The Reduction of Methane Production in the In Vitro Ruminal Fermentation of Different Substrates is Linked with the Chemical Composition of the Essential Oil

 ,
,  , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Essential Oils
2.3. Substrate
2.4. In Vitro Incubation
2.5. Chemical Analyses, Determinations, and Calculations
2.5.1. pH
2.5.2. Gas Production and Methane
2.5.3. Substrate Digestibility
2.5.4. Values Relative to the Control
2.6. Statistical Analysis
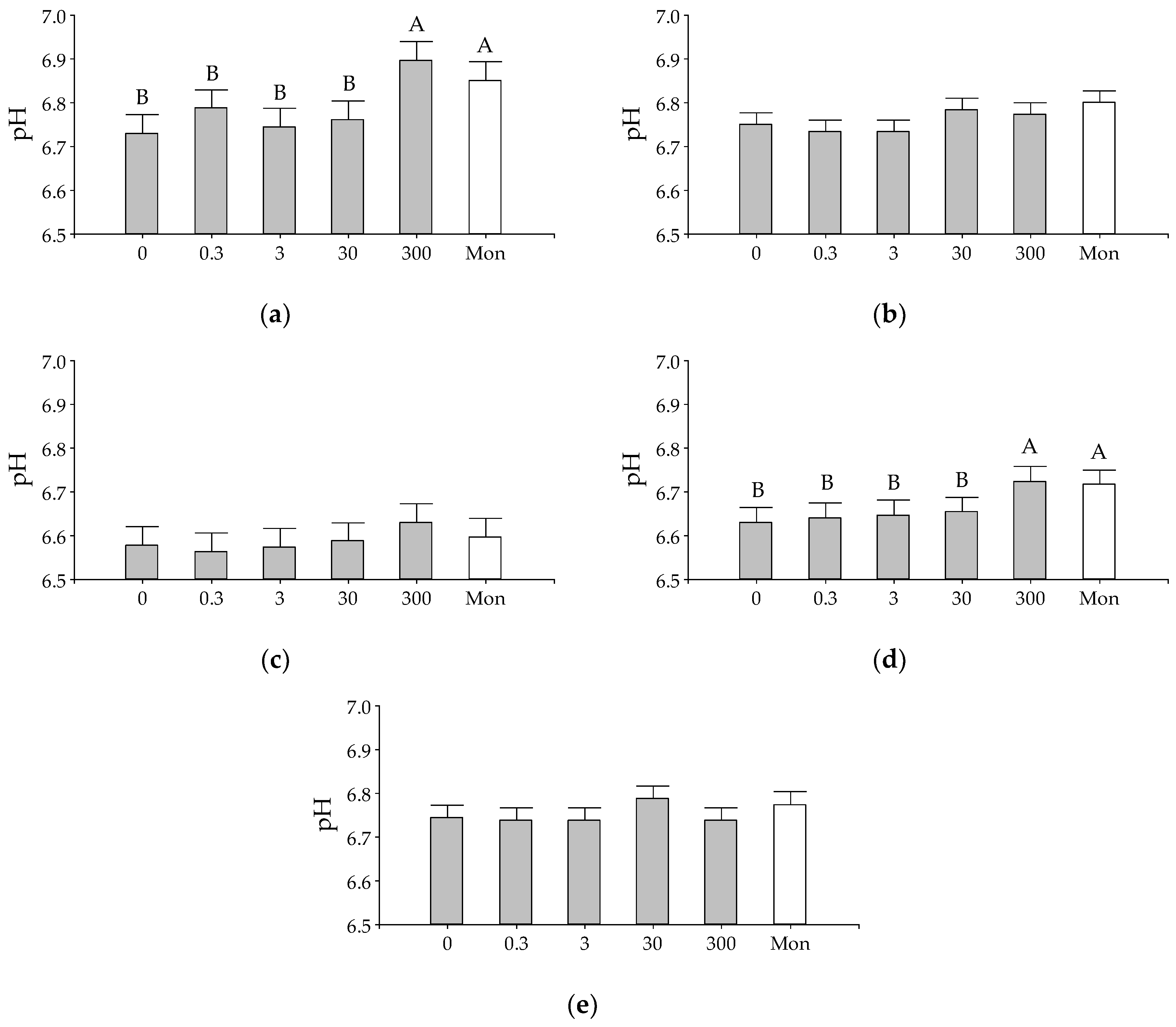
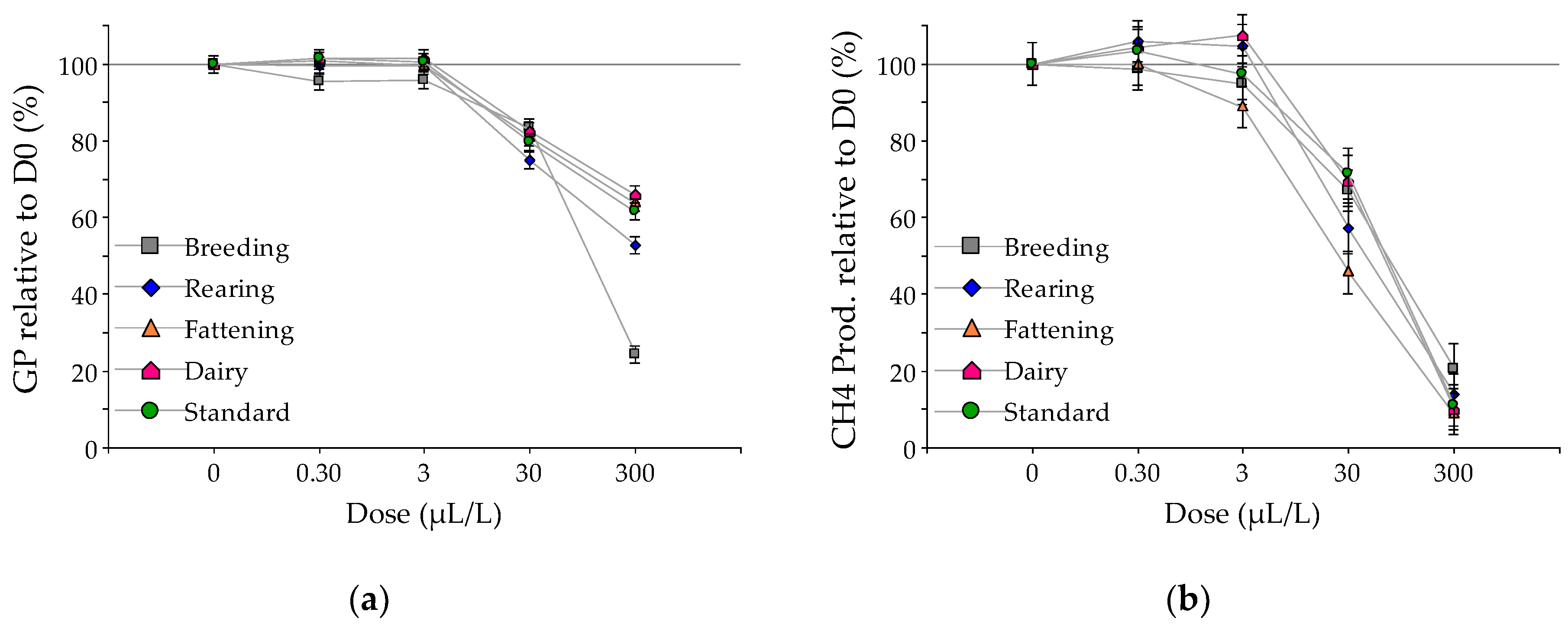
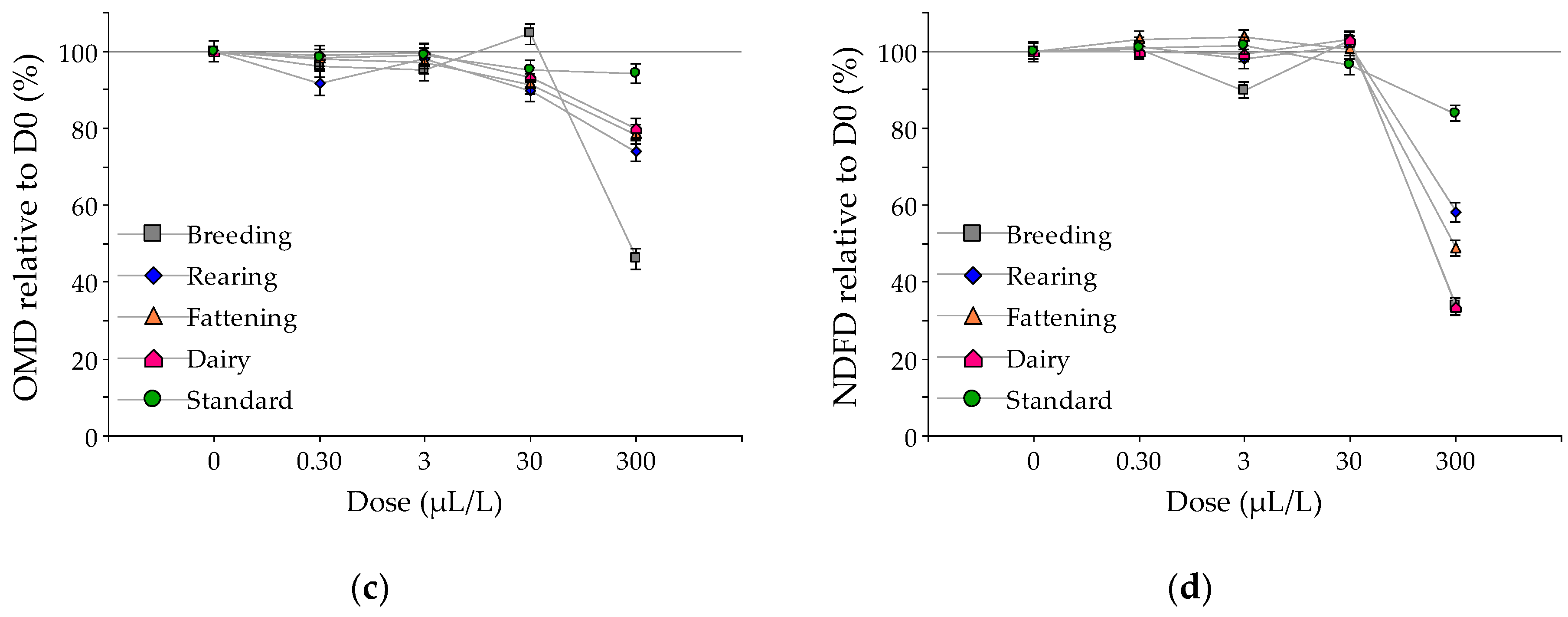
3. Results
Effect of EO on Fermentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Ganadería Mundial 2011-La Ganadería En La Seguridad Alimentaria; FAO: Rome, Italy, 2012. [Google Scholar]
- Moumen, A.; Azizi, G.; Chekroun, K.B.; Mourad, B. The Effects of Livestock Methane Emission on the Global Warming: A Review. Int. J. Glob. Warm. 2016, 9, 229–253. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1535. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane Emissions From Cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen Methanogens and Mitigation of Methane Emission by Anti-Methanogenic Compounds and Substances. J. Anim. Sci. Biotechnol. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Appuhamy, J.A.D.R.N.; Strathe, A.B.; Jayasundara, S.; Wagner-Riddle, C.; Dijkstra, J.; France, J.; Kebreab, E. Anti-Methanogenic Effects of Monensin in Dairy and Beef Cattle: A Meta-Analysis. J. Dairy Sci. 2013, 96, 5161–5173. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fèvre, E.M.; Gilbert, M.; Grace, D.; Hay, S.I.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic Resistance: Mitigation Opportunities in Livestock Sector Development. Animal 2017, 11, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Acamovic, T.; Brooker, J.D. Biochemistry of Plant Secondary Metabolites and Their Effects in Animals. Proc. Nutr. Soc. 2005, 64, 403–412. [Google Scholar] [CrossRef]
- Castillejos, L.; Calsamiglia, S.; Ferret, A.; Losa, R. Effects of a Specific Blend of Essential Oil Compounds and the Type of Diet on Rumen Microbial Fermentation and Nutrient Flow from a Continuous Culture System. Anim. Feed Sci. Technol. 2005, 119, 29–41. [Google Scholar] [CrossRef]
