Transcriptional Regulator YqeI, Locating at ETT2 Locus, Affects the Pathogenicity of Avian Pathogenic Escherichia coli

 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids and Growth Conditions
2.2. Antimicrobial Susceptibility Testing of AE81
2.3. Construction of the Mutant Strain and the Complemented Strain
2.4. Detection of Growth Curve
2.5. RNA Extraction and Transcriptional Profiling Assay
2.6. Differential Expression Analysis
2.7. Detection of Transcription Level by Quantitative Real-Time PCR
2.8. Micromorphology Observation by Transmission Electron Microscopy
2.9. Evaluation of Motility on Semisolid Medium
2.10. Serum Bactericidal Assay
2.11. Bacterial Adhesion Assay
2.12. Animal Infection Experiments In Vivo
2.13. Statistical Analysis
3. Results
3.1. Genome Sequencing Analysis and Antimicrobial Susceptibility of AE81
3.2. Absence of yqeI Did Not Affect the Growth of AE81
3.3. YqeI Participated in Multiple Bacterial Biological Pathways
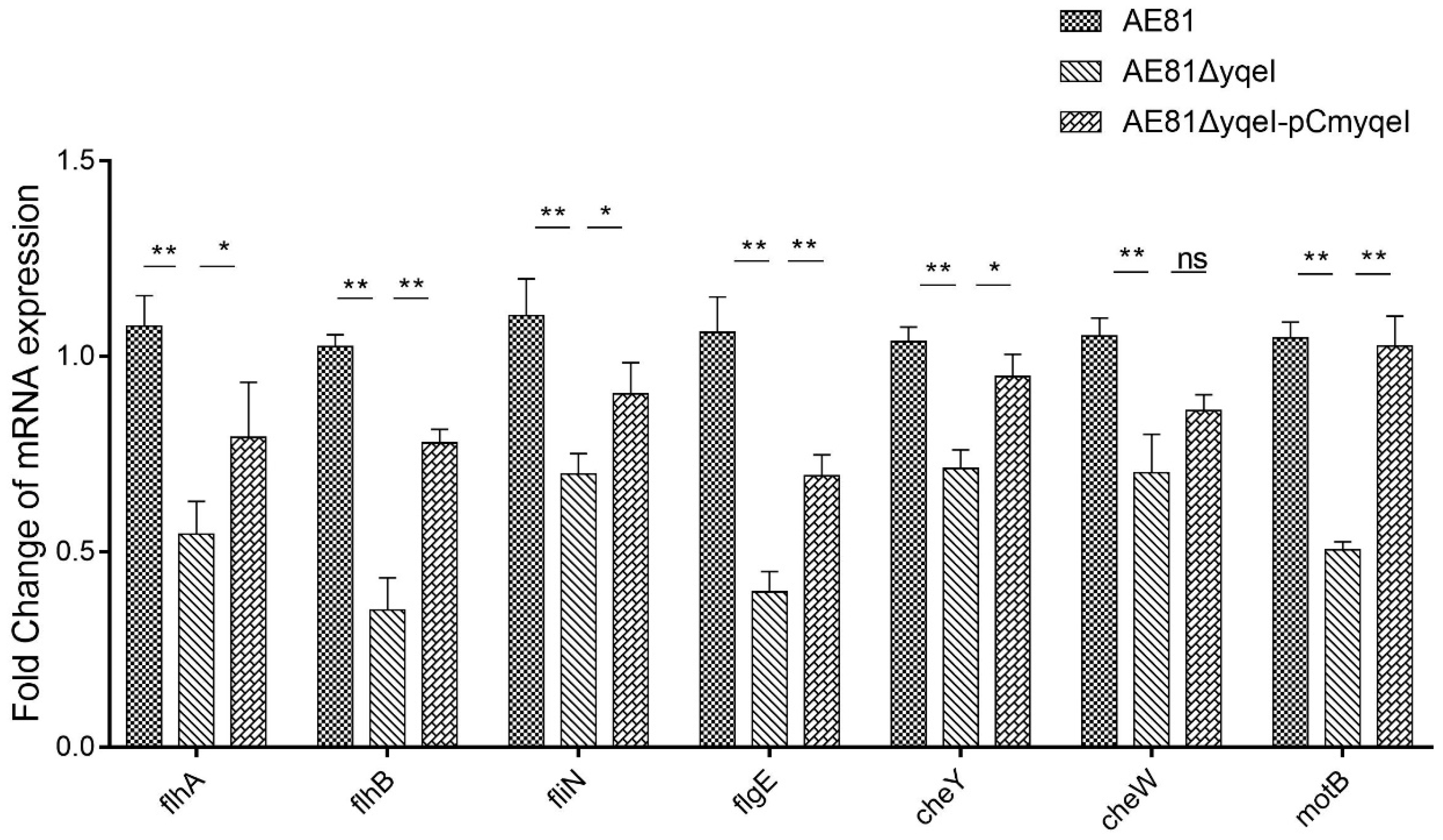
3.4. YqeI Up-Regulated the Expression of Flagella-Related Genes
3.5. Inactivation of yqeI Decreased the Formation of Flagella and Motility
3.6. YqeI Contributed to the Survival Ability of AE81 in the Serum Resistance
3.7. YqeI Affected the Adhesion Ability of AE81
3.8. Absence of yqeI Attenuated Bacterial Colonization during Systemic Infection In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Vandekerchove, D.; De Herdt, P.; Laevens, H.; Pasmans, F. Colibacillosis in caged layer hens: Characteristics of the disease and the aetiological agent. Avian Pathol. 2004, 33, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Kabir, S.M.L. Avian colibacillosis and salmonellosis: A closer look at epidemiology, pathogenesis, diagnosis, control and public health concerns. Int. J. Environ. Res. Public Health 2010, 7, 89–114. [Google Scholar] [CrossRef] [PubMed]
- Kemmett, K.; Williams, N.J.; Chaloner, G.; Humphrey, S.; Wigley, P.; Humphrey, T. The contribution of systemic Escherichia coli infection to the early mortalities of commercial broiler chickens. Avian Pathol. 2014, 43, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Dho-Moulin, M.; Fairbrother, J.M. Avian pathogenic Escherichia coli (APEC). Vet. Res. 1999, 30, 299–316. [Google Scholar]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Fakhr, M.K.; Nolan, L.K. Comparison of Escherichia coli isolates implicated in human urinary tract infection and avian colibacillosis. Microbiology (Reading) 2005, 151, 2097–2110. [Google Scholar] [CrossRef]
- Johnson, J.R.; Russo, T.A. Molecular epidemiology of extraintestinal pathogenic (uropathogenic) Escherichia coli. Int. J. Med. Microbiol. 2005, 295, 383–404. [Google Scholar] [CrossRef]
