
Effect of Fresh Citrus Pulp Supplementation on Animal Performance and Meat Quality of Feedlot Steers

 , ,
, ,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Dietary Treatments
2.2. Animal Determinations
2.3. Feed Determinations
2.4. Energy Calculation of the Diet
2.5. Carcass and Meat Quality Determinations
2.6. Micronutrient Analysis
2.7. Antioxidant Capacity Determinations
2.8. Statistical Analysis
3. Results
3.1. Animal Performance
3.2. Ruminal pH
3.3. Carcass and Meat Quality
3.4. Micronutrients and Meat Antioxidant Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kajikawa, H. Utilization of by-products from food processing as livestock feed in Japan. In Extension Bulletins of Food and Fertilizer Technology Center; ASPAC Food & Fertilizer Technology Center: Taipei, Taiwan, 1996; pp. 1–8. [Google Scholar]
- Kasapidou, E.; Sossidou, E.; Mitlianga, P. Fruit and Vegetable Co-Products as Functional Feed Ingredients in Farm Animal Nutrition for Improved Product Quality. Agriculture 2015, 5, 1020–1034. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, A.; Sarkar, S.; Lal, B.; Kumawat, P.; Sharma, S.; De, K. Utilization of fruit and vegetable waste as an alternative feed resource for sustainable and eco-friendly sheep farming. Waste Manag. 2021, 128, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Ominski, K.; McAllister, T.; Stanford, K.; Mengistu, G.; Kebebe, E.G.; Omonijo, F.; Cordeiro, M.; Legesse, G.; Wittenberg, K. Utilization of by-products and food waste in livestock production systems: A Canadian perspective. Anim. Front. 2021, 11, 55–63. [Google Scholar] [CrossRef]
- Wadhwa, M.W.M. Waste to worth: Fruit wastes and by-products as animal feed. CAB Rev. 2015, 10, 1–26. [Google Scholar] [CrossRef]
- Bampidis, V.; Robinson, P. Citrus by-products as ruminant feeds: A review. Anim. Feed. Sci. Technol. 2006, 128, 175–217. [Google Scholar] [CrossRef]
- Fernandez-Lopez, J.; Fernández-Ginés, J.; Aleson-Carbonell, L.; Sendra, E.; Sayas-Barberá, E.; Pérez-Alvarez, J.A. Application of functional citrus by-products to meat products. Trends Food Sci. Technol. 2004, 15, 176–185. [Google Scholar] [CrossRef]
- Peacock, F.M.; Kirk, W.G. Comparative Feeding Value of Dried Citrus Pulp, Corn Feed Meal and Ground Snapped Corn for Fattening Steers in Drylot; Bulletin 616; University of Florida Agricultural Experiment Station: Gainesville, FL, USA, 1959. [Google Scholar]