- Benchaar, C.; Greathead, H. Essential Oils and Opportunities to Mitigate Enteric Methane Emissions from Ruminants. Anim. Feed Sci. Technol. 2011, 166–167, 338–355. [Google Scholar] [CrossRef]
- Garcia, F.; Vercoe, P.E.; Martínez, M.J.; Durmic, Z.; Brunetti, M.A.; Moreno, M.V.; Colombatto, D.; Lucini, E.; Martínez Ferrer, J. Essential Oils from Lippia Turbinata and Tagetes Minuta Persistently Reduce in Vitro Ruminal Methane Production in a Continuous-Culture System. Anim. Prod. Sci. 2018, 59, 709–720. [Google Scholar] [CrossRef]
- Bodas, R.; Prieto, N.; García-González, R.; Andrés, S.; Giráldez, F.J.; López, S. Manipulation of Rumen Fermentation and Methane Production with Plant Secondary Metabolites. Anim. Feed Sci. Technol. 2012, 176, 78–93. [Google Scholar] [CrossRef]
- Klevenhusen, F.; Muro-Reyes, A.; Khiaosa-ard, R.; Metzler-Zebeli, B.U.; Zebeli, Q. A Meta-Analysis of Effects of Chemical Composition of Incubated Diet and Bioactive Compounds on in Vitro Ruminal Fermentation. Anim. Feed Sci. Technol. 2012, 176, 61–69. [Google Scholar] [CrossRef]
- Molero, R.; Ibars, M.; Calsamiglia, S.; Ferret, A.; Losa, R. Effects of a Specific Blend of Essential Oil Compounds on Dry Matter and Crude Protein Degradability in Heifers Fed Diets with Different Forage to Concentrate Ratios. Anim. Feed Sci. Technol. 2004, 114, 91–104. [Google Scholar] [CrossRef]
- Castillejos, L. Modificación de La Fermentación Microbiana Ruminal Mediante Compuestos de Aceites Esenciales. Ph.D. Thesis, Universitat Autònoma de Barcelona, Bellaterra, Spain, 2005. [Google Scholar]
- Mateos, I.; Ranilla, M.J.; Tejido, M.L.; Saro, C.; Kamel, C.; Carro, M.D. The Influence of Diet Type (Dairy versus Intensive Fattening) on the Effectiveness of Garlic Oil and Cinnamaldehyde to Manipulate in Vitro Ruminal Fermentation and Methane Production. Anim. Prod. Sci. 2013, 53, 299–307. [Google Scholar] [CrossRef]
- Pérez-Zamora, C.M.; Torres, C.; Aguado, M.I.; Bela, A.J.; Nuñez, M.B.; Bregni, C. Antibacterial Activity of Essential Oils of Aloysia Polystachya and Lippia Turbinata (Verbenaceae). Bol. Latinoam. Del Caribe Plantas Med. Aromát. 2016, 15, 199–205. [Google Scholar]
