) | 112.7 ± 4.1 (106.0–118.0) | 104.8 ± 6.3 (101.0–119.0) | – | – | – | – | 107.8 (98–119) | – |
| Anal/cloacal body width | 10.2 ± 1.4 (8.8–13.0) | 8.9 ± 1.0 (8.1–10.3) | 9.7 ± 0.7 (9.0–10.8) | 10.0 ± 0.9 (8.1–10.9) | 9.8 ± 0.8 (9.0–11.0) | – | – | – | – | – | – |
| Spicule length | – | 17.0 ± 1.4 (16.0–19.0) | – | – | – | – | – | – | – | – | 9.5 (8.8–11.1) |
| Tail length | 59.4 ± 9.5 (50.0–77.0) | 70.3 ± 1.7 (68.0–72.0) | 61.9 ± 6.4 (55.0–72.0) | 61.5 ± 6.3 (52.0–71.0) | 63.9 ± 7.7 (55.0–75.0) | – | – | 53 | – | 67 (59–82) | 68.6 (66.8–70.7) |
3.1.3. Description of B. volutus Lima and Siddiqi, 1963
3.2. Habitat and Locality
3.3. Distribution and Associated Hosts of Boleodorus Species
| Character | Canadian Populations | Original Description | |
|---|---|---|---|
| Populations | 61 | 40 | Lima and Siddiqi [6] |
| n | 10 | 8 | 16 |
| Body length | 547.4 ± 31.5 (501.0–581.0) | 533.0 ± 30.7 (487.0–577.0) | 480 (450–510) |
| a | 27.4 ± 2.5 (24.5–31.3) | 28.9 ± 2.6 (24.9–32.1) | 26 (20–28) |
| b | 4.6 ± 0.2 (4.2–4.8) | 4.5 ± 0.2 (4.2–4.9) | 5.3 (4.9–5.7) |
| c | 9.6 ± 0.8 (8.5–10.6) | 10.0 ± 1.0 (8.8–11.5) | 10 (8.7–12) |
| c′ | 5.4 ± 0.8 (4.3–6.3) | 5.2 ± 0.5 (4.6–5.8) | – |
| V | 68.9 ± 0.7 (68.0–70.0) | 69.0 ± 1.2 (68.0–71.0) | 69 (67–72) |
| Lip height | 2.7 ± 0.2 (2.4–3.0) | 2.3 ± 0.3 (2.1–2.8) | – |
| Lip width | 5.9 ± 0.2 (5.4–6.3) | 5.6 ± 0.1 (5.5–5.8) | – |
| Stylet length | 8.5 ± 0.4 (8.0–9.3) | 9.2 ± 0.6 (8.2–9.8) | 8.7 (8.3–9.2) |
| Anterior end to excretory pore | 90.7 ± 4.2 (86.0–97.0) | 88.4 ± 3.8 (85.0–95.0) | 78 |
| Pharynx | 118.5 ± 4.7 (109.0–125.0) | 119.0 ± 7.3 (108.0–129.0) | – |
| Maximum body width | 20.1 ± 2.4 (16.0–22.7) | 18.6 ± 2.3 (16.0–22.4) | – |
| Vulva body width | 17.4 ± 1.7 (15.0–19.8) | 16.9 ± 1.4 (15.0–19.0) | – |
| Post uterine sac length | 10.4 ± 1.7 (7.2–12.6) | 9.6 ± 1.2 (8.0–12.1) | – |
| Distance from vulva to anus | 118.1 ± 8.1 (105.0–129.0) | 110.5 ± 7.3 (102.0–121.0) | – |
| Anal body width | 10.9 ± 1.3 (9.5–13.2) | 10.3 ± 0.7 (9.2–11.0) | – |
| Tail length | 57.8 ± 3.0 (52.0–62.0) | 53.9 ± 4.8 (46.0–59.0) | – |
| Species Name | Country | Host | References | |
|---|---|---|---|---|
| 1 | B. acurvus | Nigeria | Saccharum officinarum L. | Jairajpuri [44] |
| Sudan | Citrus limon | Zeidan and Geraert [50] | ||
| 2 | B. acutus | USA | – | Thorne and Malek [51] |
| 3 | B. azadkashmirensis | Pakistan | Allium cepa, Zea mays Fragaria ananassa | Maqbool, Shahina, and Firoza [46] |
| 4 | B. bambosus | India | Bambusa tuida | Mohilal, Anandi, and Dhanachand [43] |
| 5 | B. citri | India | Citrus reticulata | Edward and Rai [42] |
| 6 | B. clavicaudatus | USA | Alfalfa crowns | Thorne [5] |
| German | Loamy soil from meadows | Sturhan and Hohberg [52] | ||
| 7 | B. constrictus | India | Carica papaya | Rahman and Ahmad [32] |
| 8 | B. cylindricus | India | Saccharum officinarum L. | Dhanachand, Renubala, and Anandi [36] |
| India | Brassica oleracea | Hassan and Ahangar [67] | ||
| 9 | B. cynodoni | India | Cynodon dactylon Pers. | Fotedar and Mahajan [47] |