- Bonacorsi, S.; Bingen, E. Molecular epidemiology of Escherichia coli causing neonatal meningitis. Int. J. Med. Microbiol. 2005, 295, 373–381. [Google Scholar] [CrossRef]
- Manges, A.R.; Johnson, J.R. Food-Borne Origins of Escherichia coli Causing Extraintestinal Infections. Clin. Infect. Dis. 2012, 55, 712–719. [Google Scholar] [CrossRef]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef]
- Li, G.; Ewers, C.; Laturnus, C.; Diehl, I.; Alt, K.; Dai, J.; Antao, E.M.; Schnetz, K.; Wieler, L.H. Characterization of a yjjQ mutant of avian pathogenic Escherichia coli (APEC). Microbiology 2008, 154, 1082–1093. [Google Scholar] [CrossRef]
- Herren, C.D.; Mitra, A.; Palaniyandi, S.K.; Coleman, A.; Elankumaran, S.; Mukhopadhyay, S. The BarA-UvrY two-component system regulates virulence in avian pathogenic Escherichia coli O78:K80:H9. Infect. Immun. 2006, 74, 4900–4909. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Bai, H.; Liu, L.; Dong, H.; Liu, R.; Song, J.; Ding, C.; Qi, K.; Liu, H.; Yu, S. The luxS gene functions in the pathogenesis of avian pathogenic Escherichia coli. Microb. Pathog. 2013, 55, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Hartleib, S.; Prager, R.; Hedenstrom, I.; Lofdahl, S.; Tschape, H. Prevalence of the new, SPI1-like, pathogenicity island ETT2 among Escherichia coli. Int. J. Med. Microbiol. 2003, 292, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Makino, K.; Ohnishi, M.; Kurokawa, K.; Ishii, K.; Yokoyama, K.; Han, C.G.; Ohtsubo, E.; Nakayama, K.; Murata, T.; et al. Complete genome sequence of enterohemorrhagic Escherichia coli O157:H7 and genomic comparison with a laboratory strain K-12. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2001, 8, 11–22. [Google Scholar] [CrossRef]
- Wang, S.; Liu, X.; Xu, X.; Zhao, Y.; Yang, D.; Han, X.; Tian, M.; Ding, C.; Peng, D.; Yu, S. Escherichia coli type III secretion system 2 (ETT2) is widely distributed in avian pathogenic Escherichia coli isolates from Eastern China. Epidemiol. Infect. 2016, 144, 2824–2830. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.P.; Chaudhuri, R.R.; Fivian, A.; Bailey, C.M.; Antonio, M.; Barnes, W.M.; Pallen, M.J. The ETT2 gene cluster, encoding a second type III secretion system from Escherichia coli, is present in the majority of strains but has undergone widespread mutational attrition. J. Bacteriol. 2004, 186, 3547–3560. [Google Scholar] [CrossRef]
- Zhang, L.H.; Chaudhuri, R.R.; Constantinidou, C.; Hobman, J.L.; Patel, M.D.; Jones, A.C.; Sarti, D.; Roe, A.J.; Vlisidou, I.; Shaw, R.K.; et al. Regulators encoded in the Escherichia coli type III secretion system 2 gene cluster influence expression of genes within the locus for enterocyte effacement in enterohemorrhagic E. coli O157: H7. Infect. Immun. 2004, 72, 7282–7293. [Google Scholar] [CrossRef]
