Effects of 5-Aminolevulinic Acid as a Supplement on Animal Performance, Iron Status, and Immune Response in Farm Animals: A Review


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Literature Search
2.2. Inclusion and Exclusion Criteria
2.3. Data Extraction
2.4. Quality Criteria
3. Results
3.1. Literature Search
3.2. Description of Included Studies
3.3. Effects of 5-ALA on Animal Performance
3.4. Effect of 5-ALA on Iron Status
3.5. Effect of 5-ALA on Immune Response
3.6. Effects of 5-ALA Compared with Other Alternative Treatments
3.7. Mechanism of Action of 5-ALA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Fund
Conflicts of Interest
References
- Chattopadhyay, M.K. Use of antibiotics as feed additives: A burning question. Front. Microbiol. 2014, 5, 334. [Google Scholar] [CrossRef] [PubMed]
- Sato, K. Molecular nutrition: Interaction of nutrients, gene regulations and performances. Anim. Sci. J. 2016, 87, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Weekes, T.L.; Luimes, P.H.; Cant, J.P. Responses to amino acid imbalances and deficiencies in lactating dairy cows. J. Dairy Sci. 2006, 89, 2177–2187. [Google Scholar] [CrossRef]
- Mann, G.R.; Duncan, S.E.; Knowlton, K.F.; Dietrich, A.D.; O’Keefe, S.F. Effects of mineral content of bovine drinking water: Does iron content affect milk quality? J. Dairy Sci. 2013, 96, 7478–7489. [Google Scholar] [CrossRef]
- Genther, O.N.; Beede, D.K. Preference and drinking behavior of lactating dairy cows offered water with different concentrations, valences, and sources of iron. J. Dairy Sci. 2013, 96, 1164–1176. [Google Scholar] [CrossRef]
- Fujino, M.; Nishio, Y.; Ito, H.; Tanaka, T.; Li, X.K. 5-Aminolevulinic acid regulates the inflammatory response and alloimmune reaction. Int. Immunopharmacol. 2016, 37, 71–78. [Google Scholar] [CrossRef]
- Zhu, Y.; Hon, T.; Ye, W.; Zhang, L. Heme deficiency interferes with the Ras-mitogen-activated protein kinase signaling pathway and expression of a subset of neuronal genes. Cell Growth Differ. 2002, 13, 431–439. [Google Scholar]
- Chen, Y.J.; Kim, I.H.; Cho, J.H.; Min, B.J.; Yoo, J.S.; Wang, Q. Effect of δ-aminolevulinic acid on growth performance, nutrient digestibility, blood parameters and the immune response of weanling pigs challenged with Escherichia coli lipopolysaccharide. Livest. Sci. 2008, 114, 108–116. [Google Scholar] [CrossRef]
