
The Effect of Non-Circadian Photoperiod on Growth and Puberty Onset of Brook Trout Salvelinus fontinalis Mitchill



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
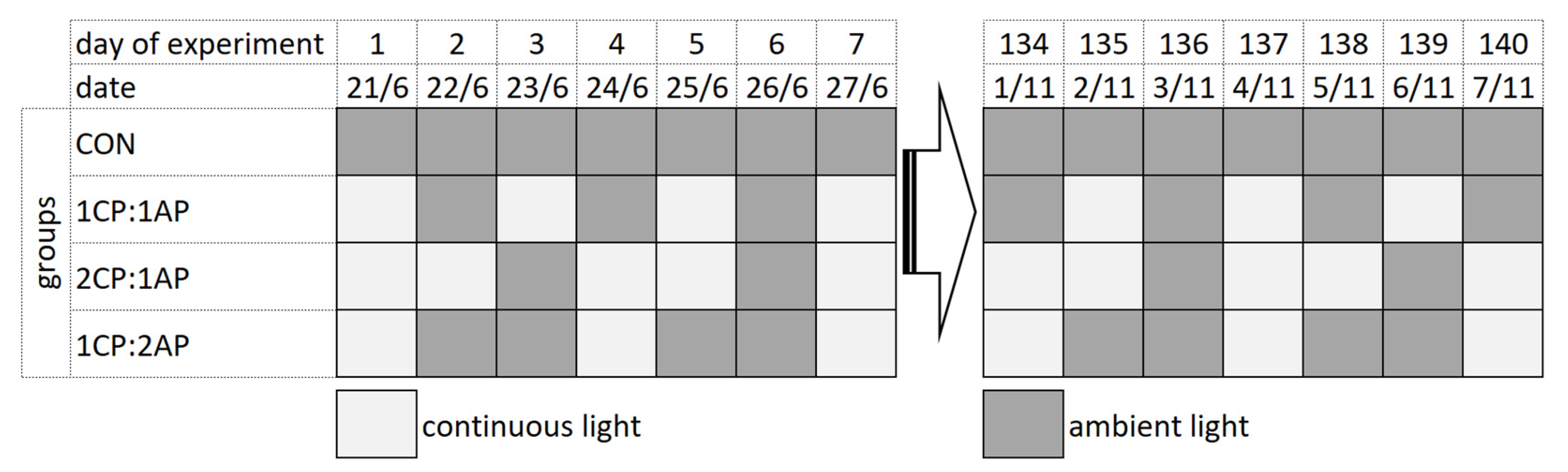
2.1. Experimental Protocol
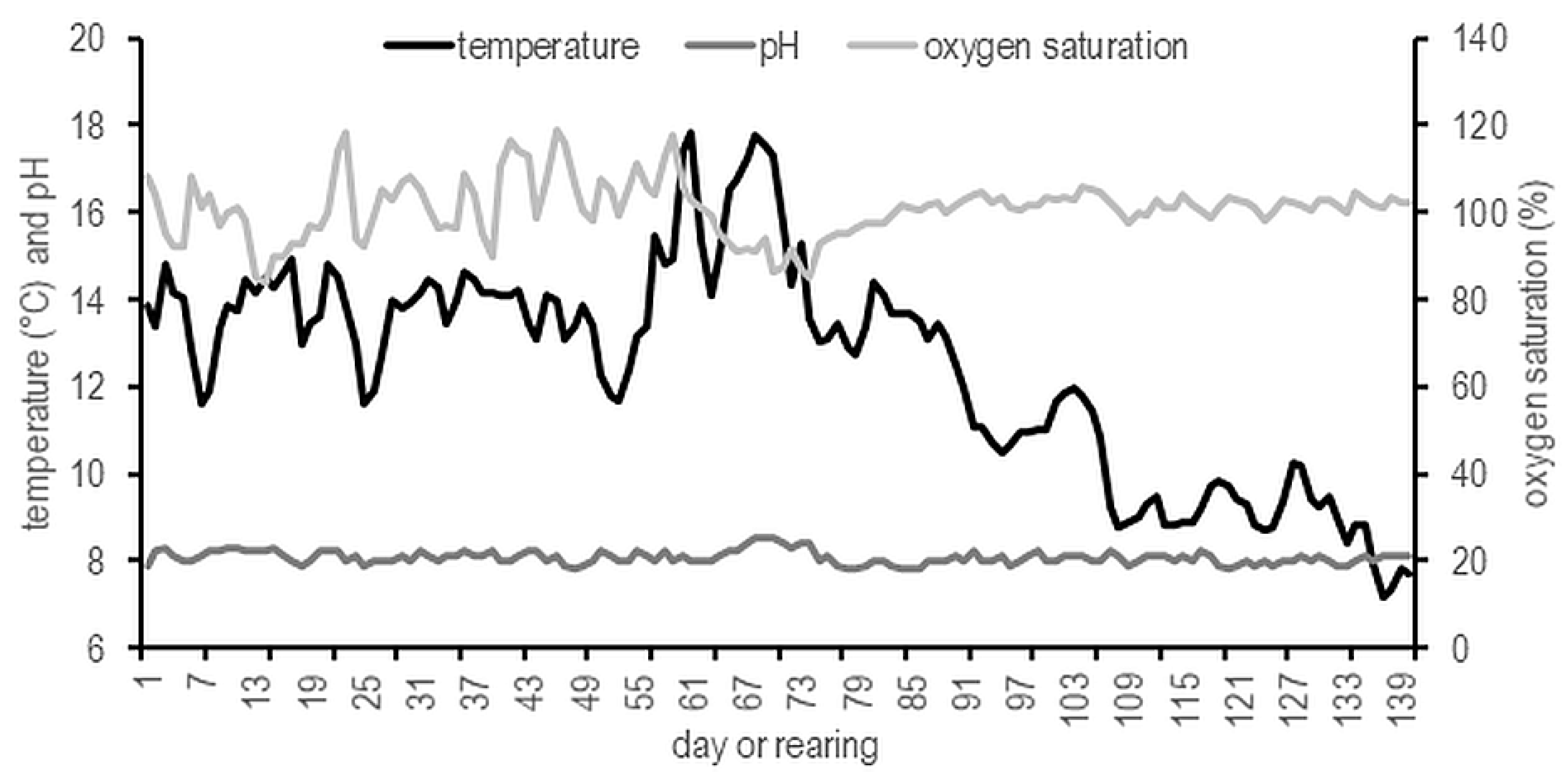
2.2. Data Collection and Sample Analysis
2.3. Statistical Analysis
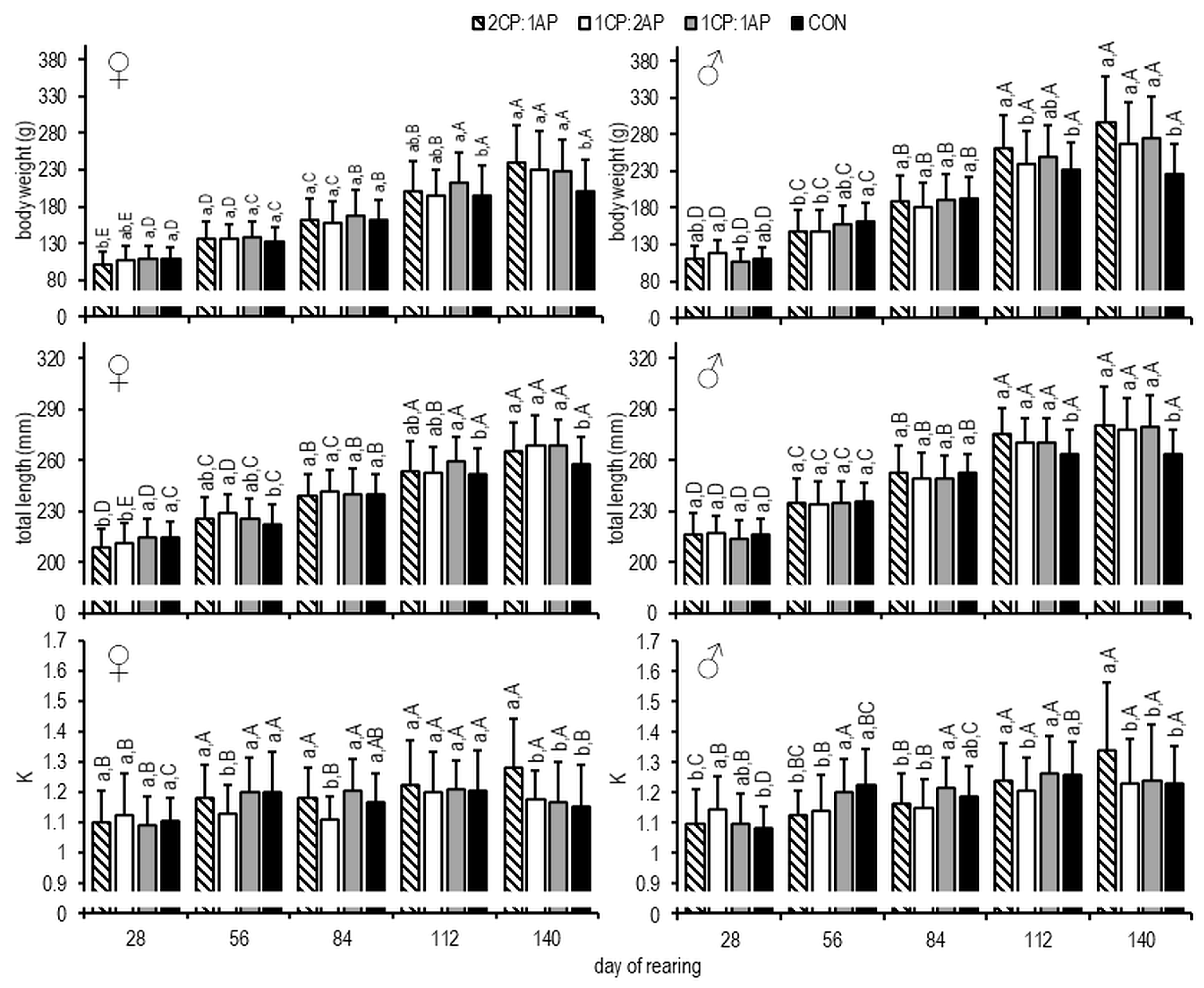
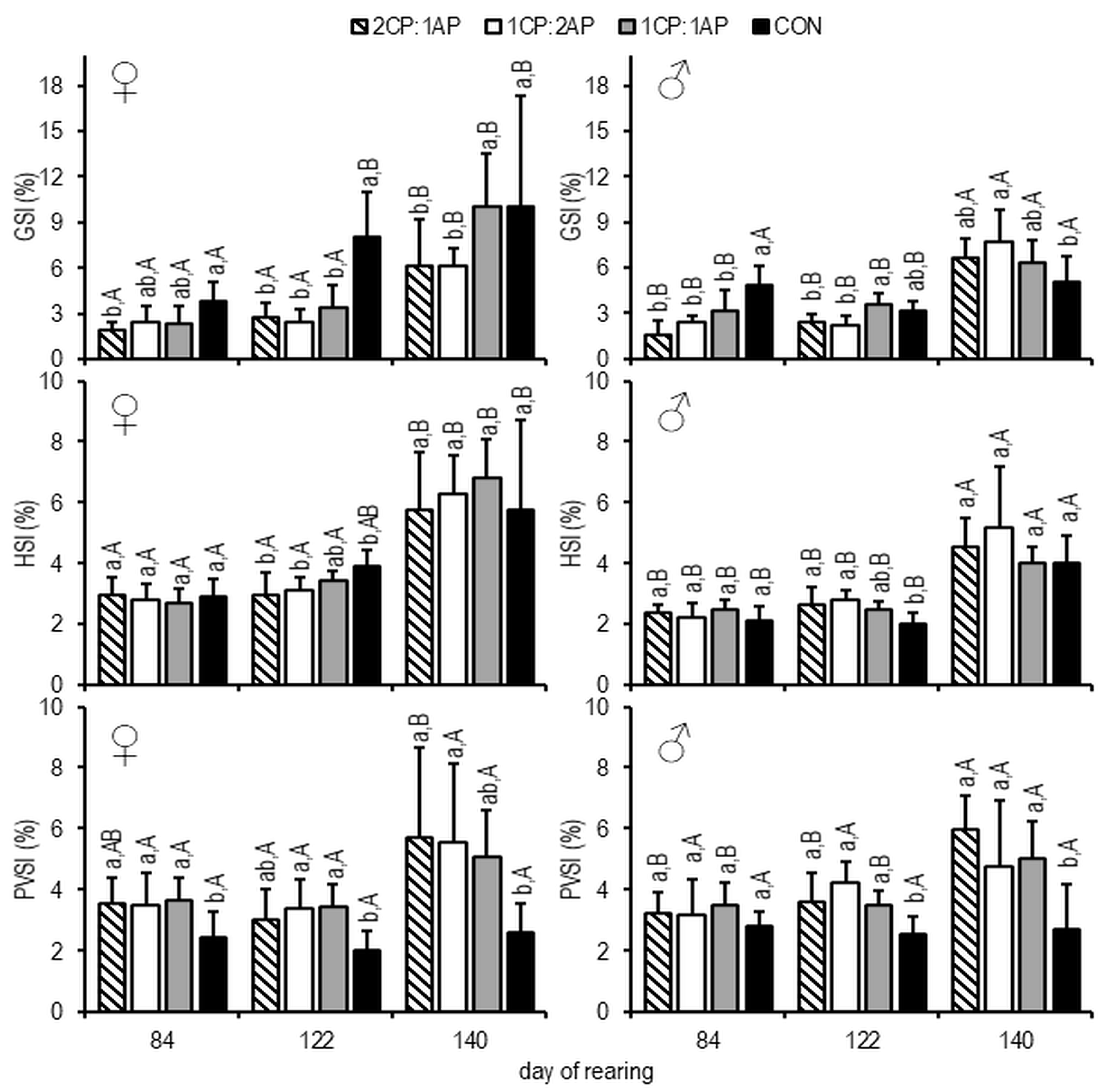
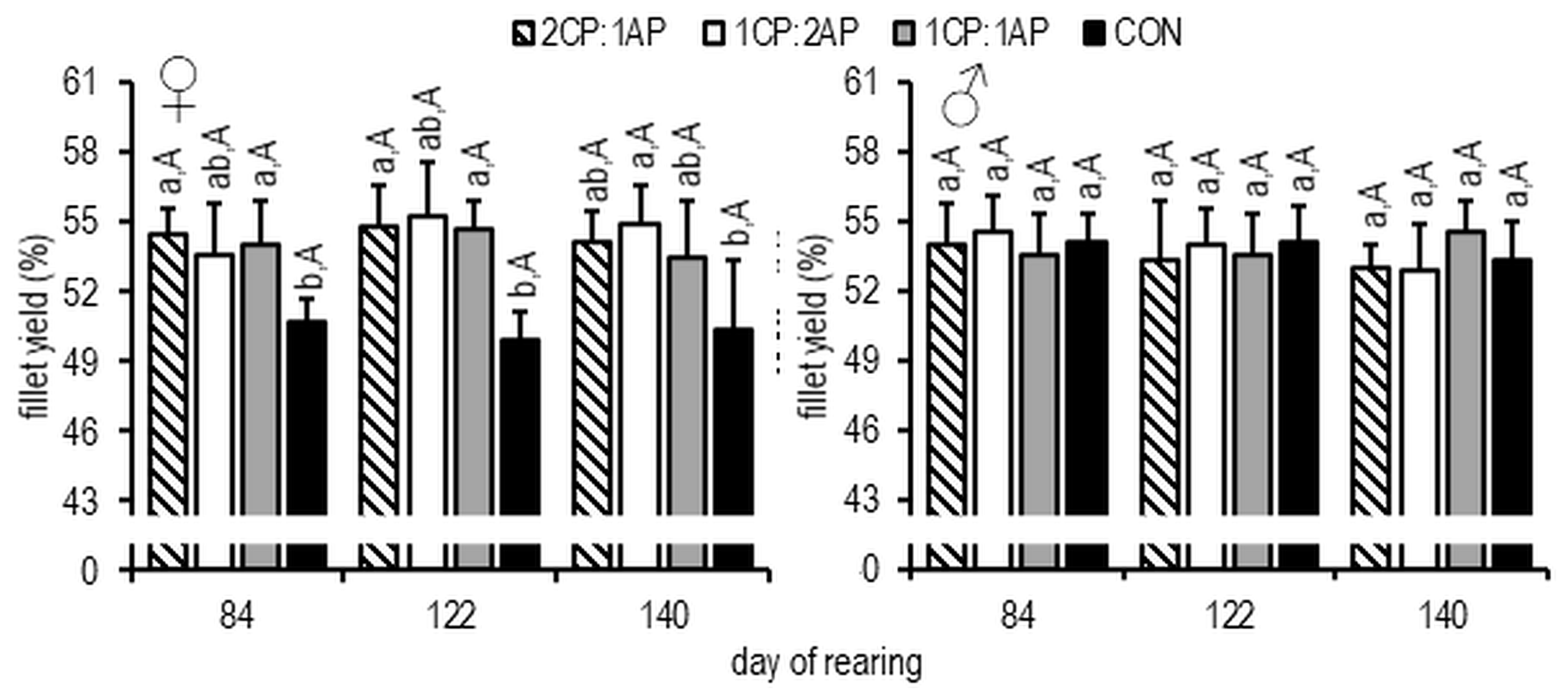
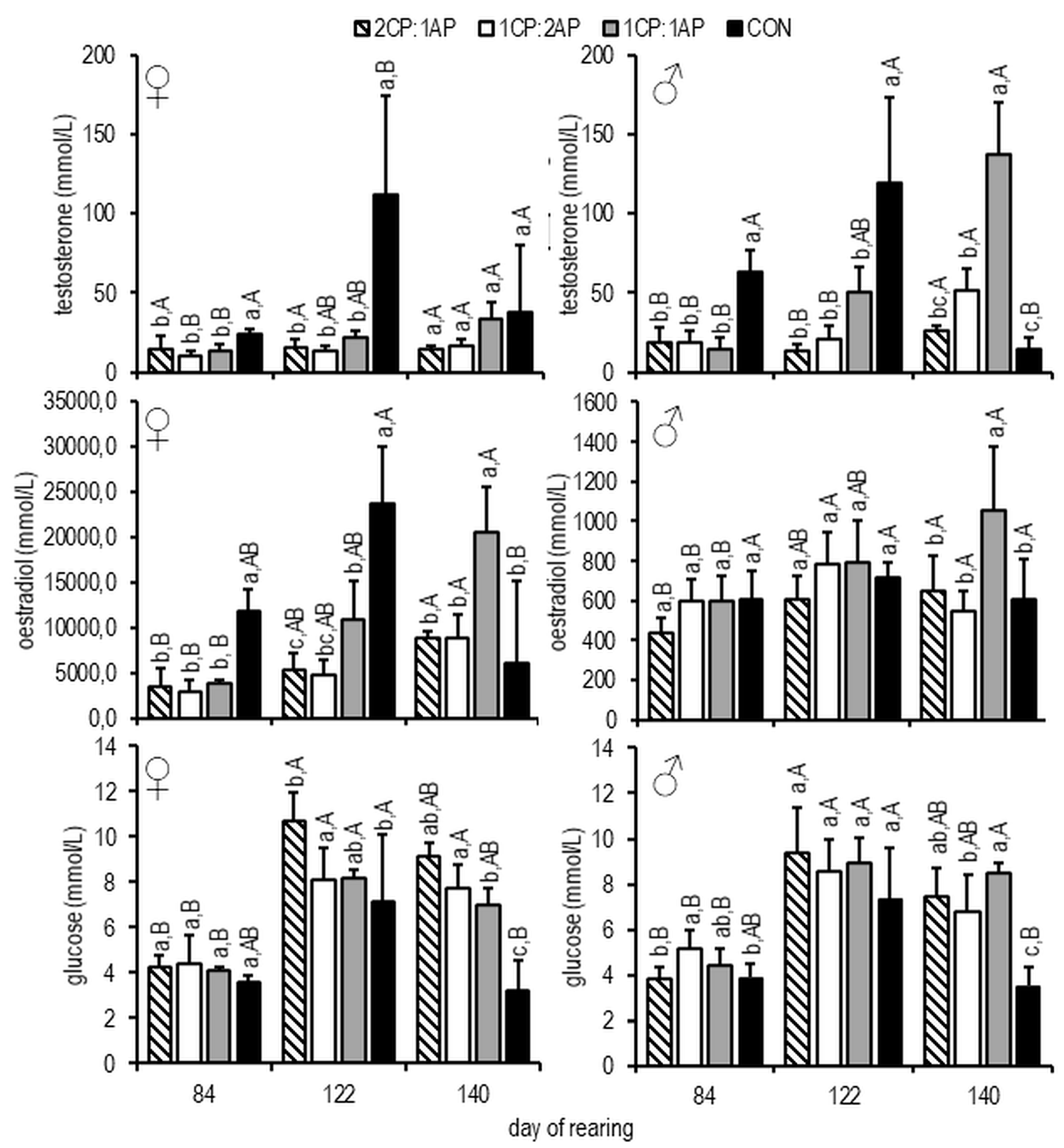
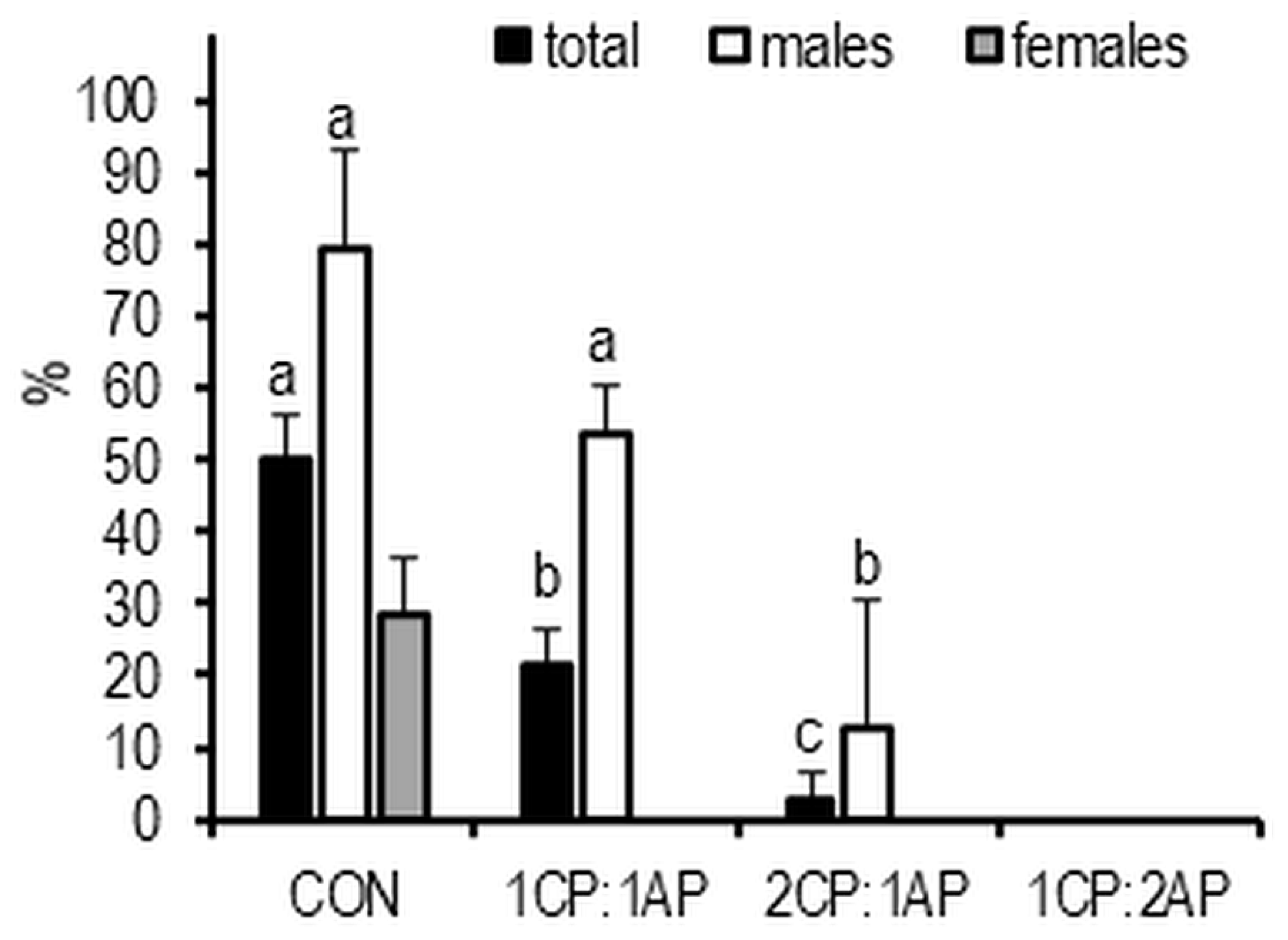
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Compliance with Ethical Standards
References
- Svinger, V.; Policar, T.; Steinbach, C.; Polakova, S.; Jankovych, A.; Kouril, J. Synchronization of ovulation in brook char (Salvelinus fontinalis, Mitchill 1814) using emulsified d-Arg6Pro9Net sGnRHa. Aquacult. Int. 2013, 21, 783–799. [Google Scholar] [CrossRef]
- Zajic, T.; Mraz, J.; Samples, S.; Pickova, J. Finishing feeding strategy as an instrument for modification of fatty acid composition of brook trout (Salvelinus fontinalis). Aquacult Int. 2016, 24, 1641–1656. [Google Scholar] [CrossRef]
- Lundova, K.; Matousek, J.; Prokesova, M.; Vanina, T.; Sebesta, R.; Urban, J.; Stejskal, V. The effects of a prolonged photoperiod and light sources on growth, sexual maturation, fin condition, and vulnerability to fungal disease in brook trout Salvelinus fontinalis. Aquacult. Res. 2019, 50, 256–267. [Google Scholar] [CrossRef]
