Dietary Supplementation with Compound Probiotics and Berberine Alters Piglet Production Performance and Fecal Microbiota

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Compound Probiotics and Berberine Preparation
2.2. Animals Diets and Managements
2.3. Sample Measurements
2.4. DNA Extraction and 16S rRNA Sequencing for Fecal Microbial Community Ananlysis
2.5. Bioinformatics Analysis of Sequencing Data
2.6. Statistical Analysis
3. Results
3.1. Growth Performance, Nutrient Digestibility and Serum Parameters
3.2. Sequencing Data
3.3. Diversity and Compositions of Fecal Microbiota
3.4. Differences in Bacterial Communities
3.5. Association and Model Predictive Analysis
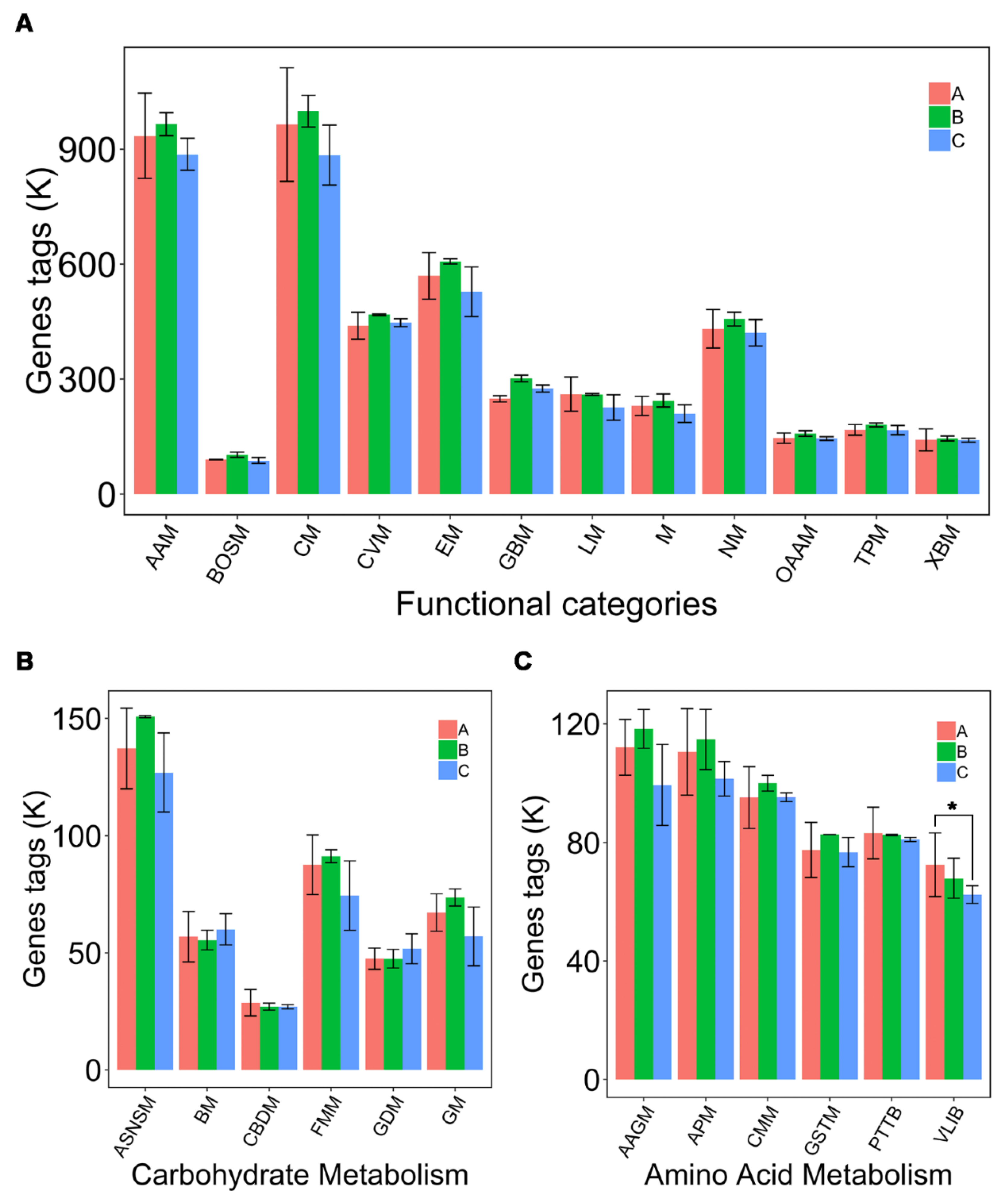
3.6. Predicted Functional Profiles of Microbial Communities Using PICRUSt
4. Discussion
5. Conclusions
Data Availability
Author Contributions
Funding
Conflicts of Interest
References
- Smillie, C.S.; Smith, M.B.; Friedman, J.; Cordero, O.X.; David, L.A.; Alm, E.J. Ecology drives a global network of gene exchange connecting the human microbiome. Nature 2011, 480, 241–244. [Google Scholar] [CrossRef]
- Schokker, D.; Zhang, J.; Zhang, L.L.; Vastenhouw, S.A.; Heilig, H.G.; Smidt, H.; Rebel, J.M.; Smits, M.A. Early-life environmental variation affects intestinal microbiota and immune development in new-born piglets. PLoS ONE 2014, 9, e100040. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Aguilar, Y.M.; Zhang, L.; Ren, W.; Chen, S.; Guan, G.; Xiong, X.; Liao, P.; Li, T.; Huang, R. Dietary supplementation with sanguinarine enhances serum metabolites and antibodies in growing pigs. J. Anim. Sci. 2016, 94, 75–78. [Google Scholar] [CrossRef][Green Version]
- Giang, H.H.; Viet, T.Q.; Ogle, B.; Lindberg, J.E. Effects of different probiotic complexes of lactic acid bacteria on growth performance and gut environment of weaned piglets. Livest. Sci. 2010, 133, 182–184. [Google Scholar] [CrossRef]
- Deng, J.; Li, Y.; Zhang, J.; Yang, Q. Co-administration of Bacillus subtilis RJGP16 and Lactobacillus salivarius B1 strongly enhances the intestinal mucosal immunity of piglets. Res. Vet. Sci. 2013, 94, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chen, L.; Zheng, W.; Shi, M.; Liu, L.; Xie, C.; Wang, X.; Niu, Y.; Hou, Q.; Xu, X.; et al. Lactobacillus frumenti facilitates intestinal epithelial barrier function maintenance in early-weaned piglets. Front. Microbiol. 2018, 9, 897. [Google Scholar] [CrossRef] [PubMed]
- Broadway, P.R.; Carroll, J.A.; Sanchez, N.C.B. Live yeast and yeast cell wall supplements enhance immune function and performance in food-producing livestock: A review. Microorganisms 2015, 3, 417–427. [Google Scholar] [CrossRef]
- Chapman, C.M.; Gibson, G.R.; Rowland, I. Health benefits of probiotics: Are mixtures more effective than single strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef]
