
Effect of Fasting on the Spexin System in Broiler Chickens

 ,
,  , , , and
, , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds and Diets
2.2. Sampling
2.3. Chemical Analyses
2.4. Biochemical Profile
2.5. Hormonal Profile
2.6. Real-Time PCR
2.7. Statistical Analysis
3. Results
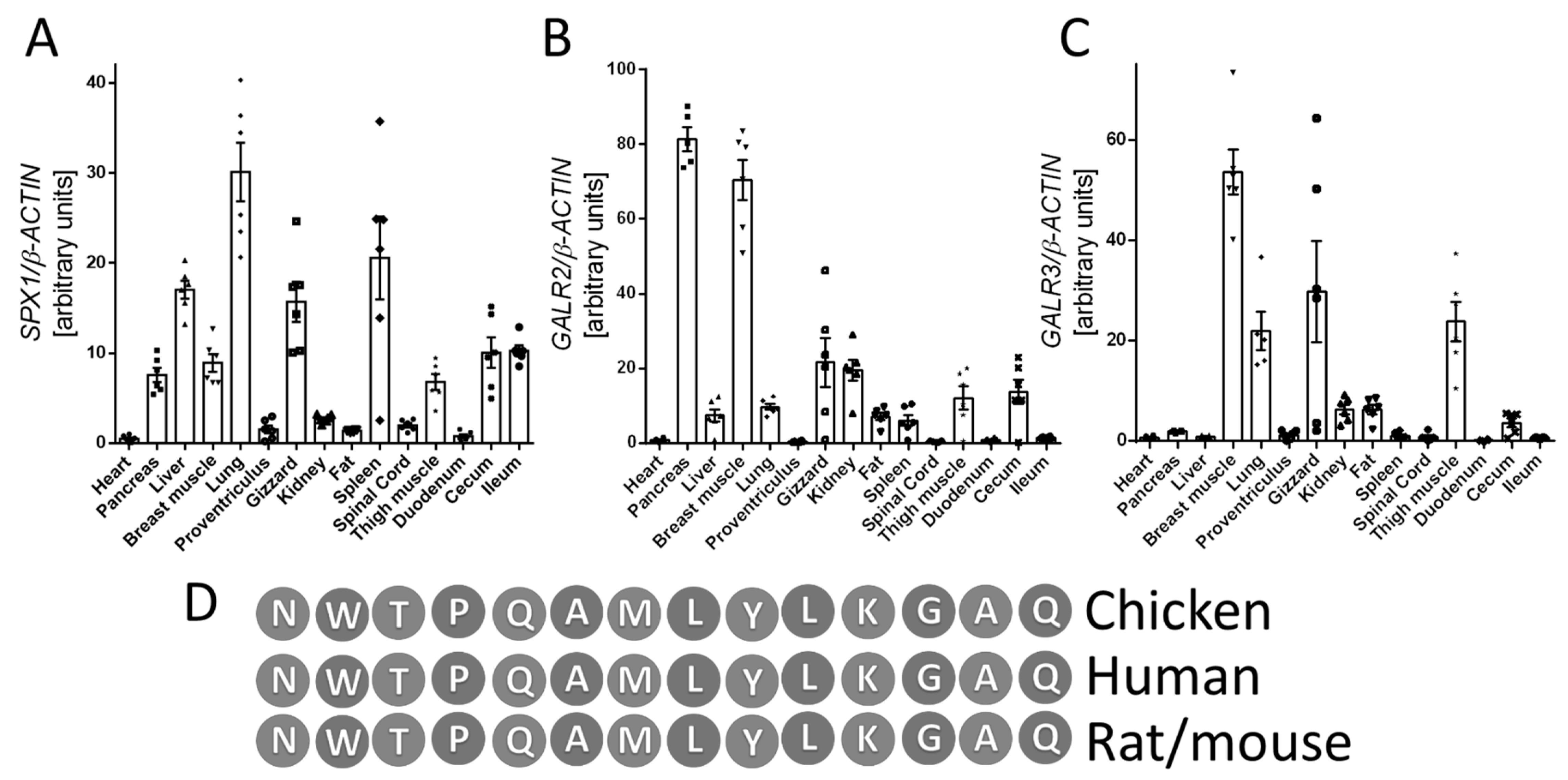
3.1. Tissue Expression of SPX1, GALR2, and GALR3
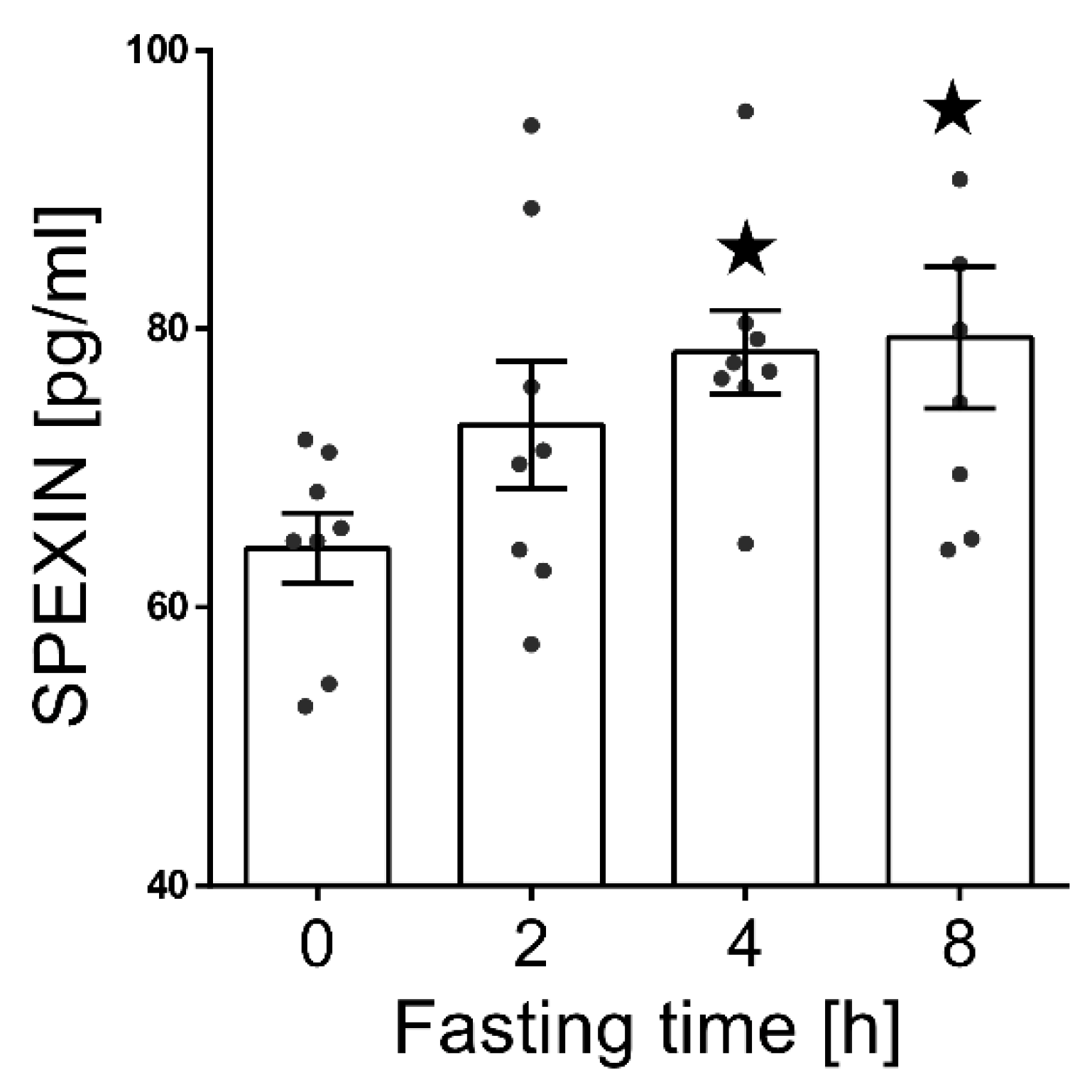
3.2. Effect of Fasting on the SPX Concentration in Blood Serum.
3.3. Effect of Fasting on Basic Metabolic and Hormonal Profile.
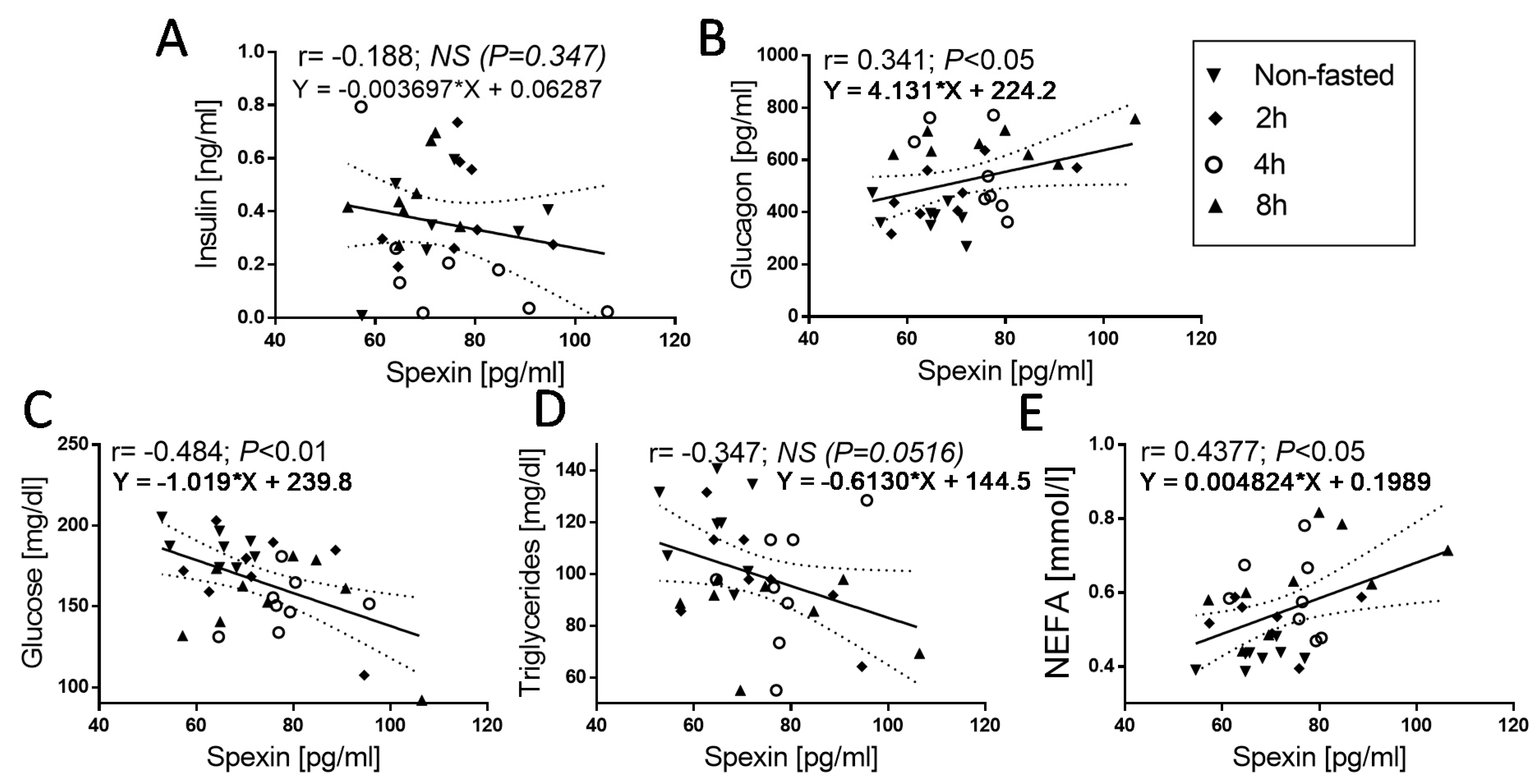
3.4. Correlation of SPX Concentration with Others Metabolic Parameters
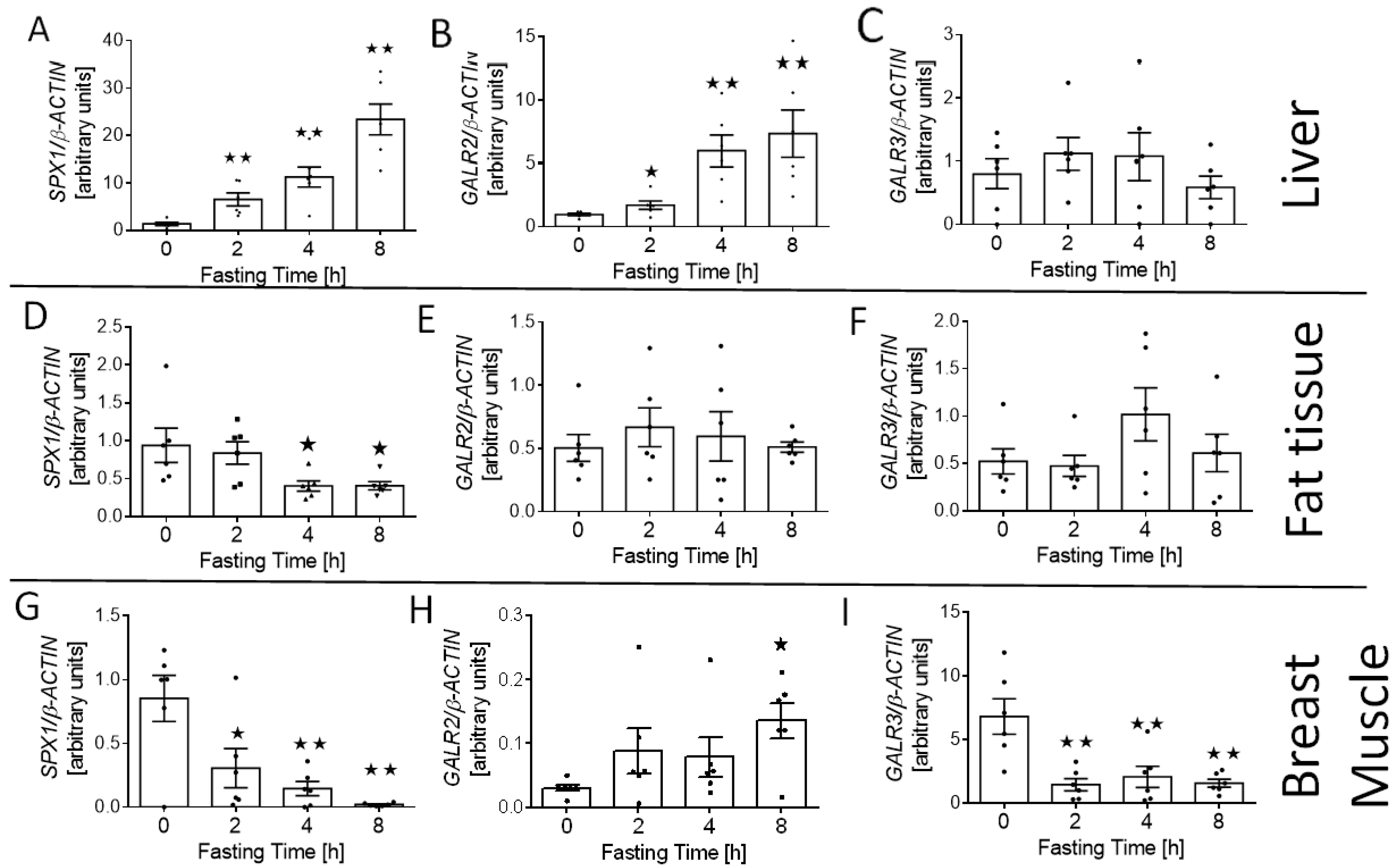
3.5. Effect of Fasting on SPX1, GALR2, and GALR3 mRNA Expression in Breast Muscle, Liver, and Fat Tissue