- Taranger, G.L.; Carrillo, M.; Schulz, R.W.; Fontaine, P.; Zanuy, S.; Felip, A.; Weltzien, F.A.; Dufour, S.; Karlsen, O.; Norberg, B.; et al. Control of puberty in farmed fish. Gen. Comp. Endocrinol. 2010, 165, 483–515. [Google Scholar] [CrossRef]
- Fatima, S.; Adams, M.; Wilkinsos, R. Seasonal variation in the profile of sex steroids and histological testicular development of brook trout (Salvelinus fontinalis Mitchill) during the annual reproductive cycle in Tasmania. Aust. J. Zool. 2017, 65, 313–318. [Google Scholar] [CrossRef]
- Holcombe, G.W.; Pasha, M.S.; Jensen, K.M.; Tietge, J.F.; Ankley, G.T. Effects of photoperiod manipulation on brook trout reproductive development, fecundity, and circulating sex steroid concentrations. N. Am. J. Aquac. 2000, 62, 1–11. [Google Scholar] [CrossRef]
- Behnke, R.J. Trout and Salmon of North America; The Free Press, Simon and Achuster, Inc.: New York, NY, USA, 2002; p. 363. [Google Scholar]
- Hokanson, K.E.F.; McCormick, J.H.; Jones, B.R.; Tucker, J.H. Thermal requirements for maturation, spawning and embryo survival of the Brook Trout, Salvelinus fontinalis. Can. J. Fish Aquat. Sci. 1973, 30, 975–984. [Google Scholar] [CrossRef]