| 10 | B. filiformis | India | Solanum melongena | Husain and Khan [33] |
| 11 | B. flexuosus | Russia | – | Eroshenko [49] |
| 12 | B. hyderi | India | Mangifera indica L. | Husain and Khan [53] |
| 13 | B. impar | India | Cynodon dactylon Pers. | Khan and Basir [34] |
| 14 | B. innuptus | Hungary | – | Andrassy [37] |
| 15 | B. longicaudatus | India | Morus alba L. | Bina, Mohilal, Pramodini, and Majur-Shah [45] |
| 16 | B. minustylus | India | Banana roots | Mohilal, Anandi, and Dhanachand [43] |
| 17 | B. mirus | India | Cynodon dactylon Pers. | Khan [35] |
| 18 | B. modicus | India | Populus sp. and Myrica sapida | Lal and Khan [38] |
| 19 | B. neosimilis | USA | - | Geraert [54] |
| 20 | B. pakistanensis | Pakistan | Pinus excelsa Wall. | Siddiqi [55] |
| Egypt | Polypogon monspeliensis L., Solanum nigrum L., | Ibrahim et al. [3] | ||
| 21 | B. punici | India | Punica granatum | Gambhir and Dhanachand [39] |
| 22 | B. rafiqi | India | Pyrus communis L. | Husain and Khan [53] |
| 23 | B. seshadrii | India | Glycine max L. | Handoo et al. [56] |
| 24 | B. similis | India | Plumeria acutifolia Poir. | Khan and Basir [48] |
| 25 | B. spinocaudatus | India | Morus alba L. | Bina, Mohilal, Pramodini, and Majur-Shah [45] |
| 26 | B. solomonensis | Solomon Islands | Moist soil in tropical forest, host unknown | Ye and Geraert [40] |
| 27 | B. spiralis | New Zealand | Dominant vegetation composed of Leptospermum scoparium, Weinmannia racemose, and Psuedopanax arboreum | Egunjobi [41] |
| 28 | B. thylactus | USA | Cultivated soil, alfalfa crown roots | Thorne [5] |
| Belgium | Meadows, arable land | Geraert [59] | ||
| Brazil | Theobroma cacao | Rashid et al. [60] | ||
| India | Pinus sp., Artocarpus integrifolia | Lal and Khan [38] | ||
| Spain | Natural plant communities | Castillo et al. [13] | ||
| Iran | Grapes | Karegar et al. [68] Deimi and Mitkowski [58] | ||
| Afghanistan | Clover | Asghari et al. [63] | ||
| Germany | Arable soil, cherry tree plantation | Sturhan and Hohberg [52] | ||
| Iran | Polianthes tuberosa | Husseinvand et al. [69] | ||
| Slovakia | Solidago gigantea | Čerevková et al. [12] | ||
| 29 | B. tenuis | India | Casurina equsitifolia | Lal and Khan [38] |
| 30 | B. teres | India | – | Nanjappa and Khan [57] |
| 31 | B. typicus | India | Narcissus sp. | Husain and Khan [30] |
| 32 | B. volutus | UK | Grass species | Lima and Siddiqi [6] |
| Netherlands | – | Bongers [61] | ||
| Poland | – | Brzeski [62] | ||
| Afghanistan | Potato, tomato | Asghari et al. [63] | ||
| Germany | Meadow soil | Sturhan and Hohberg [52] | ||
| 33 | B. zaini | Pakistan | Citrus aurantium | Maqbool [31] |
3.4. Molecular Characterization of B. thylactus and B. volutus with Phylogenetic Relationships of Boleodorus with Related Genera
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munawar, M.; Yevtushenko, D.P.; Castillo, P. Integrative taxonomy, distribution, and host associations of Geocenamus brevidens and Quinisulcius capitatus from southern Alberta, Canada. J. Nematol. 2021, 53. [Google Scholar] [CrossRef]