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Mirabeau, O.; Perlas, E.; Severini, C.; Audero, E.; Gascuel, O.; Possenti, R.; Birney, E.; Rosenthal, N.; Gross, C. Identification of novel peptide hormones in the human proteome by hidden Markov model screening. Genome Res. 2007, 17, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.H.; Lee, M.Y.M.; Soga, T.; Parhar, I. Evolution of structural and functional diversity of spexin in mammalian and non-mammalian vertebrate species. Front. Endocrinol. (Lausanne) 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Kim, D.K.; Yun, S.; Son, G.H.; Hwang, J.I.; Park, C.R.; Kim, J., II; Kim, K.; Vaudry, H.; Seong, J.Y. Coevolution of the spexin/galanin/kisspeptin family: Spexin activates galanin receptor type II and III. Endocrinology 2014, 155, 1864–1873. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejski, P.A.; Pruszynska-Oszmalek, E.; Micker, M.; Skrzypski, M.; Wojciechowicz, T.; Szwarckopf, P.; Skieresz-Szewczyk, K.; Nowak, K.W.; Strowski, M.Z. Spexin: A novel regulator of adipogenesis and fat tissue metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 1228–1236. [Google Scholar] [CrossRef]
- Jasmine, G.; Walewski, J.; Anglade, D.; Berk, P. Regulation of Hepatocellular Fatty Acid Uptake in Mouse Models of Fatty Liver Disease with and without Functional Leptin Signaling: Roles of NfKB and SREBP-1C and the Effects of Spexin. Semin. Liver Dis. 2016, 36, 360–372. [Google Scholar] [CrossRef]
- Walewski, J.L.; Ge, F.; Lobdell, H.; Levin, N.; Schwartz, G.J.; Vasselli, J.R.; Pomp, A.; Dakin, G.; Berk, P.D. Spexin is a novel human peptide that reduces adipocyte uptake of long chain fatty acids and causes weight loss in rodents with diet-induced obesity. Obesity 2014, 22, 1643–1652. [Google Scholar] [CrossRef]
- Wu, H.; Lin, F.; Chen, H.; Liu, J.; Gao, Y.; Zhang, X.; Hao, J.; Chen, D.; Yuan, D.; Wang, T.; et al. Ya-fish (Schizothorax prenanti) spexin: Identification, tissue distribution and mRNA expression responses to periprandial and fasting. Fish. Physiol. Biochem. 