- Kirk, W.G.; Koger, M. Citrus Products in Cattle Finishing Rations: A Review of Research at the Range Cattle Station; Bulletin 739; University of Florida Agricultural Experiment Station: Gainesville, FL, USA, 1970. [Google Scholar]
- Salami, S.A.; Luciano, G.; O’Grady, M.N.; Biondi, L.; Newbold, C.J.; Kerry, J.P.; Priolo, A. Sustainability of feeding plant by-products: A review of the implications for ruminant meat production. Anim. Feed. Sci. Technol. 2019, 251, 37–55. [Google Scholar] [CrossRef]
- Gravador, R.S.; Jongberg, S.; Andersen, M.L.; Luciano, G.; Priolo, A.; Lund, M. Dietary citrus pulp improves protein stability in lamb meat stored under aerobic conditions. Meat Sci. 2014, 97, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Inserra, L.; Priolo, A.; Biondi, L.; Lanza, M.; Bognanno, M.; Gravador, R.; Luciano, G. Dietary citrus pulp reduces lipid oxidation in lamb meat. Meat Sci. 2014, 96, 1489–1493. [Google Scholar] [CrossRef] [PubMed]
- Lanza, M.; Scerra, M.; Bognanno, M.; Buccioni, A.; Cilione, C.; Biondi, L.; Priolo, A.; Luciano, G. Fatty acid metabolism in lambs fed citrus pulp. J. Anim. Sci. 2015, 93, 3179–3188. [Google Scholar] [CrossRef]
- Luciano, G.; Roscini, V.; Mattioli, S.; Ruggeri, S.; Gravador, R.S.; Natalello, A.; Lanza, M.; De Angelis, A.; Priolo, A. Vitamin E is the major contributor to the antioxidant capacity in lambs fed whole dried citrus pulp. Animal 2017, 11, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Schwartzkopf-Genswein, K.S.; Beauchemin, K.A.; Gibb, D.J.; Crews, D.H.; Hickman, D.D.; Streeter, M.; McAllister, T.A. Effect of bunk management on feeding behavior, ruminal acidosis and performance of feedlot cattle: A review. J. Anim. Sci. 2003, 81 (E-Suppl. 2), 149–158. [Google Scholar] [CrossRef]
- Schutz, J.; Wagner, J.; Neuhold, K.; Archibeque, S.; Engle, T. Effect of feed bunk management on feedlot steer intake1. Prof. Anim. Sci. 2011, 27, 395–401. [Google Scholar] [CrossRef]
- Crews, D.H.; Carstens, G.E. Measuring individual feed intake and utilization in growing cattle. In Feed Efficiency in the Beef Industry; Hill, R.A., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 21–28. [Google Scholar] [CrossRef]