- Smith, D.A.; Ridgway, M.S. Temperature selection in Brook Charr: Lab experiments, field studies, and matching the Fry curve. Hydrobiologia 2019, 840, 143–156. [Google Scholar] [CrossRef]
- Stien, L.H.; Bracke, M.B.M.; Folkedal, O.; Nilsson, J.; Oppedal, F.; Torgersen, T.; Kittilsen, S.; Midtlyng, P.J.; Vindas, M.A.; Overli, O.; et al. Salmon welfare index model (SWIM 1.0): A semantic model for overall welfare assessment of caged Atlantic salmon: Review of the selected welfare indicators and model presentation. Rev. Aquacult. 2013, 5, 33–57. [Google Scholar] [CrossRef]
- Lundova, K.; Matousek, J.; Prokesova, M.; Sebesta, R.; Policar, T.; Stejskal, V. The effect of timing of extended photoperiod on growth and maturity of brook trout (Salvelinus fontinalis). Aquacult. Res. 2019, 50, 1697–1704. [Google Scholar] [CrossRef]
- Harris, J.; Bird, D.J. Modulation of the fish immune system by hormones. Vet. Immunol. Immunopathol. 2000, 77, 163–176. [Google Scholar] [CrossRef]
- Linhartova, Z.; Lunda, R.; Masilko, J.; Dvorak, P.; Lundova, K.; Stejskal, V.; Matousek, J.; Mraz, J. Impact of photostimulation for delayed maturity on flesh quality of brook trout (Salvelinus fontinalis) stored under refrigerated conditions. Aquacult. Res. 2018, 49, 3817–3829. [Google Scholar] [CrossRef]
- Iversen, M.; Myhr, A.I.; Wargelius, A. Approaches for delaying sexual maturation in salmon and their possible ecological and ethical implications. J. Appl. Aquacult. 2016, 28, 330–369. [Google Scholar] [CrossRef]