- Munawar, M.; Yevtushenko, D.P.; Palomares-Rius, J.E.E.; Castillo, P. Species diversity of pin nematodes (Paratylenchus spp.) from potato growing regions of southern Alberta, Canada. Plants 2021, 10, 188. [Google Scholar] [CrossRef]
- Ibrahim, I.; Handoo, Z.; El-Sherbiny, A. A survey of phytoparasitic nematodes on cultivated and non-cultivated plants in Northwestern Egypt. J. Nematol. 2000, 32, 478. [Google Scholar] [PubMed]
- Forge, T.A.; Larney, F.J.; Kawchuk, L.M.; Pearson, D.C.; Koch, C.; Blackshaw, R.E. Crop rotation effects on Pratylenchus neglectus populations in the root zone of irrigated potatoes in southern Alberta. Can. J. Plant Pathol. 2015, 37, 363–368. [Google Scholar] [CrossRef]
- Thorne, G. Some nematodes of the family Tylenchidae which do not possess a valvular median esophageal bulb. Great Basin Nat. 1941, 2, 37–85. [Google Scholar]
- Lima, M.; Siddiqi, M.R. B. volutus n. sp.(Nematoda: Nothotylenchinae) found in soil about grass roots in England. Nematologica 1963, 9, 19–23. [Google Scholar] [CrossRef]
- Geraert, E. The Tylenchidae of the World: Identification of the Family Tylenchidae (Nematoda); Academia Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Geraert, E. Tylenchidae in agricultural soils. Man. Agric. Nematol. 1991, 795–825. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; De Goede, R.; Freckman, D.W.; Georgieva, S. Feeding habits in soil nematode families and genera—an outline for soil ecologists. J. Nematol. 1993, 25, 315. [Google Scholar]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef]
- Čerevková, A.; Miklisová, D.; Bobuľská, L.; Renčo, M. Impact of the invasive plant Solidago gigantea on soil nematodes in a semi-natural grassland and a temperate broadleaved mixed forest. J. Helminthol. 2020, 94. [Google Scholar] [CrossRef]
- Castillo, P.; País, M.; Barcina, A.G. Nematodos de la Familia Tylenchidae Orley, 1880 (Nematoda: Tylenchida), en la sierra de Cazorla, España. Zool. Baetica 1991, 2, 137–161. [Google Scholar]
- Jenkins, W. A rapid centrifugal-flotation technique for separating nematodes from soil. Plant Dis. Report. 1964, 48, 692. [Google Scholar]
- Seinhorst, J. A rapid method for the transfer of nematodes from fixative to anhydrous glycerin. Nematologica 1959, 4, 67–69. [Google Scholar] [CrossRef]
- De Grisse, A.T. Redescription ou Modifications de Quelques Technique Utilis [a] es Dan L’etude des n [a] Ematodes Phytoparasitaires; Mededelingen Rijksfakulteit Landbouwwetenschappen: Ghent, Belgium, 1969. [Google Scholar]
- Maria, M.; Powers, T.; Tian, Z.; Zheng, J. Distribution and description of criconematids from Hangzhou, Zhejiang Province, China. J. Nematol. 2018, 50, 183–206. [Google Scholar]
- Holterman, M.; van der Wurff, A.; van den Elsen, S.; van Megen, H.; Bongers, T.; Holovachov, O.; Bakker, J.; Helder, J. Phylum-wide analysis of SSU rDNA reveals deep phylogenetic relationships among nematodes and accelerated evolution toward crown clades. Mol. Biol. Evol. 2006, 23, 1792–1800. [Google Scholar] [CrossRef] [PubMed]