- Harris, L. Compilación de Datos Analíticos y Biológicos en la Preparación de Cuadros de Composición de Alimentos para Uso en los Trópicos de América Latina; University of Florida Department of Animal Sciences: Gainesville, FL, USA, 1970. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Goering, H.; Van Soest, P.J. Forage Fiber Analysis (Apparatus, Reagents, Procedures and Some Applications). Agriculture Handbook No. 379; US Department of Agriculture: Washington, DC, USA, 1970. [Google Scholar]
- Weiss, W.; Conrad, H.; Pierre, N.S. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim. Feed. Sci. Technol. 1992, 39, 95–110. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine (NASEM). Nutrient Requirements of Beef Cattle, 8th rev. ed.; The National Academies Press: Washington, DC, USA, 2016; p. 475. [Google Scholar]
- USDA—United States Department of Agriculture. United States Standards for Grades of Carcass Beef. Agricultural Marketing Service. Livestock, Poultry and Seed Program. 2017. Available online: https://www.ams.usda.gov/sites/default/files/media/CarcassBeefStandard.pdf (accessed on 18 March 2021).
- American Meat Science Association. Research Guidelines for Cookery, Sensory Evaluation, and Instrumental Tenderness Measurements of Meat, 2nd ed.; Version 1.02; AMSA: Champaign, IL, USA, 2016; pp. 82–87. [Google Scholar]
- Xu, Z. Analysis of Tocopherols and Tocotrienols. Curr. Protoc. Food Anal. Chem. 2002, 4, D1.5.1–D1.5.12. [Google Scholar] [CrossRef]
- Bertolín, J.; Joy, M.; Rufino-Moya, P.; Lobón, S.; Blanco, M. Simultaneous determination of carotenoids, tocopherols, retinol and cholesterol in ovine lyophilised samples of milk, meat, and liver and in unprocessed/raw samples of fat. Food Chem. 2018, 257, 182–188. [Google Scholar] [CrossRef]
- Wu, C.; Duckett, S.; Neel, J.; Fontenot, J.; Clapham, W. Influence of finishing systems on hydrophilic and lipophilic oxygen radical absorbance capacity (ORAC) in beef. Meat Sci. 2008, 80, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Lenehan, C.; Moloney, A.; O’Riordan, E.; Kelly, A.; McGee, M. Comparison of rolled barley with citrus pulp as a supplement for growing cattle offered grass silage. Adv. Anim. Biosci. 2017, 8, s33–s37. [Google Scholar] [CrossRef]
- Gouvêa, V.N.; Batistel, F.; De Souza, J.; Chagas, L.J.; Sitta, C.; Campanili, P.R.B.; Galvani, D.B.; Pires, A.V.; Owens, F.N.; Santos, F.A.P. Flint corn grain processing and citrus pulp level in finishing diets for feedlot cattle. J. Anim. Sci. 2016, 94, 665–677. [Google Scholar] [CrossRef]
