Genomic Scan for Selection Signature Reveals Fat Deposition in Chinese Indigenous Sheep with Extreme Tail Types

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethnic Statement
2.2. Genotype Data
2.3. Population Structure Analysis
2.4. Selection Signature Detection Using hapFLK
2.5. Gene Annotation and Enrichment Analysis
3. Results
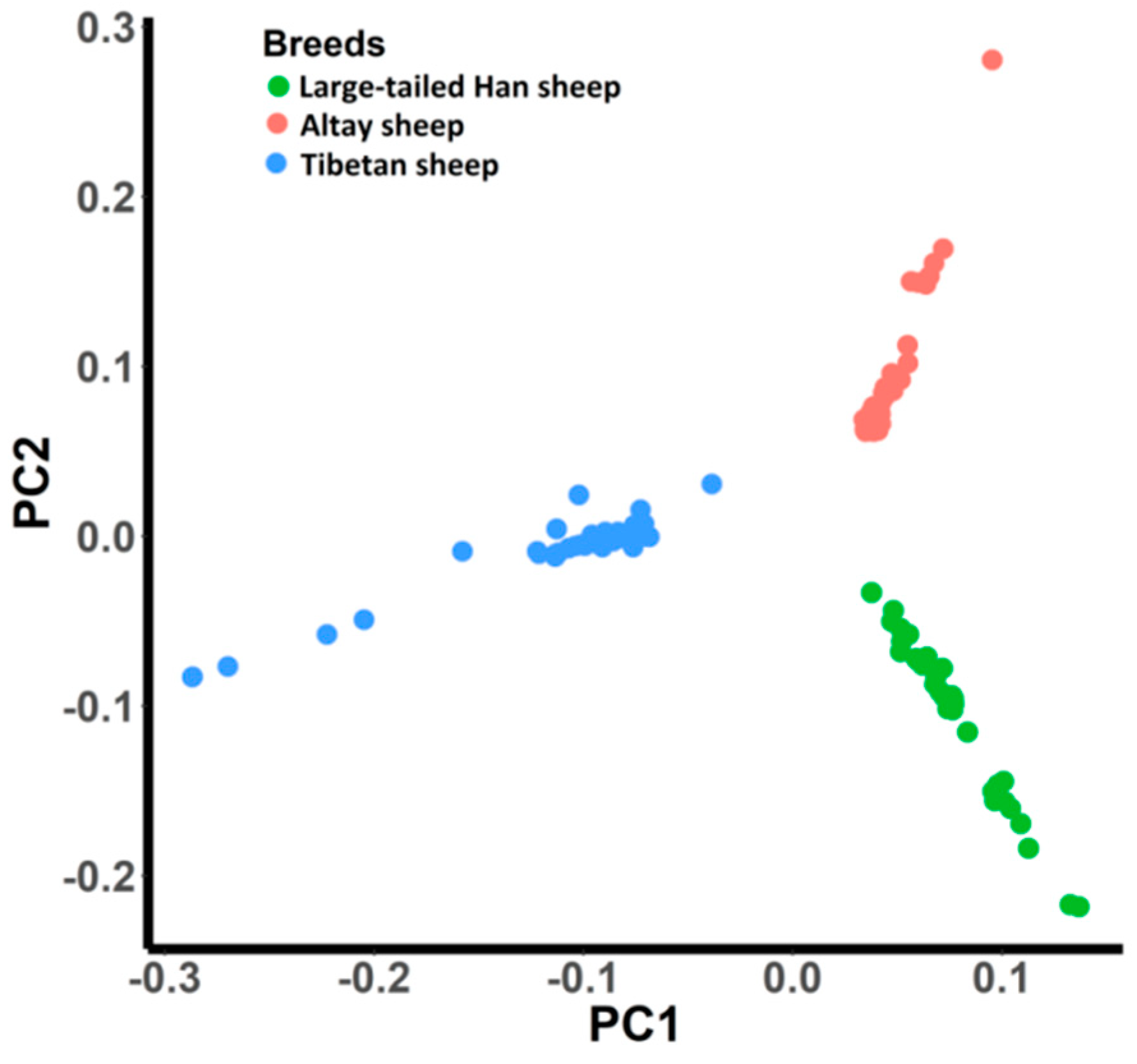
3.1. Population Genetic Structure
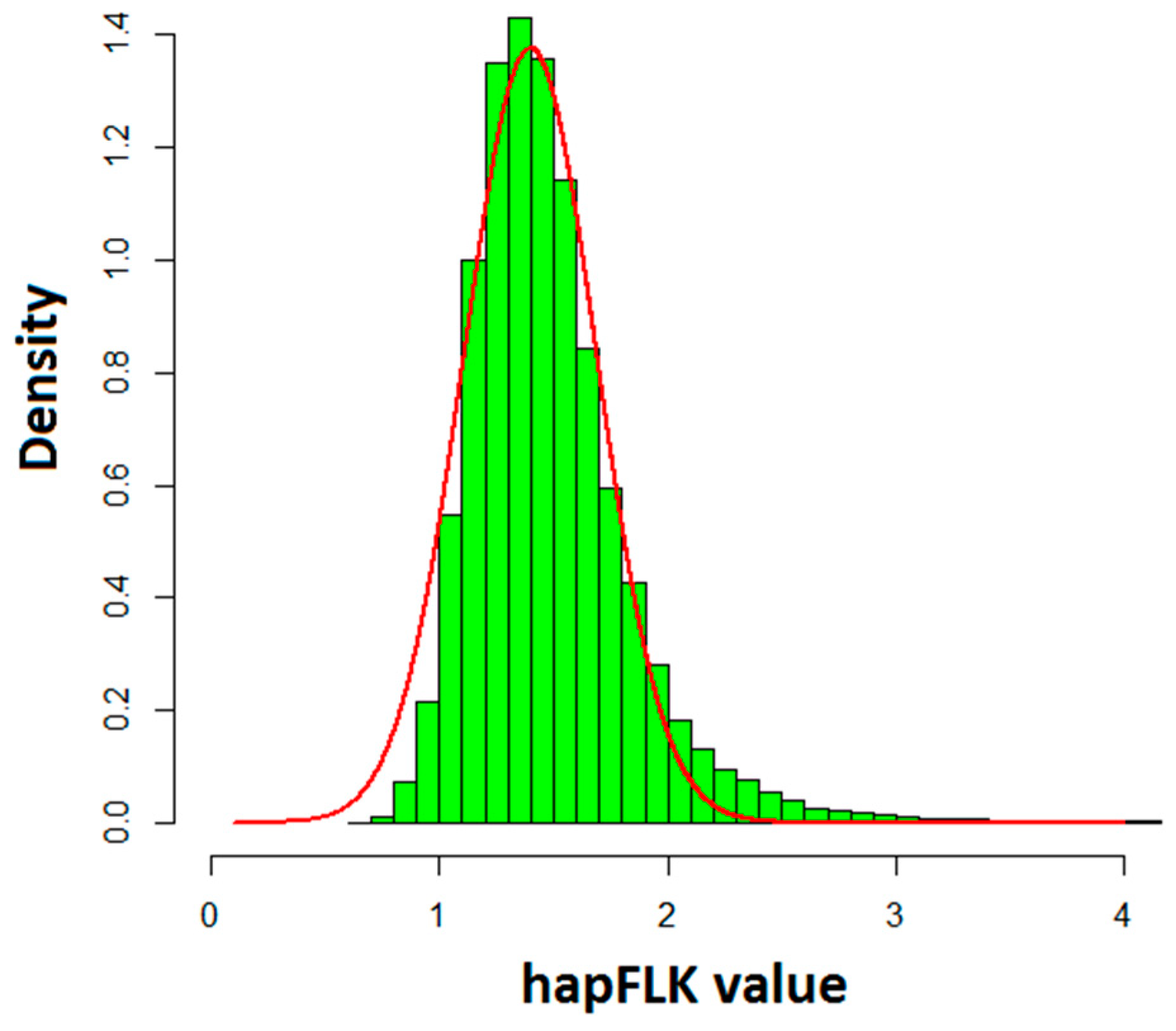
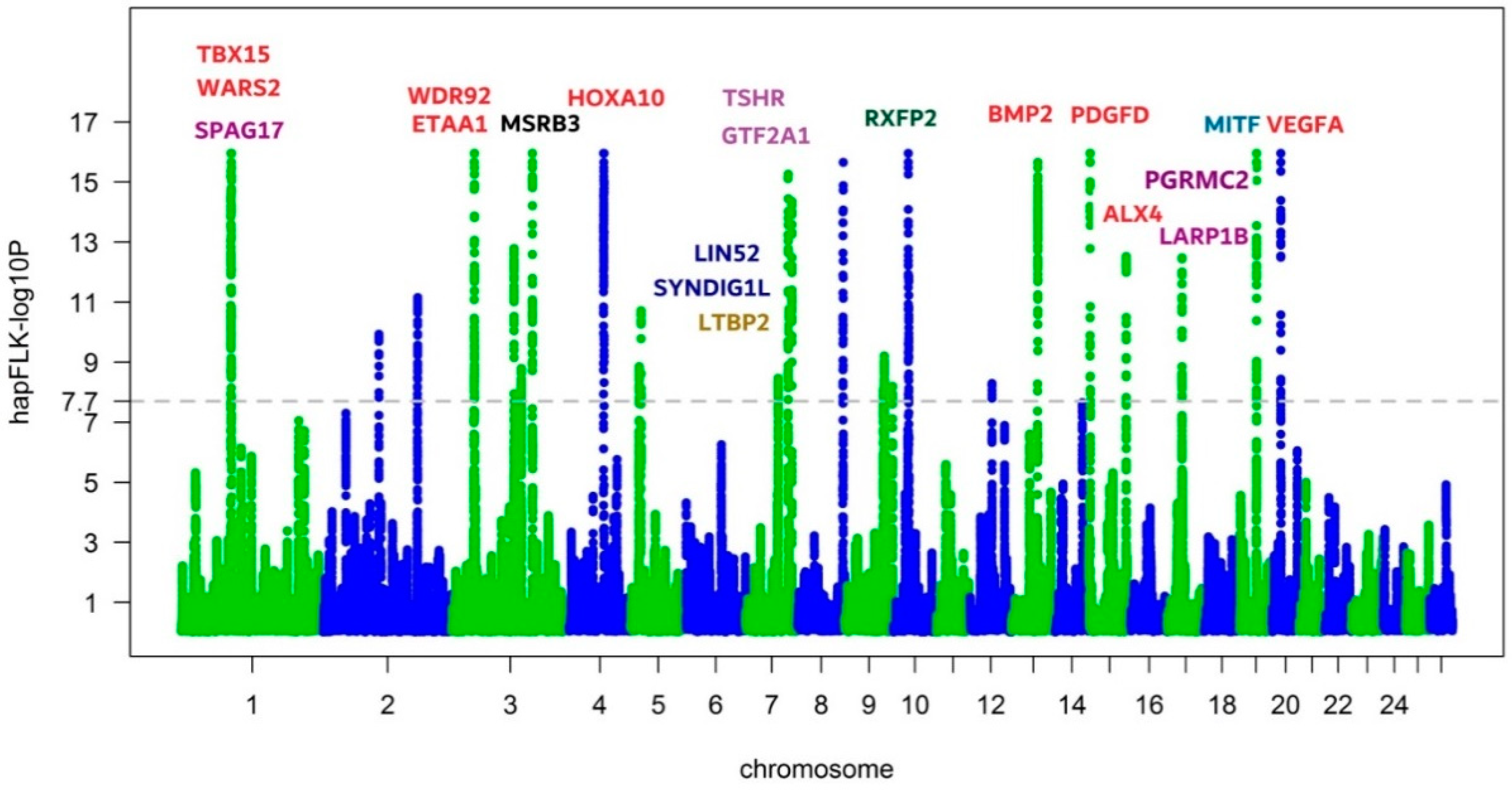
3.2. Identifying the Genomic Regions with Selection Signals Using hapFLK
3.3. Candidate Genes Annotated in Selection Regions
4. Discussion
5. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moradi, M.H.; Nejati-Javaremi, A.; Moradi-Shahrbabak, M.; Dodds, K.G.; McEwan, J.C. Genomic scan of selective sweeps in thin and fat tail sheep breeds for identifying of candidate regions associated with fat deposition. BMC Genet. 2012, 13, 10. [Google Scholar] [CrossRef]
- CNCAGR (China National Commision of Animal Genetic Resource). Animal Genetic Resources in China-Sheep and Goats; China Agriculture Press: Beijing, China, 2012. (In Chinese) [Google Scholar]
- Ermias, E.; Yami, A.; Rege, J. Fat deposition in tropical sheep as adaptive attribute to periodic feed fluctuation. J. Anim. Breed. Genet. 2002, 119, 235–246. [Google Scholar] [CrossRef]
- Safdarian, M.; Zamiri, M.J.; Hashemi, M.; Noorolahi, H. Relationships of fat-tail dimensions with fat-tail weight and carcass characteristics at different slaughter weights of Torki-Ghashghaii sheep. Meat Sci. 2008, 80, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Porto Neto, L.R.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-wide analysis of the world’s sheep breeds reveals high levels of historic mixture and strong recent selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef]
- Fariello, M.I.; Servin, B.; Tosser-Klopp, G.; Rupp, R.; Moreno, C.; International Sheep Genomics, C.; San Cristobal, M.; Boitard, S. Selection signatures in worldwide sheep populations. PLoS ONE 2014, 9, e103813. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Larranaga, O.; Langa, J.; Rendo, F.; Manzano, C.; Iriondo, M.; Estonba, A. Genomic selection signatures in sheep from the Western Pyrenees. Genet. Sel. Evol. 2018, 50, 9. [Google Scholar] [CrossRef]
