Relationship between the Quality of Colostrum and the Formation of Microflora in the Digestive Tract of Calves

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microbiological Assay
2.2. Chemical Analyses of Colostrum
2.3. Statistical Analyses
- 1st class colostrum density 1.070 g/cm3: 23 calves
- 2nd class colostrum density 1.057–1.070 g/cm3: 25 calves
- 3rd class colostrum density 1.044–1.056 g/cm3: 27 calves
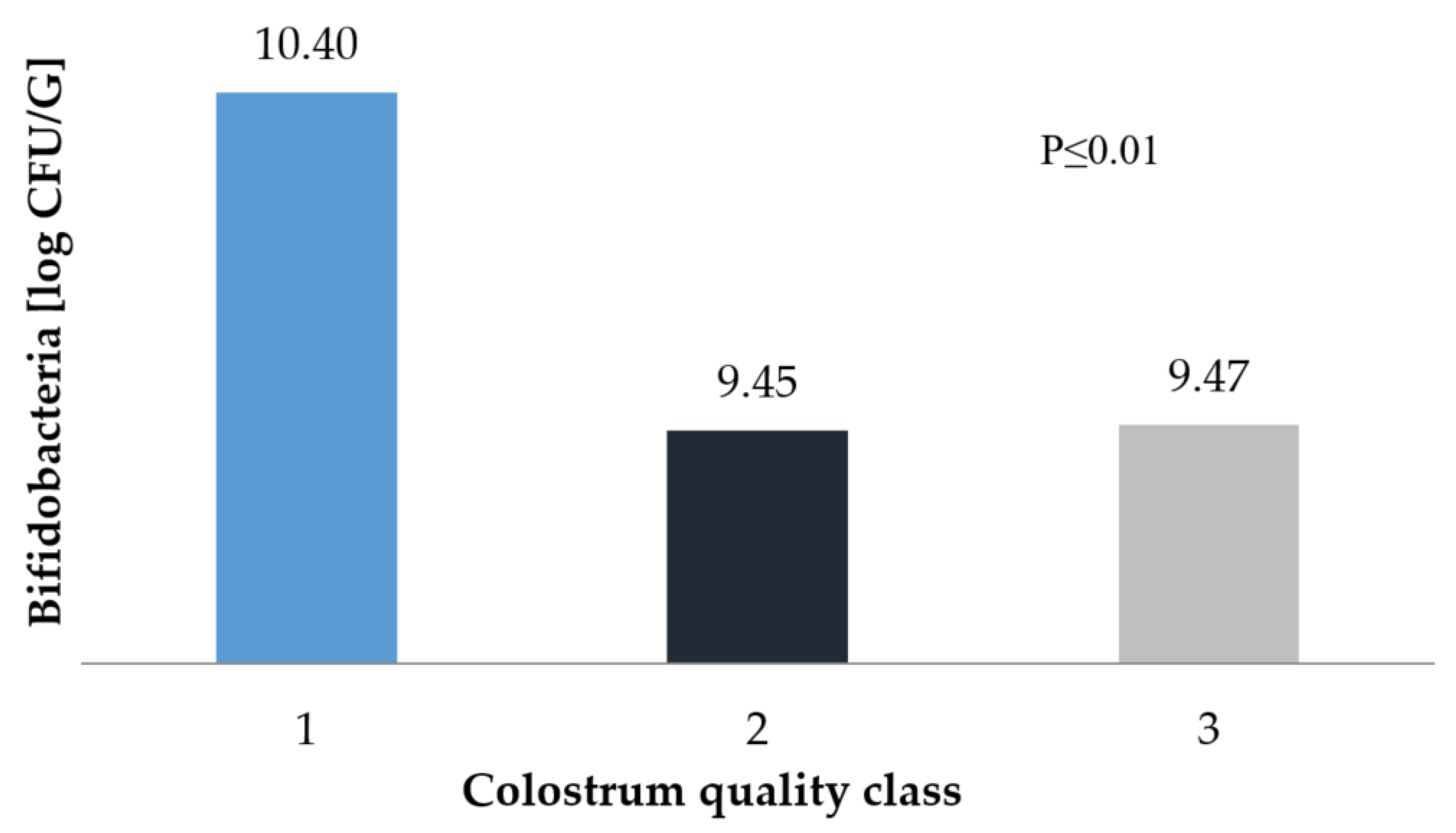
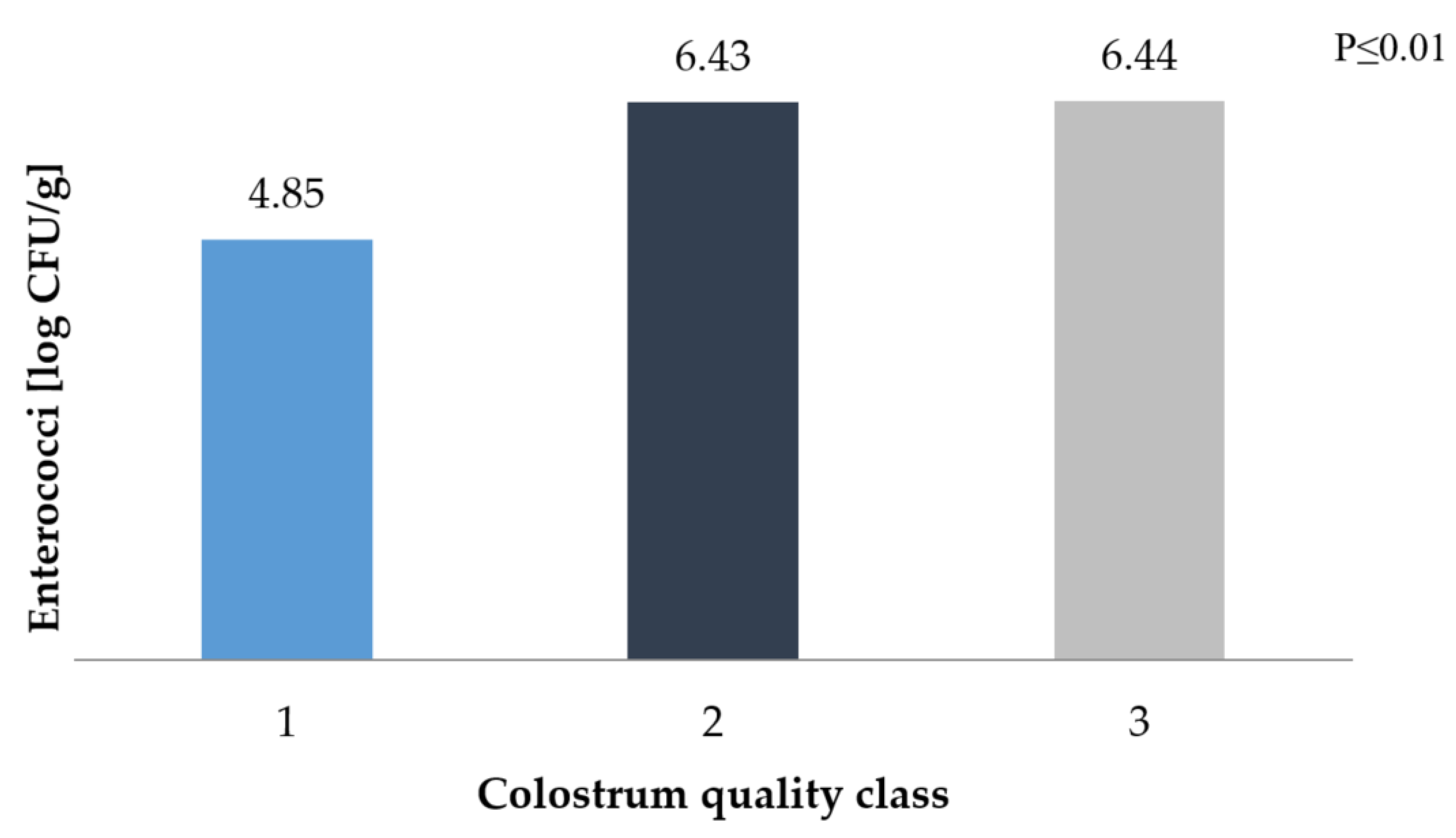
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maunsell, F.P.; Morin, D.E.; Constable, P.D.; Hurley, W.L.; McCoy, G.C.; Kakoma, I.; Isaacson, R.E. Effects of mastitis on the volume and composition of colostrum produced by Holstein cows. J. Dairy Sci. 1998, 81, 1291–1299. [Google Scholar] [CrossRef]
- Godden, S. Colostrum Managment for Dairy Calves. Vet. Clinic. Food Anim. Pract. 2008, 24, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Nardone, A.; Lacetera, N.; Bernabucci, U.; Ronchi, B. Composition of colostrum from dairy heifers exposed to high air temperatures during late pregnancy and the early postpartum period. J. Dairy Sci. 1997, 80, 838–844. [Google Scholar] [CrossRef]
- McGuirk, S.M.; Collins, M. Managing the production; storage and delivery of colostrum. Vet. Clinic. N. Am. Food Anim. Pract. 2004, 20, 593–603. [Google Scholar] [CrossRef]
- Georgiev, I.P. Differences in chemical composition between cow colostrum and milk. Bulg. J. Vet. Med. 2008, 11, 3–12. [Google Scholar]
- Bar, E.; Foris, I.; Miron, M.; Bojita, M.; Mihai, G. Determination of the Content of Immunoglobulin (IgG) and Lactoferrin. Bulletin of University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca. Anim. Sci. Biotechnol. 2009, 66, 1–2. [Google Scholar]
- Artym, J.; Zimecki, M. Rola laktoferryny w prawidłowym rozwoju noworodka the role of lactoferrin in the proper development of newborns. Postepy Hig. Med. Doświad. 2005, 59, 421–432. [Google Scholar]
- Chaterton, D.E.; Smithers, G.; Roupas, P.; Brodkorb, A. Bioactivity of β-lactoglobulin and α-lactalbumin- Technological implications for processing. Int. Dairy J. 2006, 16, 1229–1240. [Google Scholar] [CrossRef]