- Holterman, M.; Rybarczyk, K.; Van den Elsen, S.; Van Megen, H.; Mooyman, P.; Santiago, R.P.; Bongers, T.; Bakker, J.; Helder, J. A ribosomal DNA-based framework for the detection and quantification of stress-sensitive nematode families in terrestrial habitats. Mol. Ecol. Resour. 2008, 8, 23–34. [Google Scholar] [CrossRef]
- Ferris, V. Variation in spacer ribosomal DNA in some cyst-forming species of plant parasitic nematodes. Fundam. Appl. Nematol. 1993, 16, 177–184. [Google Scholar]
- Curran, J.; Driver, F.; Ballard, J.; Milner, R. Phylogeny of Metarhizium: Analysis of ribosomal DNA sequence data. Mycol. Res. 1994, 98, 547–552. [Google Scholar] [CrossRef]
- Qing, X.; Bert, W. Redefinition of genus Malenchus Andrássy, 1968 (Tylenchomorpha: Tylenchidae) with additional data on ecology. J. Nematol. 2017, 49, 189. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree v1. 4.2, a Graphical Viewer of Phylogenetic Trees. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 21 April 2021).
- Geraert, E. Morphometric relations in nematodes. Nematologica 1968, 14, 171–183. [Google Scholar] [CrossRef]
- Husain, S.I.; Khan, A.M. Paurodontella n. gen. and three new species of nematodes from North India (Nematoda: Neotylenchidae). Nematologica 1968, 13, 493–500. [Google Scholar] [CrossRef]
- Maqbool, M. Three new species of the super family Neotylenchoidea (Nematoda: Tylenchida) from Pakistan. J. Nematol. 1982, 14, 317. [Google Scholar]
- Rahman, P.F.; Ahmad, I. Two new species of Tylenchoidea from Uttar Pradesh. Indian J. Nematol. 1995, 25, 136–141. [Google Scholar]
- Husain, S. Boleodorus filiformis nomen novum for B. acutus Husain and Khan, 1974. Indian J. Nematol. 1977, 5, 246–247. [Google Scholar]
- Khan, E.; Basir, M. Boleodorus impar n. sp.(Nematoda: Tylenchida) from India. Proc. Helminthol. Soc. Wash. 1964, 31, 187–190. [Google Scholar]
- Khan, E. Boleodorus mirus n. sp. (Tylenchida: Boleodorinae n. subfam.) from Kufri, Simla (HP) India, with a key to the species of the genus Boleodorus Thorne, 1941. Zool. Anz. 1964, 173, 336–341. [Google Scholar]
- Dhanachand, C.; Renubala, K.; Anandi, Y. Three new species of Tylenchida associated with sugarcane from Manipur. Curr. Nematol. 1993, 4, 45–52. [Google Scholar]
- Andrassy, I. Zur taxonomie der Neotylenchiden. Nematologica 1961, 6, 25–36. [Google Scholar] [CrossRef]
- Lal, A.; Khan, E. Nematodes Associated with Forest Trees in Northern India-Species of Boleodorus Thorne, 1941; My Forest 24: 301-306; Diversity of Plant and Soil Nematodes in Uttarakhand: Dehradun, India, 1988; p. 267. [Google Scholar]
- Gambhir, R.; Dhanachand, C. Nematodes of fruit plants in Manipur–five new species of tylenchids (Nematoda: Tylenchida). Indian J. Nematol. 1996, 26, 197–207. [Google Scholar]
- Ye, W.; Geraert, E. Plant parasitic nematodes from the Solomon Islands with a description of Boleodorus solomonensis. Nematologica 1997, 43, 431–454. [Google Scholar] [CrossRef]