- García, C.C.; Talarico, L.; Almeida, N.; Colombres, S.; Duschatzky, C.; Damonte, E.B. Virucidal Activity of Essential Oils from Aromatic Plants of San Luis, Argentina. Phyther. Res. 2003, 17, 1073–1085. [Google Scholar] [CrossRef]
- Dellacasa, A.D.; Bailac, P.N.; Ponzi, M.I.; Ruffinengo, S.R.; Eguaras, M.J. In Vitro Activity of Essential Oils from San Luis-Argentina against Ascosphaera Apis. J. Essent. Oil Res. 2003, 15, 282–285. [Google Scholar] [CrossRef]
- Terblanché, F.C.; Kornelius, G. Essential Oil Constituents of the Genus Lippia (Verbenaceae)—A Literature Review. J. Essent. Oil Res. 1996, 8, 471–485. [Google Scholar] [CrossRef]
- Héthélyi, E.; Dános, B.; Tétényi, P. GC-MS Analysis of the Essential Oils of Four Tagetes Species and the Anti-Microbial Activity of Tagetes Minuta. Flavour Fragr. J. 1986, 1, 169–173. [Google Scholar] [CrossRef]
- Garcia, M.V.; Matias, J.; Cavalcante Barros, J.C.; Pries de Lima, D.; da Silva Lopes, R.; Andreotti, R. Chemical Identification of Tagetes Minuta Linnaeus (Asteraceae) Essential Oil and Its Acaricidal Effect on Ticks. Rev. Bras. Parasitol. Vet. 2012, 21, 405–411. [Google Scholar] [CrossRef]
- Senatore, F.; Napolitano, F.; Mohamed, M.A.-H.; Harris, P.J.C.; Mnkeni, P.N.S.; Henderson, J. Antibacterial Activity of Tagetes Minuta, L. (Asteraceae) Essential Oil with Different Chemical Composition. Flavour Fragr. J. 2004, 19, 574–578. [Google Scholar] [CrossRef]
- Chamorro, E.R.; Ballerini, G.; Sequeira, A.F.; Velasco, G.A.; Zalazar, M.F. Chemical Composition of Essential Oil from Tagetes Minuta, L. Leaves and Flowers. J. Argent. Chem. Soc. 2008, 96, 80–86. [Google Scholar]
- Burt, S. Essential Oils: Their Antibacterial Properties and Potential Applications in Foods-A Review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Kahvand, M.; Malecky, M. Dose-Response Effects of Sage (Salvia Officinalis) and Yarrow (Achillea Millefolium) Essential Oils on Rumen Fermentation in Vitro. Ann. Anim. Sci. 2018, 18, 125–142. [Google Scholar] [CrossRef]
- FASS. Guide for the Care and Use of Agricultural Animals in Research and Teaching, 3rd ed.; FASS Inc.: Champaign, IL, USA, 2010; p. 177. Available online: http://www.fass.org (accessed on 7 January 2016).
- Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S.; McAllan, A.B.; France, J. A Simple Gas Production Method Using a Pressure Transducer to Determine the Fermentation Kinetics of Ruminant Feeds. Anim. Feed Sci. Technol. 1994, 48, 185–197. [Google Scholar] [CrossRef]
- Araujo, R.C.; Pires, A.V.; Mourão, G.B.; Abdalla, A.L.; Sallam, S.M.A. Use of Blanks to Determine in Vitro Net Gas and Methane Production When Using Rumen Fermentation Modifiers. Anim. Feed Sci. Technol. 2011, 166–167, 155–162. [Google Scholar] [CrossRef]
- AOCS. Official Methods and Recommended Practices of the American Oil Chemists’ Society; American Oil Chemists’ Society: Urbana, IL, USA, 1998. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Tilley, J.M.A.; Terry, R.A. A Two-Stage Technique for the in Vitro Digestion of Forage Crops. Grass Forage Sci. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat versión 2019. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Available online: http://www.infostat.com.ar (accessed on 25 May 2019).
- Di Rienzo, J.A.; Guzmán, A.W.; Casanoves, F. A Multiple-Comparisons Method Based on the Distribution of the Root Node Distance of a Binary Tree. J. Agric. Biol. Environ. Stat. 2002, 7, 129–142. [Google Scholar] [CrossRef]
- Cardozo, P.W.; Calsamiglia, S.; Ferret, A.; Kamel, C. Screening for the Effects of Natural Plant Extracts at Different PH on in Vitro Rumen Microbial Fermentation of a High-Concentrate Diet for Beef Cattle. J. Anim. Sci. 2005, 83, 2572–2579. [Google Scholar] [CrossRef]
- Benchaar, C.; Calsamiglia, S.; Chaves, A.V.; Fraser, G.R.; Colombatto, D.; McAllister, T.A.; Beauchemin, K.A. A Review of Plant-Derived Essential Oils in Ruminant Nutrition and Production. Anim. Feed Sci. Technol. 2008, 145, 209–228. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial Agents from Plants: Antibacterial Activity of Plant Volatile Oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Castillejos, L.; Calsamiglia, S.; Ferret, A. Effect of Essential Oil Active Compounds on Rumen Microbial Fermentation and Nutrient Flow in in Vitro Systems. J. Dairy Sci. 2006, 89, 2649–2658. [Google Scholar] [CrossRef]