- Kumar, A.; Ekavali; Chopra, K.; Mukherjee, M.; Pottabathini, R.; Dhull, D.K. Current knowledge and pharmacological profile of berberine: An update. Eur. J. Pharmacol. 2015, 761, 288–297. [Google Scholar] [CrossRef]
- Battu, S.K.; Repka, M.A.; Maddineni, S.; Chittiboyina, A.G.; Avery, M.A.; Majumdar, S. Physicochemical characterization of berberine chloride: A perspective in the development of a solution dosage form for oral delivery. AAPS Pharm. Sci. 2010, 11, 1466–1475. [Google Scholar] [CrossRef]
- Bhutada, P.; Mundhada, Y.; Bansod, K.; Tawari, S.; Patil, S.; Dixit, P.; Umathe, S.; Mundhada, D. Protection of cholinergic and antioxidant system contributes to the effect of berberine ameliorating memory dysfunction in rat model of streptozotocin-induced diabetes. Behav. Brain Res. 2011, 220, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xie, Q.; Sun, S.; Huang, B.; Zhang, Y.; Xu, Y.; Zhang, S.; Xiang, H. Probiotics-fermented Massa Medicata Fermentata ameliorates weaning stress in piglets related to improving intestinal homeostasis. Appl. Microbiol. Biotechnol. 2018, 102, 10713–10727. [Google Scholar] [CrossRef] [PubMed]
- Lalles, J.P.; Bosi, P.; Smidt, H.; Stokes, C.R. Nutritional management of gut health in pigs around weaning. Proc. Nutr. Soc. 2007, 66, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Xu, H.; Mei, X.; Cao, X.; Gong, L.; Wu, Y.; Li, Y.; Yu, D.; Liu, S.; Wang, Y.; et al. Probiotic Bacillus enhance the intestinal epithelial cell barrier and immune function of piglets. Benef. Microbes 2018, 9, 743–754. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, X.; Liu, P.; Zhao, J.; Sun, J.; Guan, W.; Johnston, L.J.; Levesque, C.L.; Fan, P.; He, T.; et al. Dietary Clostridium butyricum induces a phased shift in fecal microbiota structure and increases the acetic acid-producing bacteria in a weaned piglet model. J. Agric. Food Chem. 2018, 66, 5157–5166. [Google Scholar] [CrossRef]
- Christensen, H.R.; Frokiaer, H.; Pestka, J.J. Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. J. Immunol. 2002, 168, 171–178. [Google Scholar] [CrossRef]
- Yang, J.X.; Zhang, R.F.; Guo, J.P.; Zhang, J.H.; Bai, J.M.; Qiao, L.D. Effects of compound probiotics on growth performance and serum immune index of weaning piglets. Anim. Hus. Vet. Med. 2014, 46, 74–76. [Google Scholar]
- Huang, J.H.; Liang, Z.M.; Ning, G.X.; Wang, S.C. Effects of complex probiotics on growth, nutrients digestibility and serum profiles of weaner piglets. Feed Ind. 2014, 35, 27–31. [Google Scholar] [CrossRef]
- Fouhy, F.; Deane, J.; Rea, M.C.; O’Sullivan, O.; Ross, R.P.; O’Callaghan, G.; Plant, B.J.; Stanton, C. The effects of freezing on faecal microbiota as determined using MiSeq sequencing and culture-based investigations. PLoS ONE 2015, 10, e0119355. [Google Scholar] [CrossRef]
- Baumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef]