- Egunjobi, O.A. Three new species of nematodes from New Zealand. N. Z. J. Sci. 1968, 11, 488–497. [Google Scholar]
- Edward, J.; Rai, B. Plant parasitic nematodes association with hill orange (Citrus reticulata Blanco) in Sikkim. Allahabad Farmer 1970, 44, 251–254. [Google Scholar]
- Mohilal, N.; Anandi, Y.; Dhanachand, C. Two new species of Boleodorus Thorne, 1941 and male report of Neopsilenchus affinis. Curr. Nematol. 1997, 8, 17–22. [Google Scholar]
- Shamim Jairajpuri, M. Three new species of nematodes from sugarcane fields in Nigeria. Rev. Nématol. 1982, 5, 241–246. [Google Scholar]
- Bina, L.; Mohilal, N.; Pramodini, M.; Shah, M.M. Two new species of Boleodorus (Nematoda: Tylenchida) from Manipur, India. Bioscan 2012, 7, 115–118. [Google Scholar]
- Maqbool, M.; Shahina, F.; Firoza, K. Description of Boleodorus azadkashmirensis n. sp.(Nematoda: Tylenchidae) and observation on Filenchus vulgaris (Brzeski, 1963) Lownsbery & Lownsbery, 1985 from Pakistan. Pak. J. Nematol. 1990, 8, 43–48. [Google Scholar]
- Fotedar, D.; Mahajan, R. Two new nematode species (Nothotylenchidae) from Kashmir. Indian J. Nematol. 1972, 2, 169–172. [Google Scholar]
- Khan, E.; Basir, M. Boleodorus similis n. sp.(Nematoda: Nothotylenchinae) from India. Z. Parasitenkd. 1963, 23, 121–123. [Google Scholar] [CrossRef]
- Eroshenko, A. A new species of the genus Boleodorus (Nematoda, Nothotylenchidae) from the spruce rhizosphere in the Primorsky Region. Zool. Zhurnal 1982, 61, 135–137. [Google Scholar]
- Zeidan, A.; Geraert, E. The genera Filenchus Andrássy, 1954, Sakia Khan, 1964, Boleodorus Thorne, 1941 and Basiria Siddiqi, 1959 (Nemata: Tylenchida) from Sudan. Nematologica 1991, 37, 185–212. [Google Scholar]
- Thorne, G.; Malek, R.B. Nematodes of the northern Great Plains. Part I. Tylenchida (Nemata: Secernentea). Tech. Bull. S. Dak. Agric. Exp. Stn 1968, 31, 111. [Google Scholar]
- Sturhan, D.; Hohberg, K. Nematodes of the order Tylenchida in Germany–the non-phytoparasitic species. Soil Org. 2016, 88, 19–41. [Google Scholar]
- Husain, S.I.; Khan, A. Two new species of Boleodorus Thome, 1941 (Nematoda: Neotylenchidae) from India. Proc. Helm. Soc. Wash. 1965, 32, 176–179. [Google Scholar]
- Geraert, E. Observations on the genera Boleodorus and Boleodoroides (Nematoda: Tylenchida). Nematologica 1971, 17, 263–276. [Google Scholar] [CrossRef]
- Siddiqi, M. Boleodorus pakistanensis n. sp.(Nematoda: Tylenchida), found associated with pine roots in Abbottabad, Pakistan. Sci. Cult. 1963, 29, 562–563. [Google Scholar]
- Handoo, Z.A.; Kantor, M.R.; Khan, E. Description of seven new species and one new record of plant-parasitic nematodes (Nematoda: Tylenchida) associated with economically important crops of Kashmir valley, Jammu and Kashmir (Part-1 of the series). Pak. J. Nematol. 2020, 38. [Google Scholar] [CrossRef]
- Nanjappa, C.; Khan, E. Paurodontus indicus sp. nov. and Boleodorus teres sp. nov.(Nematoda: Tylenchida) from India. Bull. Entomol. 1970, 11, 138–142. [Google Scholar]