- Schaibly, G.E.; Wing, J.M. Effect of Roughage Concentrate Ratio on Digestibility and Rumen Fermentation of Corn Silage-Citrus Pulp Rations. J. Anim. Sci. 1974, 38, 697–701. [Google Scholar] [CrossRef]
- Owens, F.N.; Secrist, D.S.; Hill, W.J.; Gill, D.R. Acidosis in cattle: A review. J. Anim. Sci. 1998, 76, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.A.; Manteca, X.; Calsamiglia, S.; Schwartzkopf-Genswein, K.S.; Ferret, A. Ruminal acidosis in feedlot cattle: Interplay between feed ingredients, rumen function and feeding behavior (a review). Anim. Feed. Sci. Technol. 2012, 172, 66–79. [Google Scholar] [CrossRef]
- Filho, G.H.B.M.; Barbosa, F.A.; Emerick, L.L.; Souza, R.C.; Figueiredo, T.C.; Azevedo, H.D.O.; Cavalcanti, L.; Mandarino, R.A. Carcass and meat traits of feedlot Nellore bulls fed different dietary energy sources. Rev. Bras. Zootec. 2016, 45, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Duarte, M.D.S.; Paulino, P.V.R.; Filho, S.D.C.V.; Paulino, M.F.; Detmann, E.; Zervoudakis, J.T.; Monnerat, J.P.I.D.S.; Viana, G.D.S.; Silva, L.H.P.; Serão, N.V.L. Performance and meat quality traits of beef heifers fed with two levels of concentrate and ruminally undegradable protein. Trop. Anim. Health Prod. 2011, 43, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid–protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Dikeman, M.E. Fat reduction in animals and the effects on palatability and consumer acceptance of meat products. In Proceedings of the 40th Annual Reciprocal Meat Conference, Chicago, IL, USA, 28 June–1 July 1987; pp. 93–103. Available online: https://meatscience.org/docs/default-source/publications-resources/rmc/1987/fat-reduction-in-animals-and-the-effects-on-palatability-and-consumer-acceptance-of-meat-products.pdf?sfvrsn=2 (accessed on 5 April 2021).
- Miller, M.F.; Huffman, K.L.; Gilbert, S.Y.; Hamman, L.L.; Ramsey, C.B. Retail consumer acceptance of beef tenderized with calcium chloride. J. Anim. Sci. 1995, 73, 2308–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.F.; Carr, M.A.; Ramsey, C.B.; Crockett, K.L.; Hoover, L.C. Consumer thresholds for establishing the value of beef tenderness. J. Anim. Sci. 2001, 79, 3062–3068. [Google Scholar] [CrossRef]
- Faustman, C.; Cassens, R.G. The biochemical basis for discoloration in fresh meat: A review. J. Muscle Foods 1990, 1, 217–243. [Google Scholar] [CrossRef]
- Faustman, C.; Sun, Q.; Mancini, R.; Suman, S.P. Myoglobin and lipid oxidation interactions: Mechanistic bases and control. Meat Sci. 2010, 86, 86–94. [Google Scholar] [CrossRef]