- Tukel, C.; Akcelik, M.; de Jong, M.F.; Simsek, O.; Tsolis, R.M.; Baumler, A.J. MarT activates expression of the MisL autotransporter protein of Salmonella enterica serotype Typhimurium. J. Bacteriol. 2007, 189, 3922–3926. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Zuo, J.; Tu, C.; Wang, Y.; Qi, K.; Hu, J.; Wang, Z.; Mi, R.; Yan, H.; Chen, Z.; Han, X. The role of the wzy gene in lipopolysaccharide biosynthesis and pathogenesis of avian pathogenic Escherichia coli. Microb. Pathog. 2019, 127, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Han, X.G.; Bai, H.; Tu, J.; Yang, L.J.; Xu, D.; Wang, S.H.; Qi, K.Z.; Fan, G.B.; Zhang, Y.X.; Zuo, J.K.; et al. Deletion of luxS further attenuates the virulence of the avian pathogenic Escherichia coli aroA mutant. Microb. Pathog. 2015, 88, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Shulman, A.; Yair, Y.; Biran, D.; Sura, T.; Otto, A.; Gophna, U.; Becher, D.; Hecker, M.; Ron, E.Z. The Escherichia coli Type III Secretion System 2 Has a Global Effect on Cell Surface. mBio 2018, 9, e01070. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xu, X.; Liu, X.; Wang, D.; Liang, H.; Wu, X.; Tian, M.; Ding, C.; Wang, G.; Yu, S. Escherichia coli type III secretion system 2 regulator EtrA promotes virulence of avian pathogenic Escherichia coli. Microbiol. Read. Engl. 2017, 163, 1515–1524. [Google Scholar] [CrossRef]
- Yao, Y.; Xie, Y.; Perace, D.; Zhong, Y.; Lu, J.; Tao, J.; Guo, X.; Kim, K.S. The type III secretion system is involved in the invasion and intracellular survival of Escherichia coli K1 in human brain microvascular endothelial cells. FEMS Microbiol. Lett. 2009, 300, 18–24. [Google Scholar] [CrossRef]
- Macnab, R.M. The bacterial flagellum: Reversible rotary propellor and type III export apparatus. J. Bacteriol. 1999, 181, 7149–7153. [Google Scholar] [CrossRef]
- Adams, P.; Fowler, R.; Kinsella, N.; Howell, G.; Farris, M.; Coote, P.; O’Connor, C.D. Proteomic detection of PhoPQ- and acid-mediated repression of Salmonella motility. Proteomics 2001, 1, 597–607. [Google Scholar] [CrossRef]
- Wiebe, H.; Gurlebeck, D.; Gross, J.; Dreck, K.; Pannen, D.; Ewers, C.; Wieler, L.H.; Schnetz, K. YjjQ Represses Transcription of flhDC and Additional Loci in Escherichia coli. J. Bacteriol. 2015, 197, 2713–2720. [Google Scholar] [CrossRef]
- De Pace, F.; de Paiva, J.B.; Nakazato, G.; Lancellotti, M.; Sircili, M.P.; Stehling, E.G.; da Silveira, W.D.; Sperandio, V. Characterization of IcmF of the type VI secretion system in an avian pathogenic Escherichia coli (APEC) strain. Microbiology 2011, 157, 2954–2962. [Google Scholar] [CrossRef]
- Ewers, C.; Janssen, T.; Wieler, L.H. Avian pathogenic Escherichia coli (APEC). Berl. Munch. Tierarztl. Wochenschr. 2003, 116, 381–395. [Google Scholar] [PubMed]