2015. [Google Scholar] [CrossRef]
- Liu, Y.; Li, S.; Qi, X.; Zhou, W.; Liu, X.; Lin, H.; Zhang, Y.; Cheng, C.H.K. A novel neuropeptide in suppressing luteinizing hormone release in goldfish, Carassius auratus. Mol. Cell. Endocrinol. 2013, 374, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zhang, M.; Huang, T.; Yang, L.; Fu, H.; Zhao, L.; Zhong, L.L.; Mu, H.; Shi, X.; Leung, C.F.; et al. Spexin Enhances Bowel Movement through Activating L-type Voltage-dependent Calcium Channel via Galanin Receptor 2 in Mice. Sci. Rep. 2015, 5, 12095. [Google Scholar] [CrossRef] [PubMed]
- Sassek, M.; Kolodziejski, P.A.; Szczepankiewicz, D.; Pruszynska-Oszmalek, E. Spexin in the physiology of pancreatic islets—mutual interactions with insulin. Endocrine 2019, 63, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Sassek, M.; Kolodziejski, P.A.; Strowski, M.Z.; Nogowski, L.; Nowak, K.W.; Mackowiak, P. Spexin Modulates Functions of Rat Endocrine Pancreatic Cells. Pancreas 2018, 47, 904–909. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.; He, M.; Bai, J.; Wong, M.K.H.; Ko, W.K.W.; Wong, A.O.L. Dual role of insulin in Spexin regulation: Functional link between food intake and Spexin expression in a fish model. Endocrinology 2017, 158, 560–577. [Google Scholar] [CrossRef]
- Smulikowska, S. Recommended Allowances and Nutritive Value of Feedstuffs—Poultry Feeding Standards, 5th ed.; Kielanowski Institute of Animal Physiology and Nutrition, PAS: Jablonna, Poland, 2005. (In Polish) [Google Scholar]
- Aviagen Ross 308 Broiler: Nutrition Specifications 2019. Aviagen 2020, 1–10. Available online: https://tmea.aviagen.com/assets/Tech_Center/Ross_Broiler/RossBroilerNutritionSpecs2019-EN.pdf (accessed on 20 October 2020).