- Kouazounde, J.B.; Jin, L.; Assogba, F.M.; Ayedoun, M.A.; Wang, Y.; Beauchemin, K.A.; McAllister, T.A.; Gbenou, J.D. Effects of Essential Oils from Medicinal Plants Acclimated to Benin on in Vitro Ruminal Fermentation of Andropogon Gayanus Grass. J. Sci. Food Agric. 2015, 95, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Cattani, M.; Maccarana, L.; Rossi, G.; Tagliapietra, F.; Schiavon, S.; Bailoni, L. Dose-Response and Inclusion Effects of Pure Natural Extracts and Synthetic Compounds on in Vitro Methane Production. Anim. Feed Sci. Technol. 2016, 218, 100–109. [Google Scholar] [CrossRef]
- Joch, M.; Cermak, L.; Hakl, J.; Hucko, B.; Duskova, D.; Marounek, M. In Vitro Screening of Essential Oil Active Compounds for Manipulation of Rumen Fermentation and Methane Mitigation. Asian-Australas J. Anim. Sci. 2016, 29, 952–959. [Google Scholar] [CrossRef]
- Garcia, F.; Martínez Ferrer, J.; Durmic, Z.; Vercoe, P.E. Effect of Ethanol as Carrier of Essential Oils on in Vitro Methane Production. In Proceedings of the XXIV Reunión de la Asociación Latinoamericana de Producción Animal (ALPA), Puerto Varas, Chile, 9–13 November 2015; p. 518. [Google Scholar]
- Joch, M.; Kudrna, V.; Hakl, J.; Božik, M.; Homolka, P.; Illek, J.; Tyrolová, Y.; Výborná, A. In Vitro and in Vivo Potential of a Blend of Essential Oil Compounds to Improve Rumen Fermentation and Performance of Dairy Cows. Anim. Feed Sci. Technol. 2019, 251, 176–186. [Google Scholar] [CrossRef]
- Patra, A.K.; Yu, Z. Effects of Essential Oils on Methane Production and Fermentation by, and Abundance and Diversity of, Rumen Microbial Populations. Appl. Environ. Microbiol. 2012, 78, 4271–4280. [Google Scholar] [CrossRef]
- Patra, A.K.; Yu, Z. Essential Oils Affect Populations of Some Rumen Bacteria in Vitro as Revealed by Microarray (RumenBactArray) Analysis. Front. Microbiol. 2015, 6, 297. [Google Scholar] [CrossRef]
- Cobellis, G.; Trabalza-Marinucci, M.; Marcotullio, M.C.; Yu, Z. Evaluation of Different Essential Oils in Modulating Methane and Ammonia Production, Rumen Fermentation, and Rumen Bacteria in Vitro. Anim. Feed Sci. Technol. 2016, 215, 25–36. [Google Scholar] [CrossRef]