- Jonsson, B.; Finstad, A.G.; Jonsson, N. Winter temperature and food quality affect age at maturity: An experimental test with Atlantic salmon (Salmo salar). Can. J. Fish Aquat. Sci. 2012, 69, 1817–1826. [Google Scholar] [CrossRef]
- Thorpe, J.; Mangel, M.; Metcalfe, N.; Huntingford, F. Modelling the proximate basis of salmonid life-history variation, with application to Atlantic salmon, Salmo salar L. Evol. Ecol. 1998, 12, 581–599. [Google Scholar] [CrossRef]
- Pankhurst, N.W.; Munday, P.L. Effects of climate change on fish reproduction and early life history stages. Mar. Freshwater Res. 2011, 62, 1015–1026. [Google Scholar] [CrossRef]
- Bromage, N.; Randall, C.; Duston, J.; Thrush, M.; Jones, J. Environmental control of reproduction in salmonids. In Recent Advances in Aquaculture 4; Muir, J.F., Roberts, R.J., Eds.; Blackwell Scientific Publications: Oxford, UK, 1993; pp. 55–65. [Google Scholar]
- Schulz, R.W.; Andersson, E.; Taranger, G.L. Photoperiod manipulation can stimulate or inhibit pubertal testis maturation in Atlantic salmon (Salmo salar). Anim. Reprod. 2006, 3, 121–126. [Google Scholar]
- Fang, Y. Effects of Salinity and Photoperiod on Growth, Aerobic Scope, and Hypoxia Tolerance of Atlantic and Coho Salmon in Recirculating Aquaculture Systems; University of British Columbia: Vancouver, BC, Canada, 2018; p. 102. [Google Scholar] [CrossRef]