- Sasaki, K.; Tanaka, T.; Nagai, S. Use of Photosynthetic Bacteria for the Production of SCP and Chemicals from Organic Wastes; Martin, A.M., Ed.; Springer: Boston, MA, USA, 1998; pp. 247–292. [Google Scholar] [CrossRef]
- Sasaki, K.; Watanabe, M.; Tanaka, T.; Tanaka, T. Biosynthesis, biotechnological production and applications of 5-aminolevulinic acid. Appl. Microbiol. Biotechnol. 2002, 58, 23–29. [Google Scholar] [CrossRef]
- Vansevičiute, R.; Venius, J.; Letautiene, S. 5-Aminolevulinic acid-based fluorescence diagnostics of cervical preinvasive changes. Medicina 2014, 50, 137–143. [Google Scholar] [CrossRef]
- Nakamura, M.; Nishikawa, J.; Hamabe, K.; Goto, A.; Nishimura, J.; Shibata, H.; Nagao, M.; Sasaki, S.; Hashimoto, S.; Okamoto, T.; et al. Preliminary study of photodynamic diagnosis using 5-aminolevulinic acid in gastric and colorectal tumors. World J. Gastroenterol. 2015, 21, 6706–6712. [Google Scholar] [CrossRef] [PubMed]
- Mateo, R.D.; Morrow, J.L.; Dailey, J.W.; Ji, F.; Kim, S.W. Use of δ-aminolevulinic acid in swine diet: Effect on growth performance, behavioral characteristics and hematological/immune status in nursery pigs. Asian-Aust. J. Anim. Sci. 2006, 19, 97–101. [Google Scholar] [CrossRef]
- Wang, J.P.; Kim, H.J.; Chen, Y.J.; Yoo, J.S.; Cho, J.H.; Kang, D.K.; Hyun, Y.; Kim, I.H. Effects of delta-aminolevulinic acid and vitamin C supplementation on feed intake, backfat, and iron status in sows. J. Anim. Sci. 2009, 87, 3589–3595. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Yan, L.; Lee, J.H.; Zhou, T.X.; Kim, I.H. Effects of dietary delta-aminolevulinic acid and vitamin C on growth performance, immune organ weight and ferrum status in broiler chicks. Livest. Sci. 2011, 135, 148–152. [Google Scholar] [CrossRef]
- Sato, K.; Matsushita, K.; Takahashi, K.; Aoki, M.; Fuziwara, J.; Miyanari, S.; Kamada, T. Dietary supplementation with 5-aminolevulinic acid modulates growth performance and inflammatory responses in broiler chickens. Poult. Sci. 2012, 91, 1582–1589. [Google Scholar] [CrossRef]
- Wang, J.P.; Jung, J.H.; Kim, I.H. Effects of dietary supplementation with delta-aminolevulinic acid on growth performance, hematological status, and immune responses of weanling pigs. Livest. Sci. 2011, 140, 131–135. [Google Scholar] [CrossRef]