- Mellata, M.; Dho-Moulin, M.; Dozois, C.M.; Curtiss, R.; Brown, P.K.; Arne, P.; Bree, A.; Desautels, C.; Fairbrother, J.M. Role of virulence factors in resistance of avian pathogenic Escherichia coli to serum and in pathogenicity. Infect. Immun. 2003, 71, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Xu, H.; Wang, X.; Zhang, D.; Ye, Z.; Gao, S.; Liu, X. RfaH promotes the ability of the avian pathogenic Escherichia coli O2 strain E058 to cause avian colibacillosis. J. Bacteriol. 2013, 195, 2474–2480. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Woodward, M.J. Virulence factors of Escherichia coli serotypes associated with avian colisepticaemia. Res. Vet. Sci. 2002, 73, 27–35. [Google Scholar] [CrossRef]
- Soto, G.E.; Hultgren, S.J. Bacterial adhesins: Common themes and variations in architecture and assembly. J. Bacteriol. 1999, 181, 1059–1071. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmid | Genotype or Description | Source |
|---|---|---|
| Strains | ||
| AE81 | APEC clinical strain, isolated from lung | Laboratory stock |
| AE81ΔyqeI | AE81 yqeI deletion mutant | This study |
| AE81ΔyqeI-pCmyqeI | AE81ΔyqeI with the plasmid pCmyqeI, Cmr 1 | This study |
| Plasmid | ||
| pCmyqeI | pSTV28 with yqeI gene, Cmr 1 | This study |
| Primer | Sequence (5′-3′) | Product Length (bp) |
|---|---|---|
| yqeI-up-cm | f: TGATGGTCTTGATGTTGCC r: CCAGCCTACAGCTCAGGCCAGAACTCGATA | 635 |
| yqeI-down-cm | f: TATTCATATGTAACCCTGTCTTACCGTG r: TTCCACTCAACGCAACAG | 242 |
| pKD3-yqeI | f: TGGCCTGAGCTGTAGGCTGGAGCTGCTT r: GACAGGGTTACATATGAATATCCTCCTTAG | 1033 |
| C-yqeI | f: CCGGAATTCCATGTACTGGATTATTAACGA r: CGCGGATCCTCAACCACGATTAACCTCAC | 870 |
| yqeI-out | f: TTCGGTCCAACATTGATA r: TAAGATTTGCTCGTCCCT | 1309 |
| flgE-rt | f: TCAGTCGGACGACATGGTAGAT r: CGAGTATGTTGCGAATGTGGAT | 148 |
| fliN-rt | f: CGGTGGTGATGTCAGCGG r: GCTCTTTGATGGTCATTCGTGTG | 104 |
| flhA-rt | f: GAAGTGGTGGTCGTTGCCGATAAA r: GCTGGAGTTGAAGATAATGGGGG | 121 |
| flhB-rt | f: CCTACCAAATCCATCGCATTACCC r: CTCCTGGTTGGCAGCGTGA | 113 |
| cheY-rt | f: TAACACTGGCATTGCCGACATCG r: AATGTTGAGGAAGCGGAAGATGGC | 159 |
| motB-rt | f: GGCGCTTACTGGCTCATTCTGG r: GCCGTCAACCGTCGCATCAG | 99 |
| cheW-rt | f: CACATCCACCTGGCTGAACT r: AAAGTGCAGGAGATCCGTGG | 128 |
| Genes | Symbol | Description | Log2FoldChange |
|---|---|---|---|
| Ecs_0681 | ybeL | methyl-accepting chemotaxis protein II, aspartate sensor receptor | 1.50 |
| Ecs_1448 | flgN | flagella synthesis protein FlgN | −1.29 |
| Ecs_1449 | flgM | negative regulator of flagellin synthesis FlgM | −1.43 |
| Ecs_1454 | flgE | flagellar hook protein FlgE | −1.06 |
| Ecs_1460 | flgK | flagellar hook-associated protein 1 FlgK | −1.23 |
| Ecs_2589 | flhA_2 | flagellar biosynthesis protein FlhA | −1.54 |