- Fleming, K.; Thompson-Crispi, K.A.; Hodgins, D.C.; Miglior, F.; Corredig, M.; Mallard, B.A. Short communication: Variation of total immunoglobulin G and β-lactoglobulin concentrations in colostrum and milk from Canadian Holsteins classified as high; average; or low immune responders. J. Dairy Sci. 2016, 99, 2358–2363. [Google Scholar] [CrossRef]
- Wąsowska, E.; Puppel, K. Changes In the content of immunostimulating components of colostrum obtained from dairy cows at different levels of production. J. Sci. Food Agric. 2018, 98, 5062–5068. [Google Scholar] [CrossRef]
- Goel, K. Characterisation and Process Optimisation for the Production of Kharwas-a Colostrum Based Delicacy. Ph.D. Thesis, ICAR–National Dairy Research Institute, Karnal, India, 2015; pp. 6–32. [Google Scholar]
- Caffin, J.P.; Poutrel, B.; Rainard, P. Physiological and pathological factors influencing bovine α-lactalbumin and β-lactoglobulin concentrations in milk. J. Dairy Sci. 1985, 68, 1087–1094. [Google Scholar] [CrossRef]
- Król, J.; Litwińczuk, A.; Zarajczyk, A.; Litwińczuk, Z. Alfa-laktoalbumina i beta-laktoglobulina jako związki biologicznie czynne frakcji białkowej mleka. Med. Wet. 2008, 12, 1375–1378. [Google Scholar]
- Chaneton, L.; Sáez, J.P.; Bussmann, L.E. Antimicrobial activity of bovine β-lactoglobulin against mastitis-causing bacteria. J. Dairy Sci. 2011, 94, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Puppel, K.; Gołębiewski, M.; Grodkowski, G.; Slósarz, J.; Kunowska-Slósarz, M.; Solarczyk, P.; Łukasiewicz, M.; Balcerak, M.; Przysucha, T. Composition and Factors Affecting Quality of Bovine Colostrum: A Review. Animals 2019, 9, 1070. [Google Scholar] [CrossRef]
- Yeoman, C.J.; White, B.A. Gastrointestinal tract microbiota and probiotics in production animals. Ann. Rev. Anim. Biosci. 2014, 2, 469–486. [Google Scholar] [CrossRef]
- Francino, M.P. Early development of the gut microbiota and immune health. Pathogens 2014, 3, 769–790. [Google Scholar] [CrossRef]
- Nowacki, M.R. Cell proliferation in colonic crypts of germ-free and conventional mice-preliminary report. Folia Histochem. Cytobiol. 1993, 31, 77–81. [Google Scholar]
- Sommer, F.; Bäckhed, F. The gut microbiota- masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Nowak, A.; Slizewska, K.; Libudzisz, Z. Probiotyki-historia i mechanizmy działania. Żywność Nauka Technologia Jakość 2010, 17, 5–19. [Google Scholar]