- Yang, J.; Li, W.R.; Lv, F.H.; He, S.G.; Tian, S.L.; Peng, W.F.; Sun, Y.W.; Zhao, Y.X.; Tu, X.L.; Zhang, M.; et al. Whole-Genome Sequencing of Native Sheep Provides Insights into Rapid Adaptations to Extreme Environments. Mol. Biol. Evol. 2016, 33, 2576–2592. [Google Scholar] [CrossRef]
- Zhao, F.; Wei, C.; Zhang, L.; Liu, J.; Wang, G.; Zeng, T.; Du, L. A genome scan of recent positive selection signatures in three sheep populations. J. Integr. Agric. 2016, 15, 162–174. [Google Scholar] [CrossRef]
- Wei, C.; Wang, H.; Liu, G.; Wu, M.; Cao, J.; Liu, Z.; Liu, R.; Zhao, F.; Zhang, L.; Lu, J.; et al. Genome-wide analysis reveals population structure and selection in Chinese indigenous sheep breeds. BMC Genom. 2015, 16, 194. [Google Scholar] [CrossRef]
- Yuan, Z.; Liu, E.; Liu, Z.; Kijas, J.W.; Zhu, C.; Hu, S.; Ma, X.; Zhang, L.; Du, L.; Wang, H.; et al. Selection signature analysis reveals genes associated with tail type in Chinese indigenous sheep. Anim. Genet. 2017, 48, 55–66. [Google Scholar] [CrossRef]
- Moioli, B.; Pilla, F.; Ciani, E. Signatures of selection identify loci associated with fat tail in sheep. J. Anim. Sci. 2015, 93, 4660–4669. [Google Scholar] [CrossRef] [PubMed]
- Ahbara, A.; Bahbahani, H.; Almathen, F.; Al Abri, M.; Agoub, M.O.; Abeba, A.; Kebede, A.; Musa, H.H.; Mastrangelo, S.; Pilla, F.; et al. Genome-Wide Variation, Candidate Regions and Genes Associated with Fat Deposition and Tail Morphology in Ethiopian Indigenous Sheep. Front. Genet. 2018, 9, 699. [Google Scholar] [CrossRef]
- Zhu, C.; Fan, H.; Yuan, Z.; Hu, S.; Ma, X.; Xuan, J.; Wang, H.; Zhang, L.; Wei, C.; Zhang, Q.; et al. Genome-wide detection of CNVs in Chinese indigenous sheep with different types of tails using ovine high-density 600K SNP arrays. Sci. Rep. 2016, 6, 27822. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Li, M.; Qin, S.; Zhao, F.; Fang, S. Detection of copy number variation and selection signatures on the X chromosome of Chinese indigenous sheep with different tail types. Asian-Australas J. Anim. Sci. 2020. Online. [Google Scholar] [CrossRef]