- Hossain, M.M.; Park, J.W.; Kim, I.H. δ-Aminolevulinic acid, and lactulose supplements in weaned piglets diet: Effects on performance, fecal microbiota, and in-vitro noxious gas emissions. Livest. Sci. 2016, 183, 84–91. [Google Scholar] [CrossRef]
- Chen, Y.J.; Cho, J.H.; Yoo, J.S.; Wang, Y.; Huang, Y.; Kim, I.H. Evaluation of δ-aminolevulinic acid on serum iron status, blood characteristics, egg performance and quality in laying hens. Asian-Australas. J. Anim. Sci. 2008, 21, 1355–1360. [Google Scholar] [CrossRef]
- Wang, J.P.; Lee, J.H.; Jang, H.D.; Yan, L.; Cho, J.H.; Kim, I.H. Effects of δ-aminolevulinic acid and vitamin C supplementation on iron status, production performance, blood characteristics and egg quality of laying hens. J. Anim. Physiol. Anim. Nutr. 2011, 95, 417–423. [Google Scholar] [CrossRef]
- Yan, L.; Lee, J.H.; Meng, Q.W.; Ao, X.; Kim, I.H. Evaluation of dietary supplementation of delta-aminolevulinic acid and chito-oligosaccharide on production performance, egg quality and hematological characteristics in laying hens. Asian-Aust. J. Anim. Sci. 2010, 23, 1028–1033. [Google Scholar] [CrossRef]
- Hendawy, A.O.; Shiraishi, M.; Takeya, H.; Sugimura, S.; Miyanari, S.; Taniguchi, S.; Sato, K. Effects of 5-aminolevulinic acid supplementation on milk production, iron status, and immune response of dairy cows. J. Dairy Sci. 2019, 102, 11009–11015. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Li, T.S.; Kim, I.H. Dietary supplementation of delta-aminolevulinic acid to lactating sows improves growth performance and concentration of iron and hemoglobin of suckling piglets. Indian J. Anim. Sci. 2016, 86, 781–785. [Google Scholar]
- Cho, J.H.; Kim, I.H. δ-aminolevulinic acid (ALA) as a potential feed additive in pig: A review. J. Anim. Vet. Adv. 2011, 10, 1627–1630. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Academia and Clinic Annals of Internal Medicine Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. Annu. Intern. Med. 2009, 151, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.S.S.; Garbossa, C.A.P.; Oberlender, G.; Pereira, L.J.; Zangeronimo, M.G.; Sousa, R.V.; Cantarelli, V.S. Effect of ractopamine on lipid metabolism in vivo—A systematic review. Braz. Arch. Biol. Technol. 2013, 56, 35–43. [Google Scholar] [CrossRef]