- Choi, S.; Lee, C.H.; Park, W.; Kim, D.-J.; Sohn, Y.C. Effects of shortened photoperiod on gonadotropin-releasing hormone, gonadotropin, and vitellogenin gene expression associated with ovarian maturation in rainbow trout. Zool. Sci. 2010, 27, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Sacobie, C.F.D.; Burke, H.A.; Lall, S.P.; Berfley, T.J. The effect of dietary energy level on growth and nutrient utilization by juvenile diploid and triploid brook char, Salvelinus fontinalis. Aquacult. Nutr. 2016, 22, 1091–1100. [Google Scholar] [CrossRef]
- Brown, E.E.; Baumann, H.; Conover, D.O. Temperature and photoperiod effects on sex determination in a fish. J. Exp. Mar. Bio. Ecol. 2014, 461, 39–43. [Google Scholar] [CrossRef]
- Türker, A.; Yıldırım, Ö. Interrelationship of Photoperiod with Growth Performance and Feeding of Seawater Farmed Rainbow Trout, (Oncorhynchus mykiss). Turk. J. Fish. Aquat. Sci. 2011, 11, 393–397. [Google Scholar] [CrossRef]
- Önder, M.Y.; Basçınar, N.; Khan, U.; Sonay, F.D. Effect of photoperiod on growth and efficiency of yolk-sac utilization in alevins of Brook Trout (Salvelinus fontinalis). Pak. J. Zool. 2016, 48, 533–537. [Google Scholar]