- Chan, K.M.; Decker, E.A.; Feustman, C. Endogenous skeletal muscle antioxidants. Crit. Rev. Food Sci. Nutr. 1994, 34, 403–426. [Google Scholar] [CrossRef] [PubMed]
- Descalzo, A.; Sancho, A. A review of natural antioxidants and their effects on oxidative status, odor and quality of fresh beef produced in Argentina. Meat Sci. 2008, 79, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Tayengwa, T.; Chikwanha, O.; Gouws, P.; Dugan, M.E.; Mutsvangwa, T.; Mapiye, C. Dietary citrus pulp and grape pomace as potential natural preservatives for extending beef shelf life. Meat Sci. 2020, 162, 108029. [Google Scholar] [CrossRef] [PubMed]
- Salami, S.A.; O’Grady, M.N.; Luciano, G.; Priolo, A.; McGee, M.; Moloney, A.P.; Kerry, J.P. Quality indices and sensory attributes of beef from steers offered grass silage and a concentrate supplemented with dried citrus pulp. Meat Sci. 2020, 168, 108181. [Google Scholar] [CrossRef] [PubMed]
- Faustman, C.; Cassens, R.; Schaefer, D.; Buege, D.; Williams, S.; Scheller, K. Improvement of Pigment and Lipid Stability in Holstein Steer Beef by Dietary Supplementation with Vitamin E. J. Food Sci. 1989, 54, 858–862. [Google Scholar] [CrossRef]
- Arnold, R.N.; Arp, S.C.; Scheller, K.K.; Williams, S.N.; Schaefer, D.M. Tissue equilibration and subcellular distribution of vitamin E relative to myoglobin and lipid oxidation in displayed beef. J. Anim. Sci. 1993, 71, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Lanari, M.C.; Schaefer, D.M. A review of dietary vitamin E supplementation for improvement of beef quality. J. Anim. Sci. 1995, 73, 3131–3140. [Google Scholar] [CrossRef] [PubMed]
- Pouzo, L.; Descalzo, A.; Zaritzky, N.; Rossetti, L.; Pavan, E. Antioxidant status, lipid and color stability of aged beef from grazing steers supplemented with corn grain and increasing levels of flaxseed. Meat Sci. 2016, 111, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, M.; Haraoka, M.; Hatate, H.; Tanaka, R. High-Performance Liquid Chromatography with Fluorescence Detection for Simultaneous Analysis of Retinoids (Retinyl Palmitate, Retinyl Acetate, and Free Retinol) and α-, β-, γ-, and δ-Tocopherols in Foods. Food Anal. Methods 2017, 10, 92–99. [Google Scholar] [CrossRef]
- Yang, A.; Brewster, M.; Lanari, M.; Tume, R. Effect of vitamin E supplementation on α-tocopherol and β-carotene concentrations in tissues from pasture- and grain-fed cattle. Meat Sci. 2002, 60, 35–40. [Google Scholar] [CrossRef]