| Ecs_2590 | flhB | flagellar biosynthetic protein FlhB | −1.32 |
| Ecs_2591 | cheZ | chemotaxis protein CheZ | −1.69 |
| Ecs_2592 | cheY | two-component system, chemotaxis family, chemotaxis protein CheY | −1.67 |
| Ecs_2596 | tsr_1 | methyl-accepting chemotaxis protein II, aspartate sensor receptor | −1.14 |
| Ecs_2597 | cheW | purine-binding chemotaxis protein CheW | −1.03 |
| Ecs_2599 | motB | chemotaxis protein MotB | −1.20 |
| Ecs_2600 | motA | chemotaxis protein MotA | −1.48 |
| Ecs_2601 | flhC | flagellar transcriptional activator FlhC | −1.39 |
| Ecs_2602 | flhD | flagellar transcriptional activator FlhD | −1.20 |
| Ecs_2662 | fliC | flagellin | −1.60 |
| Ecs_2663 | fliD | flagellar hook-associated protein 2 | −1.04 |
| Ecs_2665 | fliT | flagellar protein FliT | −1.14 |
| Ecs_2680 | fliI | flagellum-specific ATP synthase | −1.00 |
| Ecs_2681 | fliJ | flagellar FliJ protein | −2.53 |
| Ecs_2682 | fliK | flagellar hook-length control protein FliK | −1.23 |
| Ecs_2684 | fliM | flagellar motor switch protein FliM | −1.47 |
| Ecs_2685 | fliN | flagellar motor switch protein FliN/FliY | −2.61 |
| Ecs_2686 | fliO | flagellar protein FliO/FliZ | −2.82 |
| Ecs_3042 | mglB | methyl-galactoside transport system substrate-binding protein | 2.13 |
| Ecs_4424 | dppA | dipeptide transport system substrate-binding protein | 2.18 |
| Ecs_4437 | yiaD | OmpA-OmpF porin, OOP family | −1.17 |
| Ecs_5017 | malE | maltose/maltodextrin transport system substrate-binding protein | 2.82 |
| Ecs_5315 | tsr | methyl-accepting chemotaxis protein I, serine sensor receptor | −1.51 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, M.; Xiao, Y.; Fu, D.; Raheem, M.A.; Shao, Y.; Song, X.; Tu, J.; Xue, T.; Qi, K. Transcriptional Regulator YqeI, Locating at ETT2 Locus, Affects the Pathogenicity of Avian Pathogenic Escherichia coli. Animals 2020, 10, 1658. https://doi.org/10.3390/ani10091658
Xue M, Xiao Y, Fu D, Raheem MA, Shao Y, Song X, Tu J, Xue T, Qi K. Transcriptional Regulator YqeI, Locating at ETT2 Locus, Affects the Pathogenicity of Avian Pathogenic Escherichia coli. Animals. 2020; 10(9):1658. https://doi.org/10.3390/ani10091658
Chicago/Turabian StyleXue, Mei, Yating Xiao, Dandan Fu, Muhammad Akmal Raheem, Ying Shao, Xiangjun Song, Jian Tu, Ting Xue, and Kezong Qi. 2020. "Transcriptional Regulator YqeI, Locating at ETT2 Locus, Affects the Pathogenicity of Avian Pathogenic Escherichia coli" Animals 10, no. 9: 1658. https://doi.org/10.3390/ani10091658
APA StyleXue, M., Xiao, Y., Fu, D., Raheem, M. A., Shao, Y., Song, X., Tu, J., Xue, T., & Qi, K. (2020). Transcriptional Regulator YqeI, Locating at ETT2 Locus, Affects the Pathogenicity of Avian Pathogenic Escherichia coli. Animals, 10(9), 1658. https://doi.org/10.3390/ani10091658