- Gerdts, V.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S.; Griebel, P.J. Fetal immunization by a DNA vaccine delivered orally into the amniotic fluid. Nat. Med. 2000, 6, 929–932. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Griebel, P.J.; Guan, L.L. The gut microbiome and its potential role in the development and function of newborn calf gastrointestinal tract. Front. Vet. Sci. 2015, 2, 36. [Google Scholar] [CrossRef] [PubMed]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, G.; Teixeira, A.G.V.; Foditsch, C.; Bicalho, M.L.; Machado, V.S.; Bicalho, R.C. Fecal microbial diversity in pre-weaned dairy calves as described by pyrosequencing of metagenomic 16S rDNA. Associations of Faecalibacterium species with health and growth. PLoS ONE 2013, 8, e63157. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed]
- Różańska, H.; Lewtak-Piłat, A.; Osek, J. Enterokoki- bakterie o wielu obliczach. Życie Weterynaryjne 2013, 88, 562–564. [Google Scholar]
- Kłossowska, A.; Malinowski, E. Shigatoksyczne szczepy Escherichia coli w infekcjach gruczołu mlekowego u krów. Med. Wet. 2007, 63, 1108–1110. [Google Scholar]
- Frydendahl, K. Prevalence of serogroups and virulence genes in Escherichia coli associated with postweaning diarrhoea and edema disease in pigs and a comparison of diagnostic approaches. Vet. Microbiol. 2002, 85, 169–182. [Google Scholar] [CrossRef]
- Moxley, R.A.; Francis, D.H. Natural and experimental infection with an attaching and effacing strain of Escherichia coli in calves. Infect. Immun. 1986, 53, 339–346. [Google Scholar] [CrossRef]
- Smith, H.W. The development of the flora of the alimentary tract in young animals. J. Pathol. Bacteriol. 1965, 90, 495–513. [Google Scholar] [CrossRef]
- Tannock, G.W. Effect of dietary and environmental stress on the gastrointestinal microbiota. Hum. Intest. Microflora Health Dis. 1983, 517–539. [Google Scholar]
- Franczuk, A.; Jagusztyn-Krynicka, E.K. Rola mikroflory jelit w indukcji choroby Leśniowskiego-Crohna w świetle programu badań Human Microbiome Project. Postępy Mikrobiol. 2012, 51, 257–264. [Google Scholar]