- Cobellis, G.; Trabalza-Marinucci, M.; Yu, Z. Critical Evaluation of Essential Oils as Rumen Modifiers in Ruminant Nutrition: A Review. Sci. Total Environ. 2016, 545–546, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Ruiz, D.R.; Bannink, A.; Dijkstra, J.; Kebreab, E.; Morgavi, D.P.; O’Kiely, P.; Reynolds, C.K.; Schwarm, A.; Shingfield, K.J.; Yu, Z.; et al. Design, Implementation and Interpretation of n Vitro Batch Culture Experiments to Assess Enteric Methane Mitigation in Ruminants—A Review. Anim. Feed Sci. Technol. 2016, 216, 1–18. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Standard | Breeding | Rearing | Fattening | Dairy |
|---|---|---|---|---|---|
| Ingredients (%, DM basis) | |||||
| Alfalfa hay | 80 | - | 15 | 15 | 15 |
| Corn grain | 20 | - | - | 80 | - |
| Wet corn grain | - | - | - | - | 28 |
| Corn silage | - | - | - | - | 46 |
| Sorghum silage | - | - | 70 | - | - |
| Soybean expeller | - | - | 15 | 5 | 10 |
| Panicum maximum | - | 100 | - | - | - |
| Urea | - | - | - | - | 1 |
| Chemical composition (g/kg DM) | |||||
| Organic matter | 913 | 897 | 910 | 968 | 946 |
| Crude protein | 178 | 40 | 167 | 124 | 164 |
| Neutral detergent fiber | 373 | 770 | 472 | 175 | 265 |
| Acid detergent fiber | 239 | 462 | 281 | 71 | 142 |
| In vitro dry matter digestibility | 739 | 554 | 681 | 877 | 821 |
| Substrate | Additive 1 | Gas Production mL/g OM | Methane Production mL/g OM | OMD 2 g/g | NDFD 3 g/g |
|---|---|---|---|---|---|
| Standard | 0 | 289 aB | 34.3 aA | 0.662 aC | 0.485 aC |
| 0.3 | 293 aB | 35.5 aA | 0.648 aB | 0.489 aC | |
| 3 | 290 aB | 33.1 aA | 0.655 aB | 0.492 aB | |
| 30 | 230 bB | 24.3 bA | 0.631 aB | 0.470 bC | |
| 300 | 178 cB | 3.7 cA | 0.624 aA | 0.407 cA | |
| Monensin | 223 bB | 20.2 bA | 0.636 aB | 0.456 bA | |
| Breeding | 0 | 199 aD | 25.2 aB | 0.420 aE | 0.441 aD |
| 0.3 | 190 aD | 24.8 aC | 0.404 aD | 0.443 aD | |
| 3 | 191 aD | 23.4 aB | 0.398 aD | 0.396 aD | |
| 30 | 167 bC | 16.8 bA | 0.439 aD | 0.454 aC | |
| 300 | 49 dD | 5.2 cA | 0.193 bC | 0.149 bB | |
| Monensin | 74 cD | 7.3 cC | 0.242 bD | 0.180 bC | |
| Rearing | 0 | 243 aC | 29.1 aB | 0.581 aD | 0.421 aD |
| 0.3 | 242 aC | 30.7 aB | 0.532 aC | 0.426 aD | |
| 3 | 243 aC | 30.5 aA | 0.569 aC | 0.412 aD | |
| 30 | 182 bC | 16.3 bA | 0.521 aC | 0.427 aD | |
| 300 | 128 dC | 3.9 cA | 0.430 bB | 0.245 cB | |