- Natividad, J.M.; Verdu, E.F. Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharmacol. Res. 2013, 69, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary sodium butyrate decreases postweaning diarrhea by modulating intestinal permeability and changing the bacterial communities in weaned piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Bruggeman, G.; van den Berg, M.; Borewicz, K.; Scheurink, A.J.; Bruininx, E.; de Vos, P.; Smidt, H.; Schols, H.A.; Gruppen, H. Effects of pectin on fermentation characteristics, carbohydrate utilization, and microbial community composition in the gastrointestinal tract of weaning pigs. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, P.; Wu, Y.; Guo, P.; Liu, L.; Ma, N.; Levesque, C.; Chen, Y.; Zhao, J.; Zhang, J.; et al. Dietary fiber increases butyrate-producing bacteria and improves the growth performance of weaned piglets. J. Agric. Food Chem. 2018, 66, 7995–8004. [Google Scholar] [CrossRef]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R.; et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Microbial shifts in the swine distal gut in response to the treatment with antimicrobial growth promoter, tylosin. Proc. Natl. Acad. Sci. USA 2012, 109, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Pajarillo, E.A.; Chae, J.P.; Balolong, M.P.; Kim, H.B.; Seo, K.S.; Kang, D.K. Pyrosequencing-based analysis of fecal microbial communities in three purebred pig lines. J. Microbiol. 2014, 52, 646–651. [Google Scholar] [CrossRef]
- Kim, H.B.; Isaacson, R.E. The pig gut microbial diversity: Understanding the pig gut microbial ecology through the next generation high throughput sequencing. Vet. Microbiol. 2015, 177, 242–251. [Google Scholar] [CrossRef]
- Tanaka, K.; Tsukahara, T.; Yanagi, T.; Nakahara, S.; Furukawa, O.; Tsutsui, H.; Koshida, S. Bifidobacterium bifidum OLB6378 simultaneously enhances systemic and mucosal humoral immunity in low birth weight infants: A non-randomized study. Nutrients 2017, 9, 195. [Google Scholar] [CrossRef]
- Guo, M.; Wu, F.; Hao, G.; Qi, Q.; Li, R.; Li, N.; Wei, L.; Chai, T. Bacillus subtilis improves immunity and disease resistance in rabbits. Front. Immunol. 2017, 8, 354. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content | Items | Content |
|---|---|---|---|
| Diet compositions | |||
| Soybean oil | 2.0 | Acidulant | 0.1 |
| Corn meal | 50.5 | Lysine | 0.5 |
| Wheat flour | 13.0 | Methionine | 0.25 |
| Riedel wheat | 0.5 | Threonine | 0.15 |
| Puffed soybean | 6.0 | Tryptophan | 0.03 |
| Soybean meal | 18.0 | Stone powder | 0.47 |
| Sugar | 1.0 | Calcium hydrophosphate | 1.0 |
| Glucose | 0.5 | Montmorillonite | 0.4 |
| Salt | 0.3 | Premix | 5.0 |
| Citric acid | 0.3 | Total | 100 |
| Nutrient levels | |||
| CP | 19.79 | Met + Cys | 0.86 |
| DE (MJ/Kg) | 12.86 | Ca | 0.61 |
| Lys | 1.38 | TP | 0.59 |
| Met | 0.57 | AP (Available P) | 0.36 |
| Items | A | B | C | D | E |
|---|---|---|---|---|---|