- Deimi, A.M.; Mitkowski, N. Nematodes associated with vineyards throughout Markazi province (Arak), Iran. Australas. Plant Pathol. 2010, 39, 571–577. [Google Scholar] [CrossRef]
- Geraert, E. On some Tylenchidae and Neotylenchidae from Belgium with the description of a new species, Tylenchorhynchus microdorus. Nematologica 1966, 12, 409–416. [Google Scholar] [CrossRef]
- Rashid, F.; Geraert, E.; Sharma, R. Seven species of Tylenchida from Brazil with description of a new species (Nematoda: Tylenchoidea). Nematol. Mediterr. 1987, 15, 29–45. [Google Scholar]
- Bongers, T. De Nematoden van Nederland: Een Identificatietabel voor de in Nederland Aangetroffen Zoetwater-en Bodembewonende Nematoden; Stichting Uitgeverij Koninklijke Nederlandse Natuurhistorische Vereniging: Voorschoten, The Netherlands, 1988. [Google Scholar]
- Brzeski, M.W. Nematodes of Tylenchina in Poland and Temperate Europe; Muzeum i Instytutu Zoologii, Polska Akademia Nauk (MiIZ PAN): Warsaw, Poland, 1998. [Google Scholar]
- Asghari, R.; Pourjam, E.; Mohamadi Goltapeh, E.; Latifi, A. Plant-parasitic nematodes from Afghanistan with discussion on the taxonomic status of Merlinius neohexagrammus Ivanova, 1978 (Nematoda: Dolichodoridae). J. Agric. Sci. Technol. 2012, 14, 1397–1404. [Google Scholar]
- Sheikh, J.H.; Chishti, M.; Rasheed, M.; Dar, S.A.; Lal, E.P.; Mohiuddin, D. Population analysis of the genera buildup on some commercially important vegetable crops grown in Kashmir Valley. J. Parasit. Dis. 2016, 40, 877–880. [Google Scholar] [CrossRef][Green Version]
- Pan, F.J.; Xu, Y.L.; McLaughlin, N.B.; Xue, A.G.; Yu, Q.; Han, X.-Z.; Liu, W.; Zhan, L.L.; Zhao, D.; Li, C.J. Response of soil nematode community structure and diversity to long-term land use in the black soil region in China. Ecol. Res. 2012, 27, 701–714. [Google Scholar] [CrossRef]
- Yeates, G.W. Nematodes as soil indicators: Functional and biodiversity aspects. Biol. Fertil. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]
- Hassan, J.; Ahangar, M. Phytonematode infestation on Brassica oleracea with special emphasis on Boleodorus cylindricus Dhanachand, Renubala & Anandi, 1993. Int. J. Adv. Educ. Res. 2018, 3, 157–159. [Google Scholar]
- Karegar, A.; Geraert, E.; Kheiri, A. Tylenchs associated with Grapevine in the province of Hamadan, Iran. Med. Fac. Landbouww. Univ. Gent. 1995, 6O/3b, 1063–1086. [Google Scholar]
- Husseinvand, M.; Abdollahi, M.; Karegar, A. Description of some nematode species of Tylenchidae, associated with Polianthes tuberosa from Iran. J. Agric. Sci. Technol. 2016, 18, 1953–1966. [Google Scholar]
- Giblin-Davis, R.; Mundo-Ocampo, M.; Baldwin, J.; Norden, B.; Batra, S. Description of Bursaphelenchus abruptus n. sp.(Nemata: Aphelenchoididae), an associate of a digger bee. J. Nematol. 1993, 25, 161. [Google Scholar] [PubMed]
- Ritzema Bos, J. De bloemkoolziekte der aardbeien, veroorzaakt door Aphelenchus fragariae nov. spec. Voorloopige mededeeling. Maanblad Nat. 1890, 16, 107–117. [Google Scholar]
- Christie, J. Recent observations on the strawberry dwarf neaiatode in Massachusetts. Plant Dis. Rep. 1932, 16, 113–114. [Google Scholar]