| Monensin | 169 cC | 15.3 bB | 0.519 aC | 0.347 bB | |
| Fattening | 0 | 332 aA | 37.8 aA | 0.891 aA | 0.709 aA |
| 0.3 | 334 aA | 37.6 aA | 0.873 aA | 0.731 aA | |
| 3 | 330 aA | 33.7 aA | 0.864 aA | 0.735 aA | |
| 30 | 269 bA | 17.2 bA | 0.814 bA | 0.713 aA | |
| 300 | 211 cA | 3.3 cA | 0.698 cA | 0.346 cA | |
| Monensin | 259 bA | 17.0 bB | 0.813 bA | 0.463 bA | |
| Dairy | 0 | 303 aB | 34.4 aA | 0.811 aB | 0.592 aB |
| 0.3 | 307 aB | 35.5 aA | 0.801 aA | 0.591 aB | |
| 3 | 308 aB | 36.7 aA | 0.806 aA | 0.588 aB | |
| 30 | 250 bA | 22.9 bA | 0.756 bA | 0.610 aB | |
| 300 | 200 cA | 3.4 dA | 0.648 cA | 0.197 cB | |
| Monensin | 209 cB | 14.5 cB | 0.709 bB | 0.278 bB | |
| Standard error | 6.9 | 1.77 | 0.0281 | 0.0186 | |
| p-value | Substrate (S) | <0.0001 | 0.0001 | <0.0001 | <0.0001 |
| Additive (A) | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| S × A | <0.0001 | 0.0008 | 0.0002 | <0.0001 | |
| Substrate | Additive 1 | Gas Production mL/g OM | Methane Production mL/g OM | OMD 2 g/g | NDFD 3 g/g |
|---|---|---|---|---|---|
| Standard | 0 | 287 aB | 33.5 aA | 0.680 aB | 0.430 aB |
| 0.3 | 284 aB | 34.4 aB | 0.670 aB | 0.422 aC | |
| 3 | 286 aB | 34.0 aA | 0.670 aC | 0.420 aC | |
| 30 | 244 bB | 27.9 bA | 0.640 aB | 0.414 aB | |
| 300 | 149 cB | 2.2 dA | 0.557 bB | 0.297 cB | |
| Monensin | 211 bB | 18.4 cA | 0.633 aB | 0.351 bB | |
| Breeding | 0 | 193 aC | 25.9 aB | 0.344 aC | 0.365 aB |
| 0.3 | 189 aC | 23.5 aD | 0.372 aC | 0.354 aC | |
| 3 | 195 aC | 24.1 aB | 0.353 aD | 0.366 aC | |
| 30 | 162 bC | 17.1 bB | 0.337 aD | 0.364 aB | |
| 300 | 36 dC | 02.3 dA | 0.128 bC | 0.104 bB | |
| Monensin | 59 cD | 7.1 cB | 0.138 bD | 0.103 bC | |
| Rearing | 0 | 251 aB | 28.2 aB | 0.617 aB | 0.497 aB |
| 0.3 | 247 aB | 29.1 aC | 0.615 aB | 0.465 aC | |
| 3 | 246 aB | 28.8 aB | 0.606 aC | 0.456 aC | |
| 30 | 215 bB | 23.8 bB | 0.562 aC | 0.452 aB | |
| 300 | 116 dB | 2.2 dA | 0.462 bB | 0.264 bB | |
| Monensin | 167 cC | 14.4 cA | 0.545 aC | 0.292 bB | |
| Fattening | 0 | 347 aA | 35.8 aA | 0.873 aA | 0.818 aA |
| 0.3 | 347 aA | 36.5 aB | 0.839 aA | 0.811 aA | |
| 3 | 352 aA | 37.2 aA | 0.862 aA | 0.825 aA | |
| 30 | 319 aA | 33.3 aA | 0.808 aA | 0.794 aA | |