- Browning, B.L.; Browning, S.R. A unified approach to genotype imputation and haplotype-phase inference for large data sets of trios and unrelated individuals. Am. J. Hum. Genet. 2009, 84, 210–223. [Google Scholar] [CrossRef]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Fariello, M.I.; Boitard, S.; Naya, H.; SanCristobal, M.; Servin, B. Detecting signatures of selection through haplotype differentiation among hierarchically structured populations. Genetics 2013, 193, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, Z.; Cai, Y.; Xu, H.; Yang, R.; Lan, X. Genetic variants in fat- and short-tailed sheep from high-throughput RNA-sequencing data. Anim. Genet. 2018, 49, 483–487. [Google Scholar] [CrossRef]
- Pan, Z.; Li, S.; Liu, Q.; Wang, Z.; Zhou, Z.; Di, R.; An, X.; Miao, B.; Wang, X.; Hu, W.; et al. Rapid evolution of a retro-transposable hotspot of ovine genome underlies the alteration of BMP2 expression and development of fat tails. BMC Genom. 2019, 20, 261. [Google Scholar] [CrossRef]
- Heid, I.M.; Jackson, A.U.; Randall, J.C.; Winkler, T.W.; Qi, L.; Steinthorsdottir, V.; Thorleifsson, G.; Zillikens, M.C.; Speliotes, E.K.; Magi, R.; et al. Meta-analysis identifies 13 new loci associated with waist-hip ratio and reveals sexual dimorphism in the genetic basis of fat distribution. Nat. Genet. 2010, 42, 949–960. [Google Scholar] [CrossRef]
- Johnston, S.E.; McEwan, J.C.; Pickering, N.K.; Kijas, J.W.; Beraldi, D.; Pilkington, J.G.; Pemberton, J.M.; Slate, J. Genome-wide association mapping identifies the genetic basis of discrete and quantitative variation in sexual weaponry in a wild sheep population. Mol. Ecol. 2011, 20, 2555–2566. [Google Scholar] [CrossRef] [PubMed]
- Wiedemar, N.; Drogemuller, C. A 1.8-kb insertion in the 3′-UTR of RXFP2 is associated with polledness in sheep. Anim. Genet. 2015, 46, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Su, G.; Fu, J.; Wang, A.; Liu, J.F.; Lund, M.S.; Guldbrandtsen, B. Introgression of Chinese haplotypes contributed to the improvement of Danish Duroc pigs. Evol. Appl. 2019, 12, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Sha, Y.W.; Mei, L.B.; Ji, Z.Y.; Qiu, P.P.; Ji, H.; Li, P.; Wang, T.; Li, L. A familial study of twins with severe asthenozoospermia identified a homozygous SPAG17 mutation by whole-exome sequencing. Clin. Genet. 2018, 93, 345–349. [Google Scholar] [CrossRef]