- Hamouda, R.H.; Thannaa, K.H.; Nabih, A.M. Bacteriological and pathological studies on some aerobic and anaerobic bacteria causing diarrhoea in camel calves. Vet. Med. J. Giza 2010, 58, 177–197. [Google Scholar]
- Bashahun, G.M.; Amina, A. Colibacillosis in calves: A review of literature. J. Vet. Med. Sci. 2017, 2, 62–71. [Google Scholar] [CrossRef]
- Abe, F.; Ishibashi, N.; Shimamura, S. Effect of administration of Bifidobacteria and Lactic acid bacteria to newborn calves and piglets. J. Dairy Sci. 1995, 73, 2838–2846. [Google Scholar] [CrossRef]
- Sokol, H.; Seksik, P.; Furet, J.P.; Fermisse, O.; Nion-Larmurier, I.; Beaugerie, L. Low counts of Faecalibacterium prausnitzii in colitis microbiota. Inflamm. Bowel Dis. 2009, 15, 1183–1189. [Google Scholar] [CrossRef]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B. Gut Immune maturation depends on colonization with a host-specific microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef] [PubMed]
- Godden, S.M.; Smolenski, D.J.; Donahue, M.; Oakes, J.M.; Bey, R.; Wells, S.; Sreevatsan, S.; Stabel, J.; Fetrow, J. Heat-treated colostrum and reduced morbidity in preweaned dairy calves: Results of a randomized trial and examination of mechanisms of effectiveness. J. Dairy Sci. 2012, 95, 4029–4040. [Google Scholar] [CrossRef]
- Mohankumar, A.; Murugalatha, N. Characterization and antibacterial activity of bacteriocin producing Lactobacillus isolated from raw cattle milk sample. Int. J. Biol. 2011, 3, 128–143. [Google Scholar] [CrossRef]
- Ziarno, M.; Godlewska, A. Znaczenie i wykorzystani bakterii rodzaju Lactoccocus w mleczarstwie. Med. Wet. 2008, 64, 35–39. [Google Scholar]
- Puppel, K.; Bogusz, E.; Golebiewski, M.; Nalecz-Tarwacka, T.; Kuczynska, B.; Slosarz, J.; Budzinski, A.; Solarczyk, P.; Kunowska-Slosarz, M.; Przysucha, T. Effect of Dairy Cow Crossbreeding on Selected Performance Traits and Quality of Milk in First Generation Crossbreds. J. Food Sci. 2018, 83, 229–236. [Google Scholar] [CrossRef]
- IBM Crop. Released IBM SPSS for Windows, version 23.0; IBM Crop: Armonk, NY, USA, 2020. [Google Scholar]
- Demkowicz, M. Bioaktywne Preparaty z Siary Krów Zachowujące Parametry jej Wartości Biologicznej Oraz ich Przyswajalność u Jagniąt i Cieląt. Praca Doktorska. 2012, pp. 13–32. Available online: http://www.dbc.wroc.pl/dlibra/doccontent?id=20342 (accessed on 29 June 2020).