| 300 | 237 cA | 2.2 cA | 0.757 bA | 0.608 bA | |
| Monensin | 285 bA | 18.7 bA | 0.828 aA | 0.676 bA | |
| Dairy | 0 | 385 aA | 39.3 aA | 0.710 aB | 0.542 aB |
| 0.3 | 386 aA | 41.8 aA | 0.753 aA | 0.545 aB | |
| 3 | 381 aA | 36.8 aA | 0.750 aB | 0.537 aB | |
| 30 | 331 bA | 29.4 bA | 0.715 aB | 0.486 aB | |
| 300 | 225 dA | 2.7 dA | 0.593 bB | 0.234 bB | |
| Monensin | 276 cA | 16.4 cA | 0.650 bB | 0.267 bB | |
| Standard error | 15.4 | 2.65 | 0.0279 | 0.0420 | |
| p-value | Substrate (S) | <0.0001 | 0.0001 | <0.0001 | 0.0001 |
| Additive (A) | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| S × A | 0.0338 | 0.0012 | 0.0002 | 0.0114 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, F.; Colombatto, D.; Brunetti, M.A.; Martínez, M.J.; Moreno, M.V.; Scorcione Turcato, M.C.; Lucini, E.; Frossasco, G.; Martínez Ferrer, J. The Reduction of Methane Production in the In Vitro Ruminal Fermentation of Different Substrates is Linked with the Chemical Composition of the Essential Oil. Animals 2020, 10, 786. https://doi.org/10.3390/ani10050786
Garcia F, Colombatto D, Brunetti MA, Martínez MJ, Moreno MV, Scorcione Turcato MC, Lucini E, Frossasco G, Martínez Ferrer J. The Reduction of Methane Production in the In Vitro Ruminal Fermentation of Different Substrates is Linked with the Chemical Composition of the Essential Oil. Animals. 2020; 10(5):786. https://doi.org/10.3390/ani10050786
Chicago/Turabian StyleGarcia, Florencia, Darío Colombatto, M. Alejandra Brunetti, M. José Martínez, M. Valeria Moreno, M. Carolina Scorcione Turcato, Enrique Lucini, Georgina Frossasco, and Jorge Martínez Ferrer. 2020. "The Reduction of Methane Production in the In Vitro Ruminal Fermentation of Different Substrates is Linked with the Chemical Composition of the Essential Oil" Animals 10, no. 5: 786. https://doi.org/10.3390/ani10050786
APA StyleGarcia, F., Colombatto, D., Brunetti, M. A., Martínez, M. J., Moreno, M. V., Scorcione Turcato, M. C., Lucini, E., Frossasco, G., & Martínez Ferrer, J. (2020). The Reduction of Methane Production in the In Vitro Ruminal Fermentation of Different Substrates is Linked with the Chemical Composition of the Essential Oil. Animals, 10(5), 786. https://doi.org/10.3390/ani10050786