- Liu, Z.; Ji, Z.; Wang, G.; Chao, T.; Hou, L.; Wang, J. Genome-wide analysis reveals signatures of selection for important traits in domestic sheep from different ecoregions. BMC Genom. 2016, 17, 863. [Google Scholar] [CrossRef]
- Yuan, J.; Sun, C.; Dou, T.; Yi, G.; Qu, L.; Qu, L.; Wang, K.; Yang, N. Identification of Promising Mutants Associated with Egg Production Traits Revealed by Genome-Wide Association Study. PLoS ONE 2015, 10, e0140615. [Google Scholar] [CrossRef]
- Tan, C.; Wu, Z.; Ren, J.; Huang, Z.; Liu, D.; He, X.; Prakapenka, D.; Zhang, R.; Li, N.; Da, Y.; et al. Genome-wide association study and accuracy of genomic prediction for teat number in Duroc pigs using genotyping-by-sequencing. Genet. Sel. Evol. 2017, 49, 35. [Google Scholar] [CrossRef]
- Verardo, L.L.; Silva, F.F.; Lopes, M.S.; Madsen, O.; Bastiaansen, J.W.; Knol, E.F.; Kelly, M.; Varona, L.; Lopes, P.S.; Guimaraes, S.E. Revealing new candidate genes for reproductive traits in pigs: Combining Bayesian GWAS and functional pathways. Genet. Sel. Evol. 2016, 48, 9. [Google Scholar] [CrossRef]
- Duchemin, S.I.; Bovenhuis, H.; Megens, H.J.; Van Arendonk, J.A.M.; Visker, M. Fine-mapping of BTA17 using imputed sequences for associations with de novo synthesized fatty acids in bovine milk. J. Dairy Sci. 2017, 100, 9125–9135. [Google Scholar] [CrossRef]
- Park, H.B.; Han, S.H.; Lee, J.B.; Cho, I.C. Rapid Communication: High-resolution quantitative trait loci analysis identifies LTBP2 encoding latent transforming growth factor beta binding protein 2 associated with thoracic vertebrae number in a large F2 intercross between Landrace and Korean native pigs. J. Anim. Sci. 2017, 95, 1957–1962. [Google Scholar]
- Chen, C.; Liu, C.; Xiong, X.; Fang, S.; Yang, H.; Zhang, Z.; Ren, J.; Guo, Y.; Huang, L. Copy number variation in the MSRB3 gene enlarges porcine ear size through a mechanism involving miR-584-5p. Genet. Sel. Evol. 2018, 50, 72. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.F.; Xiao, D.; Gong, W.J.; Liu, H.X.; Liu, J.; Zhou, H.H.; Liu, Z.Q. JAZF1 can regulate the expression of lipid metabolic genes and inhibit lipid accumulation in adipocytes. Biochem. Biophys. Res. Commun. 2014, 445, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Proikas-Cezanne, T.; Waddell, S.; Gaugel, A.; Frickey, T.; Lupas, A.; Nordheim, A. WIPI-1alpha (WIPI49), a member of the novel 7-bladed WIPI protein family, is aberrantly expressed in human cancer and is linked to starvation-induced autophagy. Oncogene 2004, 23, 9314–9325. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, G.; Xu, X.; Geng, R.; Zhou, J.; Yang, Y.; Yang, Z.; Chen, Y. Transcriptome profile analysis of adipose tissues from fat and short-tailed sheep. Gene 2014, 549, 252–257. [Google Scholar] [CrossRef]
- Liu, C.T.; Monda, K.L.; Taylor, K.C.; Lange, L.; Demerath, E.W.; Palmas, W.; Wojczynski, M.K.; Ellis, J.C.; Vitolins, M.Z.; Liu, S.; et al. Genome-wide association of body fat distribution in African ancestry populations suggests new loci. PLoS Genet. 2013, 9, e1003681. [Google Scholar] [CrossRef]
- Ravasi, T.; Suzuki, H.; Cannistraci, C.V.; Katayama, S.; Bajic, V.B.; Tan, K.; Akalin, A.; Schmeier, S.; Kanamori-Katayama, M.; Bertin, N.; et al. An atlas of combinatorial transcriptional regulation in mouse and man. Cell 2010, 140, 744–752. [Google Scholar] [CrossRef]
- Kang, D.; Zhou, G.; Zhou, S.; Zeng, J.; Wang, X.; Jiang, Y.; Yang, Y.; Chen, Y. Comparative transcriptome analysis reveals potentially novel roles of Homeobox genes in adipose deposition in fat-tailed sheep. Sci. Rep. 2017, 7, 14491. [Google Scholar] [CrossRef]
- Xu, S.S.; Ren, X.; Yang, G.L.; Xie, X.L.; Zhao, Y.X.; Zhang, M.; Shen, Z.Q.; Ren, Y.L.; Gao, L.; Shen, M.; et al. Genome-wide association analysis identifies the genetic basis of fat deposition in the tails of sheep (Ovis aries). Anim. Genet. 2017, 48, 560–569. [Google Scholar] [CrossRef]
- Zhang, T.; Gao, H.; Sahana, G.; Zan, Y.; Fan, H.; Liu, J.; Shi, L.; Wang, H.; Du, L.; Wang, L.; et al. Genome-wide association studies revealed candidate genes for tail fat deposition and body size in the Hulun Buir sheep. J. Anim. Breed. Genet. 2019, 136, 362–370. [Google Scholar] [CrossRef]