- Wroński, M.; Sosnowska, W. Relationships between the density and composition of colostrums and the performance of calves. Pol. J. Nat. Sci. 2008, 23, 573–582. [Google Scholar] [CrossRef]
- Pecka, E.; Zachwieja, A.; Zawadzki, W.; Kaszuba, J.; Tumanowicz, J. Wpływ stadium laktacji na wydajność i właściwości fizykochemiczne oraz skład podstawowy mleka krów pierwiastek. Acta Sci. Pol. Med. Vet. 2012, 11, 5–14. [Google Scholar]
- Hyrslova, I.; Krausova, G.; Bartova, J.; Kolesar, L.; Curda, L. Goat and bovine colostrum as a basis for new probiotic functional foods and dietary supplements. J. Microb. Biochem. Technol. 2016, 8, 56–59. [Google Scholar]
- Fonty, G.; Gouet, P.; Jouany, J.P.; Senaud, J. Establishment of the microflora and anaerobic fungi in the rumen of lambs. Microbiology 1987, 133, 1835–1843. [Google Scholar] [CrossRef]
- Minato, H.; Otsuka, M.; Shirasaka, S.; Itabashi, H.; Mitsumori, M. Colonization of microorganisms in the rumen of young calves. J. Gen. App. Microbiol. 1992, 38, 447–456. [Google Scholar] [CrossRef]
- Anderson, K.L.; Nagaraja, T.G.; Morrill, J.L.; Avery, T.B.; Galitzer, S.J.; Boyer, J.E. Ruminal microbial development in conventionally or early-weaned calves. J. Anim. Sci. 1987, 64, 1215–1226. [Google Scholar] [CrossRef]
- Teraguchi, S.; Shin, K.; Ozawa, K.; Nakamura, S.; Fukuwatari, Y.; Tsuyuki, S.; Namihira, H.; Shimamura, S. Bacteriostatic effect of orally administered bovine lactoferrin on proliferation of Clostridium species in the gut of mice fed bovine milk. Appl. Environ. Microbiol. 1995, 61, 501–506. [Google Scholar] [CrossRef]
- Flint, H.J.; Bayer, E.A. Plant cell wall breakdown by anaerobic microorganisms from the mammalian digestive tract. Ann. N. Y. Acad. Sci. 2008, 1125, 280–288. [Google Scholar] [CrossRef]
- Higginbotham, G.E.; Bath, D.L. Evaluation of Lactobacillus fermentation cultures in calf feeding systems. J. Dairy Sci. 1993, 76, 615–620. [Google Scholar] [CrossRef]
- Ellinger, D.K.; Muller, L.D.; Glantz, P.J. Influence of feeding fermented colostrum and Lactobacillus acidophilus on fecal flora of dairy calves. J. Dairy Sci. 1980, 63, 478–482. [Google Scholar] [CrossRef]
- Vlková, E.; Rada, V.; Trojanová, I.; Killer, J.; Šmehilová, M.; Molatová, Z. Occurrence of bifidobacteria in faeces of calves fed milk or a combined diet. Arch. Anim. Nutr. 2008, 62, 359–365. [Google Scholar] [CrossRef]
- Suo, C.; Yin, Y.; Wang, X.; Lou, X.; Song, D.; Wang, X.; Gu, Q. Effects of lactobacillus plantarumZJ316 on pig growth and pork quality. BMC Vet. Res. 2012, 8, 89. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; Godden, S.; Molitor, T.; Ames, T.; Hagman, D. Effects of feeding heat-treated colostrum on passive transfer of immune and nutritional parameters in neonatal dairy calves. J. Dairy Sci. 2007, 90, 5189–5198. [Google Scholar] [CrossRef] [PubMed]
- Gilliland, S.E.; Bruce, B.B.; Bush, L.J.; Stale, T.E. Comparisons of Two Strains of Lactobacillus acidophilus as Dietary Adjuncts for Young Calves. J. Dairy Sci. 1980, 63, 964–972. [Google Scholar] [CrossRef]