- Mahecha, L.; Dannenberger, D.; Nuernberg, K.; Nuernberg, G.; Hagemann, E.; Martin, J. Relationship between Lipid Peroxidation and Antioxidant Status in the Muscle of German Holstein Bulls Fed n-3 and n-6 PUFA-Enriched Diets. J. Agric. Food Chem. 2010, 58, 8407–8413. [Google Scholar] [CrossRef]
- Insani, E.; Eyherabide, A.; Grigioni, G.; Sancho, A.; Pensel, N.; Descalzo, A. Oxidative stability and its relationship with natural antioxidants during refrigerated retail display of beef produced in Argentina. Meat Sci. 2008, 79, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Descalzo, A.; Insani, E.; Biolatto, A.; Sancho, A.; García, P.; Pensel, N.; Josifovich, J. Influence of pasture or grain-based diets supplemented with vitamin E on antioxidant/oxidative balance of Argentine beef. Meat Sci. 2005, 70, 35–44. [Google Scholar] [CrossRef]
- Pickworth, C.L.; Loerch, S.C.; Kopec, R.; Schwartz, S.; Fluharty, F.L. Concentration of pro-vitamin A carotenoids in common beef cattle feedstuffs1. J. Anim. Sci. 2012, 90, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Agócs, A.; Nagy, V.; Szabó, Z.; Márk, L.; Ohmacht, R.; Deli, J. Comparative study on the carotenoid composition of the peel and the pulp of different citrus species. Innov. Food Sci. Emerg. Technol. 2007, 8, 390–394. [Google Scholar] [CrossRef]
- Oh, M.; Kim, E.-K.; Jeon, B.-T.; Tang, Y.; Kim, M.S.; Seong, H.-J.; Moon, S.-H. Chemical compositions, free amino acid contents and antioxidant activities of Hanwoo (Bos taurus coreanae) beef by cut. Meat Sci. 2016, 119, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.; Nieto, G.; Ros, G.; Martínez, G.N. Total antioxidant capacity of meat and meat products consumed in a reference ‘Spanish standard diet’. Int. J. Food Sci. Technol. 2014, 49, 2610–2618. [Google Scholar] [CrossRef]
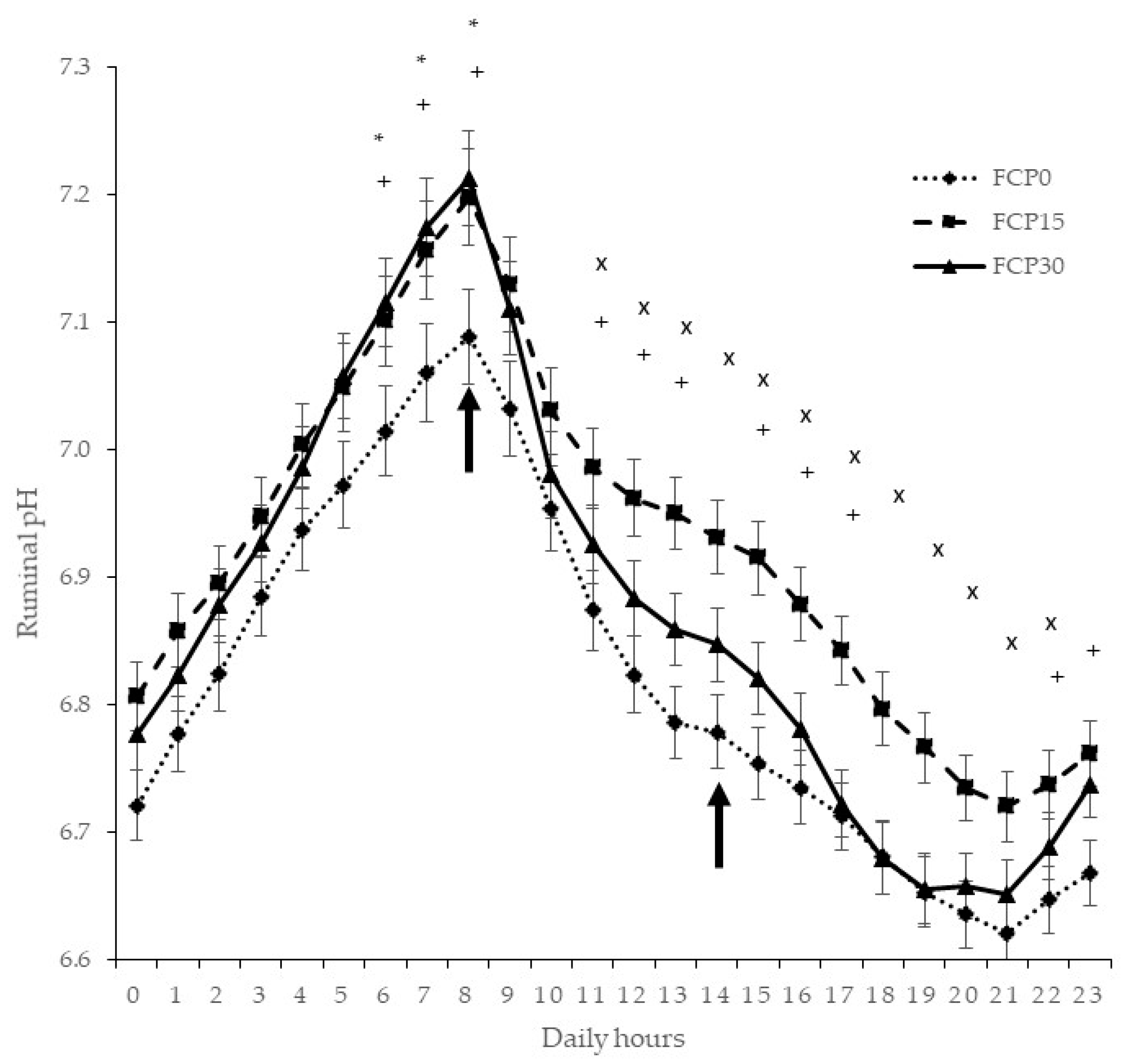

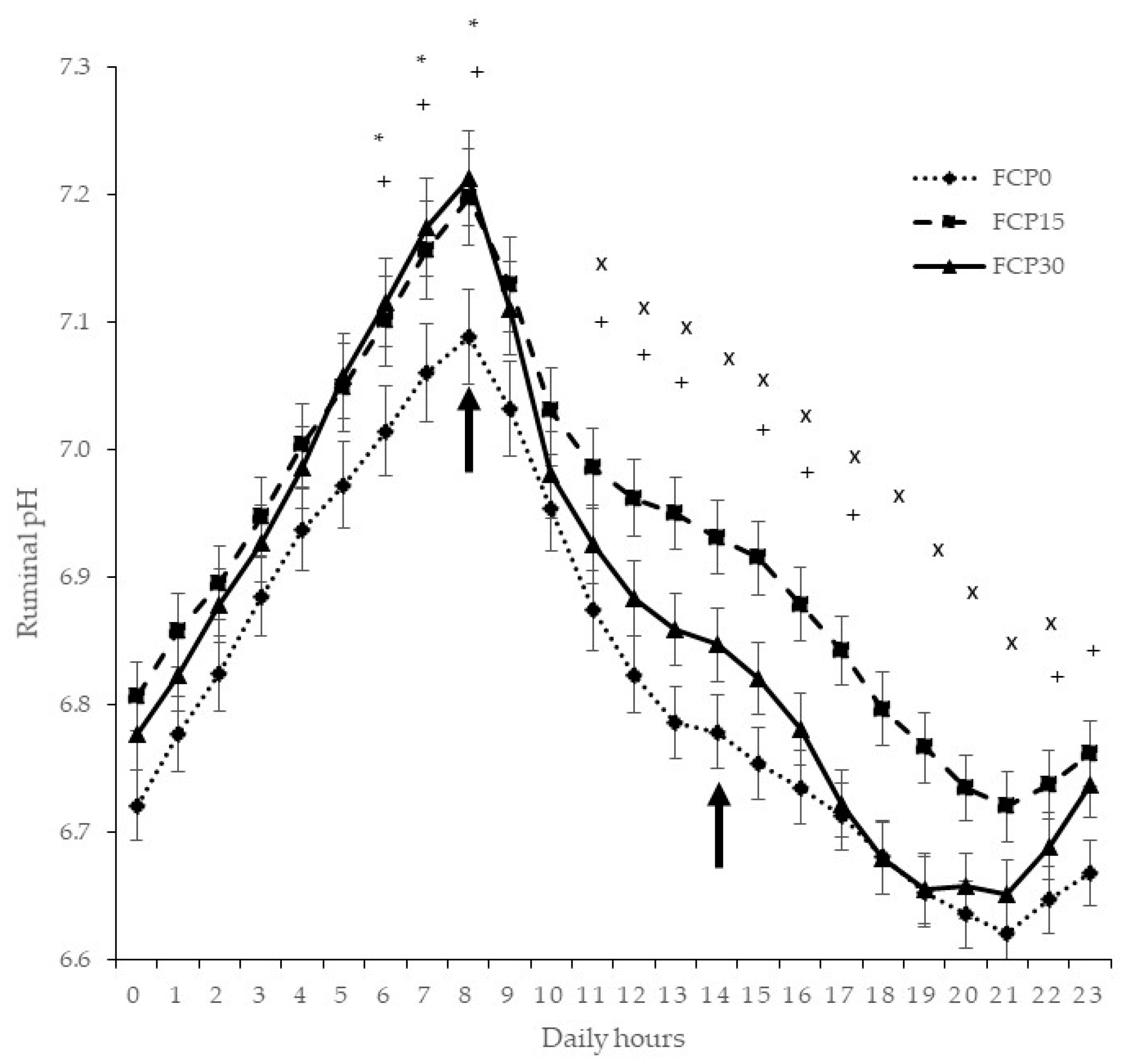{kind=link}
| Item | Dietary Treatments (Citrus Pulp as a Percentage of DM) | ||
|---|---|---|---|
| FCP0 | FCP15 | FCP30 | |
| Fresh citrus pulp (%) | 0.0 | 15.0 | 30.0 |
| Steam flake corn (%) | 19.0 | 19.0 | 19.0 |
| Corn grain (%) | 13.3 | 5.8 | 0.0 |
| Soybean meal (%) | 11.8 | 11.8 | 11.8 |
| Corn silage (%) | 40.0 | 31.1 | 20.6 |
| Wheat straw (%) | 13.9 | 15.1 | 16.3 |
| Premix (%) | 1.3 | 1.3 | 1.3 |