- Johnston, S.E.; Gratten, J.; Berenos, C.; Pilkington, J.G.; Clutton-Brock, T.H.; Pemberton, J.M.; Slate, J. Life history trade-offs at a single locus maintain sexually selected genetic variation. Nature 2013, 502, 93–95. [Google Scholar] [CrossRef]
- de Simoni Gouveia, J.J.; Paiva, S.R.; McManus, C.M.; Caetano, A.R.; Kijas, J.W.; Facó, O.; Azevedo, H.C.; de Araujo, A.M.; de Souza, C.J.H.; Yamagishi, M.E.B. Genome-wide search for signatures of selection in three major Brazilian locally adapted sheep breeds. Livest. Sci. 2017, 197, 36–45. [Google Scholar] [CrossRef]
- Kijas, J.W. Haplotype-based analysis of selective sweeps in sheep. Genome 2014, 57, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Li, S.; Liu, Q.; Wang, Z.; Zhou, Z.; Di, R.; Miao, B.; Hu, W.; Wang, X.; Hu, X.; et al. Whole-genome sequences of 89 Chinese sheep suggest role of RXFP2 in the development of unique horn phenotype as response to semi-feralization. Gigascience 2018, 7, giy019. [Google Scholar] [CrossRef] [PubMed]
- Boyko, A.R.; Quignon, P.; Li, L.; Schoenebeck, J.J.; Degenhardt, J.D.; Lohmueller, K.E.; Zhao, K.; Brisbin, A.; Parker, H.G.; vonHoldt, B.M.; et al. A simple genetic architecture underlies morphological variation in dogs. PLoS Biol. 2010, 8, e1000451. [Google Scholar] [CrossRef]
- Li, M.H.; Tiirikka, T.; Kantanen, J. A genome-wide scan study identifies a single nucleotide substitution in ASIP associated with white versus non-white coat-colour variation in sheep (Ovis aries). Heredity 2014, 112, 122–131. [Google Scholar] [CrossRef]
- Johansson, A.; Marroni, F.; Hayward, C.; Franklin, C.S.; Kirichenko, A.V.; Jonasson, I.; Hicks, A.A.; Vitart, V.; Isaacs, A.; Axenovich, T.; et al. Common variants in the JAZF1 gene associated with height identified by linkage and genome-wide association analysis. Hum. Mol. Genet. 2009, 18, 373–380. [Google Scholar] [CrossRef]
- Zhao, F.; McParland, S.; Kearney, F.; Du, L.; Berry, D.P. Detection of selection signatures in dairy and beef cattle using high-density genomic information. Genet. Sel. Evol. 2015, 47, 49. [Google Scholar] [CrossRef]
- Rubin, C.J.; Zody, M.C.; Eriksson, J.; Meadows, J.R.; Sherwood, E.; Webster, M.T.; Jiang, L.; Ingman, M.; Sharpe, T.; Ka, S.; et al. Whole-genome resequencing reveals loci under selection during chicken domestication. Nature 2010, 464, 587–591. [Google Scholar] [CrossRef]