- Gregory, N.G.; Jacobson, L.H.; Nagle, T.A.; Muirhead, R.W.; Leroux, G.J. Effect of preslaughter feeding system on weight loss; gut bacteria; and the physico-chemical properties of digesta in cattle. N. Z. J. Agric. Res. 2000, 43, 351–361. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specification | Dry Cow Groups | |
|---|---|---|
| I | III | |
| Assumptions | ||
| Cow weight (kg) | 640 | 675 |
| Pregnancy (days) | 220 | 270 |
| Maintenance requirements | ||
| NEL (Mcal/day) | 9.8 | 11.6 |
| Metabolic protein (g/day) | 463 | 653 |
| Ca (g/day) | 11 | 16.5 |
| P (g/day) | 12 | 16.3 |
| K (g/day) | 54 | 55 |
| Foetus requirements | ||
| NEL (Mcal/day) | 2.9 | 3 |
| Metabolic protein (g/day) | 239 | 245 |
| Ca (g/day) | 4 | 5 |
| P (g/day) | 3 | 4 |
| K (g/day) | 1 | 2 |
| Total requirements | ||
| NEL (Mcal/day) | 12.8 | 14.4 |
| Metabolic protein (g/day) | 700 | 901 |
| Ca (g/day) | 15 | 21.5 |
| P (g/day) | 15 | 20.3 |
| K (g/day) | 55 | 57 |
| Colostrum Class | Density [g/cm3] | Casein [%] | Protein [%] | Fat [%] |
|---|---|---|---|---|
| 1st | >1.070 | 9.60 aB | 28.42 AB | 7.29 AB |
| 2nd | 1.057–1.070 | 8.53 ac | 22.39 AC | 6.64 Ac |
| 3rd | 1.044–1.056 | 7.39 Bc | 18.22 BC | 5.12 Bc |
| Colostrum Class | LF [g/L] | α-LA [g/L] | β-LG [g/L] | IgG [g/L] |
|---|---|---|---|---|
| 1st | 4.40 AB | 7.52 AB | 8.04 AB | 92.00 AB |
| 2nd | 3.03 Ac | 4.55 AC | 6.16 AC | 74.60 AC |
| 3rd | 2.53 Bc | 3.47 BC | 4.8 BC | 53.50 BC |
| Colostrum Class | Weight [kg] | Daily Weight Gain [kg/day] | |
|---|---|---|---|
| Day 0 | Day 7 | ||
| 1st | 40.17 | 43.45 | 0.468 AB |
| 2nd | 41.67 | 43.23 | 0.222 A |
| 3rd | 42.17 | 43.50 | 0.190 B |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puppel, K.; Gołębiewski, M.; Konopka, K.; Kunowska-Slósarz, M.; Slósarz, J.; Grodkowski, G.; Przysucha, T.; Balcerak, M.; Madras-Majewska, B.; Sakowski, T. Relationship between the Quality of Colostrum and the Formation of Microflora in the Digestive Tract of Calves. Animals 2020, 10, 1293. https://doi.org/10.3390/ani10081293
Puppel K, Gołębiewski M, Konopka K, Kunowska-Slósarz M, Slósarz J, Grodkowski G, Przysucha T, Balcerak M, Madras-Majewska B, Sakowski T. Relationship between the Quality of Colostrum and the Formation of Microflora in the Digestive Tract of Calves. Animals. 2020; 10(8):1293. https://doi.org/10.3390/ani10081293
Chicago/Turabian StylePuppel, Kamila, Marcin Gołębiewski, Katarzyna Konopka, Małgorzata Kunowska-Slósarz, Jan Slósarz, Grzegorz Grodkowski, Tomasz Przysucha, Marek Balcerak, Beata Madras-Majewska, and Tomasz Sakowski. 2020. "Relationship between the Quality of Colostrum and the Formation of Microflora in the Digestive Tract of Calves" Animals 10, no. 8: 1293. https://doi.org/10.3390/ani10081293
APA StylePuppel, K., Gołębiewski, M., Konopka, K., Kunowska-Slósarz, M., Slósarz, J., Grodkowski, G., Przysucha, T., Balcerak, M., Madras-Majewska, B., & Sakowski, T. (2020). Relationship between the Quality of Colostrum and the Formation of Microflora in the Digestive Tract of Calves. Animals, 10(8), 1293. https://doi.org/10.3390/ani10081293