| Urea (%) | 0.67 | 0.83 | 0.98 |
| Premix 1 (%) | 1.3 | 1.3 | 1.3 |
| Physically effective 2 NDF (%) | 22.8 | 22.1 | 21.0 |
| Chemical Composition | |||
| DM (% as fed) | 51.90 | 39.60 | 32.40 |
| Ash (% of DM) | 5.31 | 6.22 | 7.07 |
| NDF (% of DM) | 31.21 | 31.19 | 32.16 |
| ADF (% of DM) | 20.02 | 21.97 | 23.65 |
| CP (% of DM) | 14.30 | 14.80 | 14.90 |
| EE (% of DM) | 3.21 | 3.03 | 2.90 |
| Lignin (% DM) | 2.49 | 2.56 | 2.56 |
| NEm 3 (Mcal/kg DM) | 1.76 | 1.71 | 1.68 |
| NEg 3 (Mcal/kg DM) | 1.13 | 1.10 | 1.07 |
| Dietary Treatments | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Item | FCP0 | FCP15 | FCP30 | Diet | Linear | Quadratic | FCP0 vs. FCP15 + 30 |
| Initial weight (kg) | 386.4 ± 7.97 | 385.1 ± 7.97 | 382.5 ± 8.19 | 0.8841 | 0.6278 | 0.9277 | 0.7015 |
| Final weight (kg) | 523.5 ± 10.6 | 520.1 ± 10.6 | 509.8 ± 11.0 | 0.4601 | 0.2328 | 0.7203 | 0.3761 |
| ADG (kg BW/day) 1 | 1.580 ± 0.06 | 1.535 ± 0.06 | 1.480 ± 0.06 | 0.3629 | 0.1594 | 0.9260 | 0.2293 |
| Dry matter intake (kg DM/day) | 11.80 a ± 0.33 | 11.28 a ± 0.33 | 10.31 b ± 0.35 | 0.0066 | 0.0019 | 0.5551 | 0.0101 |
| F:G (kg DM/kg BW) 2 | 7.51 a ± 0.15 | 7.37 a ± 0.15 | 6.98 b ± 0.16 | 0.0352 | 0.0168 | 0.4653 | 0.0699 |
| Water intake (lt/day) | 24.02 ± 0.79 | 23.20 ± 0.79 | 22.30 ± 0.82 | 0.4350 | 0.2309 | 0.6802 | 0.2084 |
| WI:DMI (lt/kg DM) 3 | 2.04 ± 0.79 | 2.07 ± 0.79 | 2.25 ± 0.82 | 0.0500 | 0.0231 | 0.3054 | 0.1169 |
| Dietary Treatments | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Item | FCP0 | FCP15 | FCP30 | Diet | Linear | Quadratic | FCP0 vs. FCP15 + 30 |
| Hot carcass weight (kg) 2 | 274.4 ± 2.1 | 271.6 ± 2.1 | 269.9 ± 2.2 | 0.3463 | 0.1546 | 0.8403 | 0.1732 |
| Marbling 1,3 | 407 ± 11.9 | 387 ± 11.7 | 397 ± 12.5 | 0.5113 | 0.5975 | 0.3144 | 0.3377 |
| Ribeye area 3 (cm2) | 63.2 ± 2.1 | 63.4 ± 2.1 | 63.1 ± 2.3 | 0.9941 | 0.9820 | 0.9156 | 0.9748 |
| Fat thickness 3 (mm) | 14.6 ± 1.2 | 12.7 ± 1.2 | 12.9 ± 1.3 | 0.4521 | 0.3407 | 0.4347 | 0.2221 |
| WBSF–5 day | 3.30 ± 0.23 | 3.11 ± 0.23 | 3.60 ± 0.25 | 0.3621 | 0.3920 | 0.2305 | 0.8482 |
| Cooking losses (%) | 22.5 ± 0.6 | 21.4 ± 0.6 | 21.9 ± 0.6 | 0.4058 | 0.4635 | 0.2626 | 0.2383 |
| L*–0 day | 38.9 ± 0.9 | 38.9 ± 0.9 | 38.8 ± 1.0 | 0.9911 | 0.9323 | 0.9172 | 0.9817 |
| a*–0 day | 22.4 ± 0.5 | 22.3 ± 0.5 | 21.6 ± 0.5 | 0.4233 | 0.2420 | 0.5521 | 0.4668 |
| b*–0 day | 11.8 ± 0.4 | 12.1 ± 0.4 | 11.4 ± 0.4 | 0.4103 | 0.4318 | 0.2769 | 0.8970 |