- Liu, Q.; Duston, J. Efficacy of 24 h light to reduce maturation in Arctic charr (Salvelinus alpinus) is dependent on both the start date and duration. Aquaculture 2018, 484, 44–50. [Google Scholar] [CrossRef]
- Andersson, E.; Schulz, R.W.; Male, R.; Bogerd, J.; Patiña, D.; Benedet, S.; Norberg, B.; Taranger, G.L. Pituitary gonadotropin and ovarian gonadotropin receptor transcript levels: Seasonal and photoperiod-induced changes in the reproductive physiology of female Atlantic salmon (Salmo salar). Gen. Comp. Endocrinol. 2013, 191, 247–258. [Google Scholar] [CrossRef]
- Taranger, G.L.; Haux, C.; Stefansson, S.O.; Bjornsson, B.T.; Walther, B.T.; Hansen, T. Abrupt changes in photoperiod affect age at maturity, timing of ovulation and plasma testosterone and oestradiol-17β profiles in Atlantic salmon, Salmo salar. Aquaculture 1998, 162, 85–98. [Google Scholar] [CrossRef]
- Takashima, F.; Yamada, Y. Control of maturation in masu salmon by manipulation of photoperiod. Aquaculture 1984, 43, 243–257. [Google Scholar] [CrossRef]
- Scott, A.P. Salmonids. In Reproductive Seasonality in Teleosts: Environmental Influences; Munro, A.G., Scott, A.P., Lam, T.J., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 33–53. [Google Scholar]
- Dalley, R. The survival and development of the shrimp Crangon crangon (L.), reared in the laboratory under non-circadian light-dark cycles. J. Exp. Mar. Biol. Ecol. 1980, 47, 101–112. [Google Scholar] [CrossRef]