- Retes, P.L.; Clemente, A.H.S.; Neves, D.G.; Espósito, M.; Makiyama, L.; Alvarenga, R.R.; Pereira, L.J.; Zangeronimo, M.G. In ovo feeding of carbohydrates for broilers—A systematic review. J. Anim. Physiol. Anim. Nutr. 2018, 102, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Kim, I.H. Effects of iron injection at birth on neonatal iron status in young pigs from first-parity sows fed delta-aminolevulinic acid. Anim. Feed Sci. Technol. 2012, 178, 151–157. [Google Scholar] [CrossRef]
- Yan, L.; Kim, I.H. Evaluation of dietary supplementation of delta-aminolevulinic acid and chitooligosaccharide on growth performance, nutrient digestibility, blood characteristics, and fecal microbial shedding in weaned pigs. Anim. Feed Sci. Technol. 2011, 169, 275–280. [Google Scholar] [CrossRef]
- Kang, S.N.; Chu, G.M.; Song, Y.M.; Jin, S.K.; Hwang, I.H.; Kim, I.S. The effects of replacement of antibiotics with by-products of oriental medicinal plants on growth performance and meat qualities in fattening pigs. Anim. Sci. J. 2012, 83, 245–251. [Google Scholar] [CrossRef]
- Chen, Y.J.; Kim, I.H.; Cho, J.H.; Yoo, J.S.; Kim, H.J.; Shin, S.O. Utilization of δ-aminolevulinic acid for livestock: Blood characteristics and immune organ weight in broilers. J. Anim. Feed Sci. 2008, 17, 215–223. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine: 10th Revised Edition; The National Academies Press: Washington, DC, USA, 1998; p. 212. [Google Scholar] [CrossRef]
- Yu, B.; Huang, W.-J.; Chiou, P.W.-S. Bioavailability of iron from amino acid complex in weanling pigs. Anim. Feed Sci. Technol. 2000, 86, 39–52. [Google Scholar] [CrossRef]
- Ponka, P. Tissue-specific regulation of iron metabolism and heme synthesis: Distinct control mechanisms in erythroid cells. Blood 1997, 89, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Gibson, S.L.; Foster, T.H.; Hilf, R. Effectiveness of delta-aminolevulinic acid-induced protoporphyrin as a photosensitizer for photodynamic therapy in vivo. Cancer Res. 1995, 55, 1723–1731. [Google Scholar] [PubMed]
- Jin, Z.; Kim, K.E.; Shin, H.J.; Jeong, E.A.; Park, K.A.; Lee, J.Y.; An, H.S.; Choi, E.B.; Jeong, J.H.; Kwak, W.; et al. Hippocampal lipocalin 2 is associated with neuroinflammation and iron-related oxidative stress in ob/ob mice. J. Neuropathol. Exp. Neurol. 2020, 79, 530–541. [Google Scholar] [CrossRef]