- Horwitz, W. Official Methods of Analysis of AOAC International; agricultural chemicals, contaminants, drugs; AOAC International: Gaithersburg, MD, USA, 2005; Volume I, ISBN 0935584773. [Google Scholar]
- Pruszynska-Oszmalek, E.; Kolodziejski, P.A.; Stadnicka, K.; Sassek, M.; Chalupka, D.; Kuston, B.; Nogowski, L.; Mackowiak, P.; Maiorano, G.; Jankowski, J.; et al. In ovo injection of prebiotics and synbiotics affects the digestive potency of the pancreas in growing chickens. Poult. Sci. 2015, 94, 1909–1916. [Google Scholar] [CrossRef]
- Kolodziejski, P.A.; Sassek, M.; Chalupka, D.; Leciejewska, N.; Nogowski, L.; Mackowiak, P.; Jozefiak, D.; Stadnicka, K.; Siwek, M.; Bednarczyk, M.; et al. GLP1 and GIP are involved in the action of synbiotics in broiler chickens. J. Anim. Sci. Biotechnol. 2018, 9. [Google Scholar] [CrossRef]
- Porzionato, A.; Rucinski, M.; Macchi, V.; Stecco, C.; Malendowicz, L.K.; De Caro, R. Spexin expression in normal rat tissues. J. Histochem. Cytochem. 2010, 58, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.C.W.; Jacobs, T.; Wang, Y.; Leung, F.C. Identification and characterization of the chicken galanin receptor GalR2 and a novel GalR2-like receptor (GalR2-L). Gen. Comp. Endocrinol. 2012, 179, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Boschiero, C.; Moreira, G.C.M.; Gheyas, A.A.; Godoy, T.F.; Gasparin, G.; Mariani, P.D.S.C.; Paduan, M.; Cesar, A.S.M.; Ledur, M.C.; Coutinho, L.L. Genome-wide characterization of genetic variants and putative regions under selection in meat and egg-type chicken lines. BMC Genom. 2018, 19, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gambaro, S.E.; Zubiría, M.G.; Giordano, A.P.; Portales, A.E.; Alzamendi, A.; Rumbo, M.; Giovambattista, A. Spexin improves adipose tissue inflammation and macrophage recruitment in obese mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158700. [Google Scholar] [CrossRef]
- Mills, E.G.; Izzi-Engbeaya, C.; Abbara, A.; Comninos, A.N.; Dhillo, W.S. Functions of galanin, spexin and kisspeptin in metabolism, mood and behaviour. Nat. Rev. Endocrinol. 2020. [Google Scholar] [CrossRef]
- Lv, S.Y.; Zhou, Y.C.; Zhang, X.M.; Chen, W.D.; Wang, Y.D. Emerging roles of NPQ/Spexin in physiology and pathology. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Pruszynska-Oszmalek, E.; Sassek, M.; Szczepankiewicz, D.; Nowak, K.W.; Kolodziejski, P.A. Short-term administration of spexin in rats reduces obesity by affecting lipolysis and lipogenesis: An in vivo and in vitro study. Gen. Comp. Endocrinol. 2020, 299, 113615. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.J.; Persia, M.E.; Feierstein, E.; Kingham, B.; Saylor, W.W. Comparison of a modern broiler line and a heritage line unselected since the 1950s. Poult. Sci. 2009, 88, 2610–2619. [Google Scholar] [CrossRef]
- Honda, K.; Takagi, S.; Kurachi, K.; Sugimoto, H.; Saneyasu, T.; Kamisoyama, H. Fasting and glucagon stimulate gene expression of pyruvate dehydrogenase kinase 4 in chickens. J. Poult. Sci. 2017, 54, 292–295. [Google Scholar] [CrossRef]
- Cogburn, L.A.; Trakooljul, N.; Wang, X.; Ellestad, L.E.; Porter, T.E. Transcriptome analyses of liver in newly-hatched chicks during the metabolic perturbation of fasting and re-feeding reveals THRSPA as the key lipogenic transcription factor. BMC Genom. 2020, 21, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Serr, J.; Suh, Y.; Lee, K. Regulation of adipose triglyceride lipase by fasting and refeeding in avian species. Poult. Sci. 2009, 88, 2585–2591. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/100 g, As-Fed Basis) | Starter | Grower |