- Von Saint Paul, U.; Aschoff, J. Longevity among blowflies Phormia terraenovae R.D. kept in non-24 hour light/dark cycles. J. Comp. Physiol. 1978, 127, 191–195. [Google Scholar] [CrossRef]
- Saunders, D.S. Circadian control of larval growth rate in Sarcophaga argyrostoma. Proc. Natl. Acad. Sci. USA 1972, 69, 2738–2740. [Google Scholar] [CrossRef] [PubMed]
- Dalley, R. Effects of non-circadian light cycles on the survival and development of Palaemon elegans Rathke reared in the laboratory. In Cyclic Phenomena in Marine Plants and Animals; Naylor, E., Hartnoll, R.G., Eds.; Pergamon Press: London, UK, 1979; pp. 157–163. [Google Scholar] [CrossRef]
- Qiu, D.; Xu, S.; Song, C.; Chi, L.; Li, X.; Sun, G.; Liu, B.; Liu, Y. Effects of spectral composition, photoperiod and light intensity on the gonadal development of Atlantic salmon Salmo salar in recirculating aquaculture systems (RAS). Chin. J. Ocean Limnol. 2015, 33, 45–56. [Google Scholar] [CrossRef]
- Pavlidis, M.; Dimitriou, D.; Dessypris, A. Testosterone and 17-b-estradiol plasma fluctuations throughout spawning period in male and female rainbow trout, Oncorhynchus mykiss (Walbaum), kept under several photoperiod regimes. Ann. Zool. Fennici 1994, 31, 319–327. Available online: https://www.jstor.org/stable/23735536 (accessed on 27 March 2020).