- Tomalak, M. Bursaphelenchus trypophloei sp. n. (Nematoda: Parasitaphelenchinae)–an associate of the bark beetle, Trypophloeus asperatus (Gyll.)(Coleoptera: Curculionidae, Scolytinae), in aspen, Populus tremula L. Nematology 2011, 13, 619–636. [Google Scholar] [CrossRef]
- Mamiya, Y.; Enda, N. Bursaphelenchus mucronatus n. sp. (Nematoda: Aphelenchoididae) from pine wood and its biology and pathogenicity to pine trees. Nematologica 1979, 25, 353–361. [Google Scholar] [CrossRef]
- Karegar, A.; Geraert, E. The genus Basiria Siddiqi, 1959 (Nematoda: Tylenchidae) I. Introduction and species with two lateral lines. Nematologica 1997, 43, 327–339. [Google Scholar] [CrossRef]
- Karegar, A.; Geraert, E. The genus Basiria Siddiqi, 1959 (Nematoda: Tylenchidae) IV. General discussion, genus diagnosis and key to the species. Nematologica 1998, 44, 1–13. [Google Scholar] [CrossRef]
- Qing, X.; Pereira, T.J.; Slos, D.; Couvreur, M.; Bert, W. A new species of Malenchus (Nematoda: Tylenchomorpha) with an updated phylogeny of the Tylenchidae. Nematology 2018, 20, 815–836. [Google Scholar] [CrossRef]
- Qing, X.; Decraemer, W.; Claeys, M.; Bert, W. Molecular phylogeny of Malenchus and Filenchus (Nematoda: Tylenchidae). Zool. Scr. 2017, 46, 625–636. [Google Scholar] [CrossRef]
- Pereira, T.J.; Qing, X.; Chang, K.F.; Mundo-Ocampo, M.; Cares, J.E.; Ragsdale, E.J.; Nguyen, C.N.; Baldwin, J.G. Phylogeny and biogeography of the genus Cephalenchus (Tylenchomorpha, Nematoda). Zool. Scr. 2017, 46, 506–520. [Google Scholar] [CrossRef]
- Van den Berg, E.; Tiedt, L.R.; Subbotin, S.A. Morphological and molecular characterisation of several Paratylenchus Micoletzky, 1922 (Tylenchida: Paratylenchidae) species from South Africa and USA, together with some taxonomic notes. Nematology 2014, 16, 323–358. [Google Scholar] [CrossRef]
- Qing, X.; Bert, W. Family Tylenchidae (Nematoda): An overview and perspectives. Org. Divers. Evol. 2019, 19, 391–408. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munawar, M.; Yevtushenko, D.P.; Castillo, P. Overview of the Genus Boleodorus and First Reports of Boleodorus thylactus and B. volutus from Southern Alberta, Canada. Animals 2021, 11, 1760. https://doi.org/10.3390/ani11061760
Munawar M, Yevtushenko DP, Castillo P. Overview of the Genus Boleodorus and First Reports of Boleodorus thylactus and B. volutus from Southern Alberta, Canada. Animals. 2021; 11(6):1760. https://doi.org/10.3390/ani11061760
Chicago/Turabian StyleMunawar, Maria, Dmytro P. Yevtushenko, and Pablo Castillo. 2021. "Overview of the Genus Boleodorus and First Reports of Boleodorus thylactus and B. volutus from Southern Alberta, Canada" Animals 11, no. 6: 1760. https://doi.org/10.3390/ani11061760
APA StyleMunawar, M., Yevtushenko, D. P., & Castillo, P. (2021). Overview of the Genus Boleodorus and First Reports of Boleodorus thylactus and B. volutus from Southern Alberta, Canada. Animals, 11(6), 1760. https://doi.org/10.3390/ani11061760