- Nishio, Y.; Fujino, M.; Zhao, M.; Ishii, T.; Ishizuka, M.; Ito, H.; Takahashi, K.; Abe, F.; Nakajima, M.; Tanaka, T.; et al. 5-Aminolevulinic acid combined with ferrous iron enhances the expression of heme oxygenase-1. Int. Immunopharmacol. 2014, 19, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, Y.; Hiraiwa, Y.; Hagiya, Y.; Nakajima, M.; Tanaka, T.; Ogura, S. 5-Aminolevulinic acid regulates the immune response in LPS-stimulated RAW 264.7 macrophages. BMC Immunol. 2018, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.D.; Riches, D.W.H. IFN-γ + LPS induction of iNOS is modulated by ERK, JNK/SAPK, and p38mapk in a mouse macrophage cell line. Am. J. Physiol.-Cell Physiol. 2001, 280, C441–C450. [Google Scholar] [CrossRef]
- Inoue, Y.; Hara, H.; Mitsugi, Y.; Yamaguchi, E.; Kamiya, T.; Itoh, A.; Adachi, T. 4-Hydroperoxy-2-decenoic acid ethyl ester protects against 6-hydroxydopamine-induced cell death via activation of Nrf2-ARE and eIF2α-ATF4 pathways. Neurochem. Int. 2018, 112, 288–296. [Google Scholar] [CrossRef]
- Ali, A.M.; Kunugi, H. Bee honey protects astrocytes against oxidative stress: A preliminary in vitro investigation. Neuropsychopharmacol. Rep. 2019, 39, 312–314. [Google Scholar] [CrossRef]
- Kunugi, H.; Mohammed Ali, A. Royal Jelly and Its Components Promote Healthy Aging and Longevity: From Animal Models to Humans. Int. J. Mol. Sci. 2019, 20, 4662. [Google Scholar] [CrossRef]
- Poss, K.D.; Tonegawa, S. Reduced stress defense in heme oxygenase 1-deficient cells. Proc. Natl. Acad. Sci. USA 1997, 94, 10925–10930. [Google Scholar] [CrossRef]
- Yachie, A.; Niida, Y.; Wada, T.; Igarashi, N.; Kaneda, H.; Toma, T.; Ohta, K.; Kasahara, Y.; Koizumi, S. Oxidative stress causes enhanced endothelial cell injury in human heme oxygenase-1 deficiency. J. Clin. Investig. 1999, 103, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Matthias, H.K.; Clive, W.; Todd, B.; Martha, C.; Tamir, M.E.; Mark, A.A.; Anupam, A. Heme oxygenase-1 modulates early inflammatory responses: Evidence from the heme oxygenase-1-deficient mouse. Am. J. Pathol. 2004, 165, 1045–1053. [Google Scholar] [CrossRef]
- Longtao, Z.; Zhongrong, L.; Furong, M.; Jing, L.; Yulan, F. Advance in the application of 5-aminolevulinic acid in the regulation of animal iron metabolism. Chin. Agric. Sci. Bull. 2012, 29, 1–4. [Google Scholar]