- Frantzen, M.; Arnesen, A.M.; Damsgǻrd, B.; Tveiten, H.; Johnsen, H.K. Effects of photoperiod on sex steroids and gonad maturation in Arctic charr. Aquaculture 2004, 240, 561–574. [Google Scholar] [CrossRef]
- Mayer, I.; Lundqvist, H.; Bergulund, I.; Schmitz, M.; Schulz, R.; Borg, B. Seasonal endocrine changes in Baltic salmon, Salmo salar, immature parr and mature male parr. I. Plasma levels of five androgens, 17a-hydroxy-20-b-dihydroprogesterone, and 17-b-estradiol. Can. J. Zool. 1990, 68, 1360–1365. [Google Scholar] [CrossRef]
- Schulz, R.W.; Miura, T. Spermatogenesis and its endocrine regulation. Fish Physiol. Biochem. 2002, 26, 43–56. [Google Scholar] [CrossRef]
- Taylor, J.F.; Migaud, H.; Porter, M.J.R.; Bromage, N.R. Photoperiod influences growth rate and insulin-like growth factor-I (IGF-I) levels in juvenile rainbow trout, Oncorhynchus mykiss. Gen. Comp. Endocrinol. 2005, 142, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.J.; Fjelldal, P.G.; Folkedal, O.; Vagseth, T.; Oppedal, F. Effects of light source and intensity on sexual maturation, growth and swimming behaviour of Atlantic salmon in sea cages. Aquac. Environ. Interact. 2017, 9, 193–204. [Google Scholar] [CrossRef]
- Cleveland, B.M.; Weber, G.M. Effects of steroid treatment on growth, nutrient partitioning, and expression of genes related to growth and nutrient matabolism in adult triploid rainbow trout (Oncorhynchus mykiss). Domest. Anim. Endocrinol. 2016, 56, 1–13. [Google Scholar] [CrossRef]
- Cleveland, B.M.; Weber, G.M. Effects of sex steroids on expression of genes regulating growth-related mechanisms in rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol. 2015, 216, 103–115. [Google Scholar] [CrossRef]
- Cleveland, B.M.; Manor, M.L. Effects of phytoestrogens on growthrelated and lipogenic genes in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. C 2015, 170, 28–37. [Google Scholar] [CrossRef]
- Cleveland, B.M.; Weber, G.M. Effects of sex steroids on indices of protein turnover in rainbow trout (Oncorhynchus mykiss) white muscle. Gen. Comp. Endocrinol. 2011, 174, 132–142. [Google Scholar] [CrossRef]
- Noori, A.; Mojazi Amiri, B.; Mirvaghefi, A.; Rafiee, G.; Kalvani Neitali, B. Enhanced growth and retarded gonadal development of farmed rainbow trout, Oncorhynchus mykiss (Walbaum) following a long-day photoperiod. Aquac. Res. 2015, 46, 2398–2406. [Google Scholar] [CrossRef]
- Randall, C.F.; North, B.; Futter, W.; Porter, M.; Bromage, N. Photoperiod effects on reproduction and growth in rainbow trout. Trout News 2001, 32, 12–16. [Google Scholar]
- Taylor, J.F.; North, B.P.; Porter, M.J.R.; Bromage, N.R.; Migaud, H. Photoperiod can be used to enhance growth and improve feeding efficiency in farmed rainbow trout, Oncorhynchus mykiss. Aquaculture 2006, 256, 216–234. [Google Scholar] [CrossRef]
- Shearer, K.; Parkins, P.; Gadberry, B.; Beckman, B.; Swanson, P. Effects of growth rate/body size and a low lipid diet on the incidence of early sexual maturation in juvenile male spring Chinook salmon (Oncorhynchus tshawytscha). Aquaculture 2006, 252, 545–556. [Google Scholar] [CrossRef]
- Aubin-Horth, N.; Bourque, J.F.; Daigle, G.; Hedger, R.; Dodson, J.J. Longitudinal gradients in threshold sizes for alternative male life history tactics in a population of Atlantic salmon (Salmo salar). Can. J. Fish Aquat. Sci. 2006, 63, 2067–2075. [Google Scholar] [CrossRef]
- Kumar, V. Photoperiodism in higher vertebrates: An adaptive strategy in temporal environment. Indian J. Exp. Biol. 1997, 35, 427–437. [Google Scholar]
- Dawson, A.; King, V.M.; Bentley, G.E.; Ball, G.F. Photoperiodic control of seasonality in birds. J. Biol. Rhythms 2001, 16, 365–380. [Google Scholar] [CrossRef]
- Budki, P.; Rani, S.; Kumar, V. Persistence of circannual rhythms under constant periodic and aperiodic light conditions: Sex differences and relationship with the external environment. J. Exp. Biol. 2012, 215, 3774–3785. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lundova, K.; Matousek, J.; Stejskal, V. The Effect of Non-Circadian Photoperiod on Growth and Puberty Onset of Brook Trout Salvelinus fontinalis Mitchill. Animals 2021, 11, 692. https://doi.org/10.3390/ani11030692
Lundova K, Matousek J, Stejskal V. The Effect of Non-Circadian Photoperiod on Growth and Puberty Onset of Brook Trout Salvelinus fontinalis Mitchill. Animals. 2021; 11(3):692. https://doi.org/10.3390/ani11030692
Chicago/Turabian StyleLundova, Katsiaryna, Jan Matousek, and Vlastimil Stejskal. 2021. "The Effect of Non-Circadian Photoperiod on Growth and Puberty Onset of Brook Trout Salvelinus fontinalis Mitchill" Animals 11, no. 3: 692. https://doi.org/10.3390/ani11030692
APA StyleLundova, K., Matousek, J., & Stejskal, V. (2021). The Effect of Non-Circadian Photoperiod on Growth and Puberty Onset of Brook Trout Salvelinus fontinalis Mitchill. Animals, 11(3), 692. https://doi.org/10.3390/ani11030692