- Huang, D.W.; Wang, J.X.; Liu, Q.Y.; Chu, M.X.; Di, R.; He, J.N.; Cao, G.L.; Fang, L.; Feng, T.; Li, N. Analysis on DNA sequence of TSHB gene and its association with reproductive seasonality in goats. Mol. Biol. Rep. 2013, 40, 1893–1904. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Large-Tailed Han Sheep | Altay Sheep | Tibetan Sheep | |
|---|---|---|---|
| Tail type | Long fat-tail | Fat-rumped | Long short-tail |
| No. individual | 40 | 40 | 40 |
| Body weight (average kg) | 70.4 kg in male and 60.2 kg in female | 98.3 kg in male and 77.1 kg in female | 51 kg in male and 43.6 kg in female |
| Coat color | White | Brown | White body, heads and feet in miscellaneous colors |
| Horn | Most males have spiral horns and most females have ginger-shaped horns | Males have large horns, most females have horns | Males have large horns, females have horns |
| Region | Liaocheng county, Shan Dong Province | Fuhai county, Xinjiang Uighur autonomous region | Tianzhu county, Gansu Province |
| Geographical coordinates | N35°47′–37°02’ and E115°16´–116°32´ | N40°00´–48°10´ and E87 °00´–89°04´ | N36°31´–37°55´and E102 °07´–103°46´ |
| Climate | Warm temperate continental monsoon climate | Continental arid climate | Continental plateau monsoon climate |
| Altitude (m) | 22–49 | 386–3332 | 2040–4874 |
| Temperature(°C) | −22.3–41.8 | −42.7–40 | −8–4 |
| Use | Meat-fat dual type | Meat-fat dual and coarse wool type | Coarse wool type |
| Chr | Start | End | No. SNP | Size (kb) | Candidate Genes |
|---|---|---|---|---|---|
| 1 | 94756172 | 96112605 | 224 | 1356.433 | SPAG17, RF00026, TBX15, WARS2, RF00100, HAO2, HSD3B1 |
| 2 | 105901381 | 105961277 | 11 | 59.896 | - |
| 2 | 183012215 | 183196527 | 34 | 184.312 | - |
| 3 | 39961457 | 40797601 | 172 | 836.144 | PNO1, WDR92, C1D, ETAA1 |
| 3 | 118322493 | 118505234 | 38 | 182.741 | METTL25, RF00001 |
| 3 | 132458013 | 132563310 | 26 | 105.297 | - |
| 3 | 154055122 | 154324519 | 58 | 269.397 | MSRB3 |