| L*–5 day | 39.8 ± 0.7 | 39.2 ± 0.7 | 38.3 ± 0.7 | 0.3672 | 0.1627 | 0.8843 | 0.2428 |
| a*–5 day | 22.2 ± 0.7 | 22.7 ± 0.7 | 22.0 ± 0.7 | 0.7415 | 0.7659 | 0.4749 | 0.9231 |
| b*–5 day | 10.6 ± 0.4 | 11.3 ± 0.4 | 10.4 ± 0.5 | 0.3476 | 0.7479 | 0.1580 | 0.6608 |
| Dietary Treatments | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Item | FCP0 | FCP15 | FCP30 | Diet | Linear | Quadratic | FCP0 vs. FCP15 + 30 |
| α-tocopherol 1 | 0.96 c ± 0.32 | 2.26 b ± 0.34 | 3.54 a ± 0.34 | <0.0001 | <0.0001 | 0.9722 | <0.0001 |
| Retinol 1 | 0.17 ± 0.03 | 0.15 ± 0.03 | 0.18 ± 0.03 | 0.8413 | 0.7693 | 0.6091 | 0.9885 |
| Retinoid 1 | 0.88 b ± 0.06 | 1.04 a,b ± 0.06 | 1.18 a ± 0.06 | 0.0044 | 0.0011 | 0.9096 | 0.0030 |
| β-carotene 1 | 1.38 a,b ± 0.08 | 1.53 a ± 0.08 | 1.17 b ± 0.08 | 0.0084 | 0.0585 | 0.0086 | 0.7564 |
| Carotenoids 1 | 2.99 ± 0.17 | 3.33 ± 0.18 | 2.83 ± 0.18 | 0.1506 | 0.5164 | 0.0649 | 0.6535 |
| DPPH 2 | 125.4 ± 2.6 | 117.9 ± 4.0 | 119.3 ± 2.6 | 0.1604 | 0.1899 | 0.3161 | 0.0563 |
| ORAC hydrophilic 2 | 414.6 ± 11.6 | 381.6 ± 11.6 | 393.3 ± 12.1 | 0.0999 | 0.1745 | 0.0975 | 0.0472 |
| ORAC lipophilic 2 | 91.9 b ± 4.7 | 113.8 a ± 4.5 | 95.0 b ± 4.7 | 0.0022 | 0.6065 | 0.0006 | 0.0241 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luzardo, S.; Banchero, G.; Ferrari, V.; Ibáñez, F.; Roig, G.; Aznárez, V.; Clariget, J.; La Manna, A. Effect of Fresh Citrus Pulp Supplementation on Animal Performance and Meat Quality of Feedlot Steers. Animals 2021, 11, 3338. https://doi.org/10.3390/ani11123338
Luzardo S, Banchero G, Ferrari V, Ibáñez F, Roig G, Aznárez V, Clariget J, La Manna A. Effect of Fresh Citrus Pulp Supplementation on Animal Performance and Meat Quality of Feedlot Steers. Animals. 2021; 11(12):3338. https://doi.org/10.3390/ani11123338
Chicago/Turabian StyleLuzardo, Santiago, Georgget Banchero, Virginia Ferrari, Facundo Ibáñez, Gonzalo Roig, Valentín Aznárez, Juan Clariget, and Alejandro La Manna. 2021. "Effect of Fresh Citrus Pulp Supplementation on Animal Performance and Meat Quality of Feedlot Steers" Animals 11, no. 12: 3338. https://doi.org/10.3390/ani11123338
APA StyleLuzardo, S., Banchero, G., Ferrari, V., Ibáñez, F., Roig, G., Aznárez, V., Clariget, J., & La Manna, A. (2021). Effect of Fresh Citrus Pulp Supplementation on Animal Performance and Meat Quality of Feedlot Steers. Animals, 11(12), 3338. https://doi.org/10.3390/ani11123338