- Rincker, M.J.; Hill, G.M.; Link, J.E.; Rowntree, J.E. Effects of dietary iron supplementation on growth performance, hematological status, and whole-body mineral concentrations of nursery pigs. J. Anim. Sci. 2004, 82, 3189–3197. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]





{kind=link}
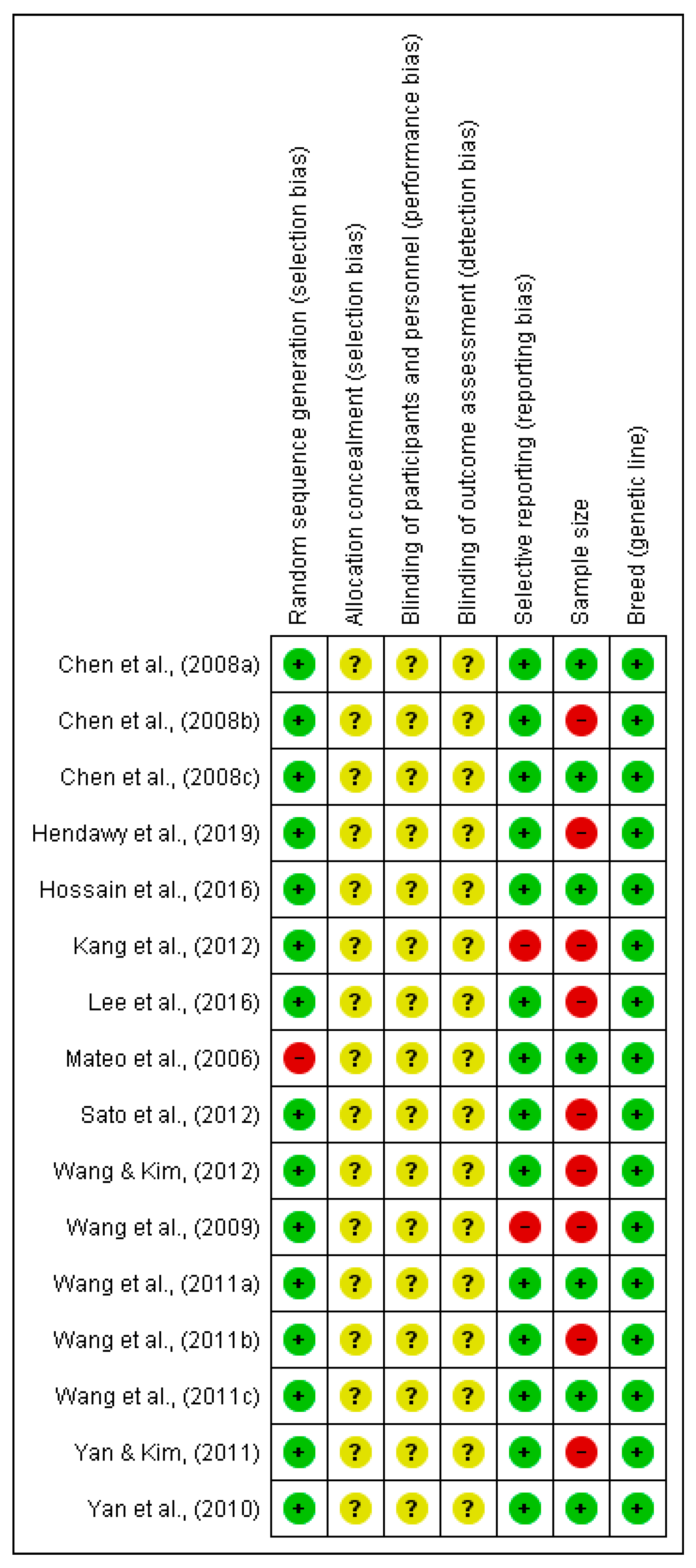{kind=link}
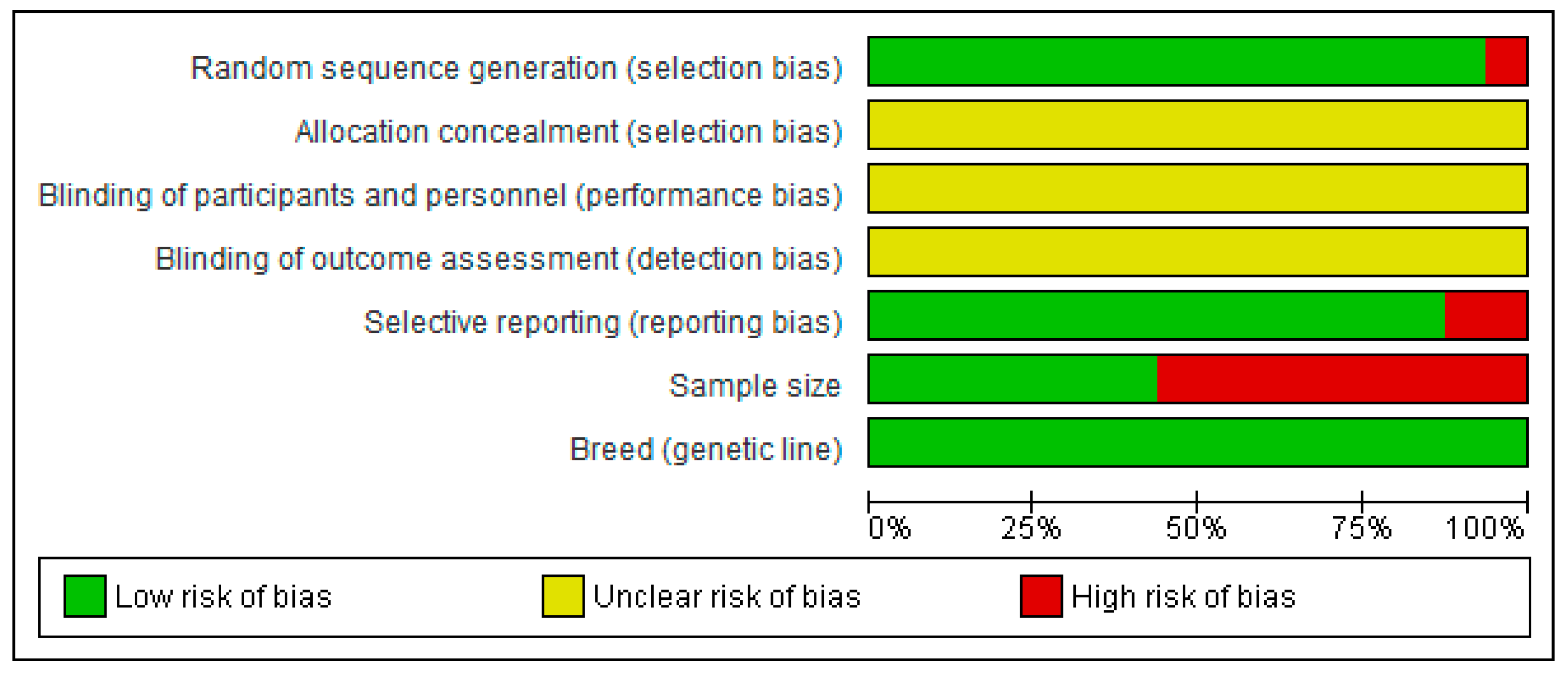{kind=link}
{kind=link}
| Species | Growth Performance | Production Performance | Immune Function | Iron Status | Ref. |
|---|---|---|---|---|---|
| Broiler chickens | 0 | ND | + | 0 | [31] |
| Broiler chickens | + | ND | + | ND | [16] |
| Broiler chickens | 0 | ND | 0 | + | [15] |
| Laying hens | ND | + | 0 | 0 | [19] |
| Laying hens | ND | + | + | + | [20] |
| Laying hens | ND | + | + | + | [21] |
| Weanling pigs | 0 | ND | + | + | [8] |
| Weanling pigs | + | ND | ND | + | [18] |
| Weanling pigs | 0 | ND | 0 | + | [13] |
| Weanling pigs | 0 | ND | + | + | [17] |
| Weanling pigs | 0 | ND | + | + | [29] |
| Fattening pigs | 0 | ND | ND | ND | [30] |
| Pregnant and lactating sows▲ | + | + | ND | + | [23] |
| Pregnant and lactating sows▲ | + | 0 | ND | + | [28] |
| Parturient sows▲ | + | + | + | + | [14] |
| Lactating dairy cows | ND | + | + | 0 | [22] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hendawy, A.O.; Khattab, M.S.; Sugimura, S.; Sato, K. Effects of 5-Aminolevulinic Acid as a Supplement on Animal Performance, Iron Status, and Immune Response in Farm Animals: A Review. Animals 2020, 10, 1352. https://doi.org/10.3390/ani10081352
Hendawy AO, Khattab MS, Sugimura S, Sato K. Effects of 5-Aminolevulinic Acid as a Supplement on Animal Performance, Iron Status, and Immune Response in Farm Animals: A Review. Animals. 2020; 10(8):1352. https://doi.org/10.3390/ani10081352
Chicago/Turabian StyleHendawy, Amin Omar, Mostafa Sayed Khattab, Satoshi Sugimura, and Kan Sato. 2020. "Effects of 5-Aminolevulinic Acid as a Supplement on Animal Performance, Iron Status, and Immune Response in Farm Animals: A Review" Animals 10, no. 8: 1352. https://doi.org/10.3390/ani10081352
APA StyleHendawy, A. O., Khattab, M. S., Sugimura, S., & Sato, K. (2020). Effects of 5-Aminolevulinic Acid as a Supplement on Animal Performance, Iron Status, and Immune Response in Farm Animals: A Review. Animals, 10(8), 1352. https://doi.org/10.3390/ani10081352