| 4 | 68223979 | 69165823 | 183 | 941.844 | JAZF1, TAX1BP1, EVX1, RF00275, RF01975, RF01976, RF01978, RF01979, RF02040, RF02041, RF02042, RF02043, RF02137, RF02138, RF02139, RF02140, RF02141, RF02142, RF02143, HOXA1, HOXA2, HOXA3, HOXA6, HOXA10, SKAP2 |
| 5 | 16293532 | 16380709 | 19 | 87.177 | ASFB2 |
| 5 | 20143416 | 20250955 | 12 | 107.539 | - |
| 7 | 63493868 | 63648665 | 16 | 154.797 | - |
| 7 | 82373886 | 82795821 | 89 | 421.935 | LIN52, VSX2, ABCD4, SYNDIG1L, NPC2, RF00004, LTBP2 |
| 7 | 89260877 | 89527917 | 51 | 267.04 | TSHR, GTF2A1, RF00600 |
| 8 | 87547094 | 88056486 | 108 | 509.392 | TBXT, MPC1, RPS6KA2 |
| 9 | 75623521 | 75637947 | 2 | 14.426 | - |
| 9 | 77403165 | 77840673 | 47 | 437.508 | VPS13B |
| 9 | 93544798 | 93558759 | 3 | 13.961 | SLC10A5, ENPP2 |
| 10 | 29320784 | 30719200 | 88 | 1398.416 | RXFP2, B3GLCT, HSPH1, TEX26, MEDAG, ALOX5AP, USPL1 |
| 12 | 42361453 | 42380380 | 8 | 18.927 | H6PD |
| 13 | 48400700 | 49177821 | 81 | 777.121 | BMP2 |
| 15 | 3434222 | 3945775 | 68 | 511.553 | PDGFD |
| 15 | 72489270 | 72637084 | 28 | 147.814 | EXT2, RF00026, ALX4 |
| 17 | 29091651 | 29420825 | 44 | 329.174 | PGRMC2, LARP1B |
| 19 | 31539577 | 31850022 | 64 | 310.445 | MITF |
| 20 | 17281001 | 17555177 | 44 | 274.176 | VEGFA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, F.; Deng, T.; Shi, L.; Wang, W.; Zhang, Q.; Du, L.; Wang, L. Genomic Scan for Selection Signature Reveals Fat Deposition in Chinese Indigenous Sheep with Extreme Tail Types. Animals 2020, 10, 773. https://doi.org/10.3390/ani10050773
Zhao F, Deng T, Shi L, Wang W, Zhang Q, Du L, Wang L. Genomic Scan for Selection Signature Reveals Fat Deposition in Chinese Indigenous Sheep with Extreme Tail Types. Animals. 2020; 10(5):773. https://doi.org/10.3390/ani10050773
Chicago/Turabian StyleZhao, Fuping, Tianyu Deng, Liangyu Shi, Wenwen Wang, Qin Zhang, Lixin Du, and Lixian Wang. 2020. "Genomic Scan for Selection Signature Reveals Fat Deposition in Chinese Indigenous Sheep with Extreme Tail Types" Animals 10, no. 5: 773. https://doi.org/10.3390/ani10050773
APA StyleZhao, F., Deng, T., Shi, L., Wang, W., Zhang, Q., Du, L., & Wang, L. (2020). Genomic Scan for Selection Signature Reveals Fat Deposition in Chinese Indigenous Sheep with Extreme Tail Types. Animals, 10(5), 773. https://doi.org/10.3390/ani10050773