|---|---|---|
| Maize | 51.81 | 54.7 |
| Soybean meal | 38.1 | 34.7 |
| Soybean oil | 5.58 | 6.57 |
| Monocalcium phosphate | 1.88 | 1.66 |
| Premix 1 | 1.00 | 1.00 |
| Limestone (<2 mm) | 0.38 | 0.29 |
| NaHCO3 | 0.33 | 0.30 |
| NaCl | 0.22 | 0.26 |
| HCl-Lysine | 0.31 | 0.19 |
| DL-Methionine | 0.22 | 0.14 |
| L-Threonine | 0.13 | 0.05 |
| L-Valine | 0.05 | 0.14 |
| Calculated Nutrient Composition (g/100 g Unless Otherwise Stated) | ||
| AMEN [MJ/kg] | 12.8 | 13.2 |
| Crude protein | 23.0 | 21.5 |
| Ca | 0.96 | 0.87 |
| Available P | 0.48 | 0.43 |
| Available Lys | 1.28 | 1.15 |
| Determined (g/100 g) | ||
| Crude protein | 23.4 | 21.1 |
| Crude fat | 6.36 | 7.72 |
| Ca | 0.88 | 0.80 |
| Total P | 0.61 | 0.59 |
| Primer | Sequence 5′ > 3′ | Product Size (bp) | Accession No. | Efficiency |
|---|---|---|---|---|
| SPX1 (Forward) | GTGCACAGGGACGTCGATT | 95 | KT235743.1 | 98.78% |
| SPX1 (Reverse) | AGGATTTGTGTTTTGGCTGCG | |||
| GALR3 (Forward) | GCTGGAATGCCTCCTCTGAC | 373 | NM_001130585.1 | 97.67% |
| GALR3 (Reverse) | CGGGACTTCAGTGGATAGCG | |||
| GALR2 (Forward) | ATCCCGAGTCGGTGCTCAT | 583 | NM_001128063 | 98.11% |
| GALR2 (Reverse) | CTCCAAAGGTAGCGAATGGT | |||
| β-ACTIN (Forward) | CCCAGACATCAGGGTGTGATG | 123 | NM_205518 | 102.86% |
| β-ACTIN (Reverse) | GTTGGTGACAATACCGTGTTCAAT |
| Variable | Fasting Time | |||
|---|---|---|---|---|
| 0 h | 2 h | 4 h | 8 h | |
| [mg/dL] | ||||
| Glucose | 186.9 ± 3.37 | 168.5 ± 9.25 | 150.6 ± 5.18 ** | 152.8 ± 9.41 ** |
| TG | 119.8 ± 5.58 | 97.96 ± 6.53 | 94.9 ± 7.39 * | 91.5 ± 6.23 * |
| Cholesterol | 125.6 ± 3.66 | 127.6 ± 5.83 | 131.2 ± 4.12 | 126.5 ± 2.59 |
| [mmol/L] | ||||
| NEFA | 0.438 ± 0.016 | 0.535 ± 0.03 | 0.575 ± 0.044 * | 0.631 ± 0.047 ** |
| [g/dL] | ||||
| Albumin | 1.473 ± 0.038 | 1.414 ± 0.05 | 1.462 ± 0.036 | 1.453 ± 0.018 |
| Total protein | 3.116 ± 0.110 | 3.062 ± 0.061 | 2.997 ± 0.067 | 3.018 ± 0.073 |
| [ng/mL] | ||||
| Insulin | 0.444 ± 0.073 | 0.349 ± 0.071 | 0.427 ± 0.071 | 0.122 ± 0.037 * |
| [pg/mL] | ||||
| Glucagon | 378.9 ± 19.99 | 474.6 ± 37.80 | 537.6 ± 52.53 * | 663.2 ± 20.97 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kołodziejski, P.A.; Pruszyńska-Oszmałek, E.; Hejdysz, M.; Sassek, M.; Leciejewska, N.; Ziarniak, K.; Bień, J.; Ślósarz, P.; Kubiś, M.; Kaczmarek, S. Effect of Fasting on the Spexin System in Broiler Chickens. Animals 2021, 11, 518. https://doi.org/10.3390/ani11020518
Kołodziejski PA, Pruszyńska-Oszmałek E, Hejdysz M, Sassek M, Leciejewska N, Ziarniak K, Bień J, Ślósarz P, Kubiś M, Kaczmarek S. Effect of Fasting on the Spexin System in Broiler Chickens. Animals. 2021; 11(2):518. https://doi.org/10.3390/ani11020518
Chicago/Turabian StyleKołodziejski, Paweł A., Ewa Pruszyńska-Oszmałek, Marcin Hejdysz, Maciej Sassek, Natalia Leciejewska, Kamil Ziarniak, Jakub Bień, Piotr Ślósarz, Marta Kubiś, and Sebastian Kaczmarek. 2021. "Effect of Fasting on the Spexin System in Broiler Chickens" Animals 11, no. 2: 518. https://doi.org/10.3390/ani11020518
APA StyleKołodziejski, P. A., Pruszyńska-Oszmałek, E., Hejdysz, M., Sassek, M., Leciejewska, N., Ziarniak, K., Bień, J., Ślósarz, P., Kubiś, M., & Kaczmarek, S. (2021). Effect of Fasting on the Spexin System in Broiler Chickens. Animals, 11(2), 518. https://doi.org/10.3390/ani11020518