| Initial weight (kg) | 9.63 ± 0.86 | 9.60 ± 0.83 | 9.56 ± 0.81 | 9.55 ± 0.90 | 9.58 ± 0.57 |
| Final weight (kg) | 25.17 ± 1.67 | 25.12 ± 1.51 | 23.77 ± 3.10 | 23.95 ± 1.22 | 26.61 ± 1.40 |
| ADG (g) | 370.04 ± 29.78 | 369.48 ± 26.85 | 359.82 ± 95.60 | 336.90 ± 21.91 | 405.51 ± 44.76 |
| ADFI (g) | 608.26 ± 0.03 | 625.32 ± 0.08 | 590.61 ± 0.05 | 574.27 ± 0.07 | 602.06 ± 0.05 |
| FCR | 1.64 ± 0.21 | 1.69 ± 0.11 | 1.64 ± 0.22 | 1.70 ± 0.06 | 1.48 ± 0.13 |
| Diarrhea rate (%) | 2.19 | 1.11 | 1.69 | 1.81 | 1.66 |
| Mortality (%) | 5.00 | 3.75 | 1.88 | 2.50 | 3.75 |
| Rejection rate (%) | 7.50 | 0.00 | 0.00 | 0.00 | 0.00 |
| Group | CP | EE | P | Ca |
|---|---|---|---|---|
| A | 69.63 ± 0.15 b | 66.49 ± 2.95 | 78.19 ± 0.44 | 68.83 ± 0.22 |
| B | 71.87 ± 0.84 b | 70.87 ± 4.59 | 77.57 ± 0.48 | 67.51 ± 0.73 |
| C | 79.43 ± 1.57 a | 72.66 ± 4.29 | 76.38 ± 1.51 | 67.44 ± 0.08 |
| D | 79.25 ± 1.01 a | 68.41 ± 3.19 | 77.21 ± 0.29 | 66.48 ± 1.18 |
| E | 80.66 ± 1.95 a | 65.32 ± 1.29 | 78.28 ± 0.08 | 67.21 ± 0.88 |
| Items | A | B | C | D | E |
|---|---|---|---|---|---|
| UN (mmol/L) | 3.79 ± 0.79 ab | 5.31 ± 1.00 a | 3.38 ± 0.38 b | 3.16 ± 0.42 b | 4.57 ± 1.80 ab |
| GLU (mmol/L) | 5.40 ± 0.41 | 6.27 ± 0.86 | 4.94 ± 0.74 | 5.71 ± 1.27 | 5.13 ± 0.71 |
| TP (g/L) | 59.78 ± 3.54 | 58.75 ± 3.34 | 59.15 ± 3.97 | 64.40 ± 4.76 | 62.48 ± 3.75 |
| IgM (g/L) | 0.35 ± 0.18 | 0.20 ± 0.04 | 0.26 ± 0.12 | 0.22 ± 0.03 | 0.31 ± 0.07 |
| IgG (g/L) | 3.27 ± 0.54 b | 4.23 ± 0.74 ab | 4.74 ± 0.56 ab | 5.10 ± 1.72 a | 4.78 ± 1.32 ab |
| ALT (U/L) | 113.25 ± 43.41 a | 47.50 ± 8.35 b | 75.50 ± 14.84 ab | 72.67 ± 12.10 ab | 78.40 ± 31.73 ab |
| AST (U/L) | 74.75 ± 17.58 ab | 51.75 ± 8.26 b | 80.25 ± 22.20 a | 71.33 ± 11.02 ab | 62.80 ± 17.56 ab |
| ALP (U/L) | 382.50 ± 57.91 a | 187.00 ± 51.62 b | 266.75 ± 94.95 ab | 318.00 ± 130.23 ab | 202.00 ± 72.36 b |
| Items | A | B | C |
|---|---|---|---|
| Cellular processing | 347977 | 462088 | 254405 |
| Environmental information processing | 1277640 | 1502226 | 1031051 |
| Genetic information processing | 2262700 | 2662893 | 2076586 |
| Human diseases | 78762 | 93508 | 74579 |
| Metabolism | 5018673 | 6010831 | 4714755 |
| None | 20708 | 26475 | 19759 |
| Organismal systems | 84390 | 102617 | 77360 |
| Unclassified | 1439542 | 1725852 | 1354165 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Yang, C.; Chang, J.; Wang, P.; Yin, Q.; Liu, C.; Gao, T.; Dang, X.; Lu, F. Dietary Supplementation with Compound Probiotics and Berberine Alters Piglet Production Performance and Fecal Microbiota. Animals 2020, 10, 511. https://doi.org/10.3390/ani10030511
Xu X, Yang C, Chang J, Wang P, Yin Q, Liu C, Gao T, Dang X, Lu F. Dietary Supplementation with Compound Probiotics and Berberine Alters Piglet Production Performance and Fecal Microbiota. Animals. 2020; 10(3):511. https://doi.org/10.3390/ani10030511
Chicago/Turabian StyleXu, Xiaoxiang, Canyu Yang, Juan Chang, Ping Wang, Qingqiang Yin, Chaoqi Liu, Tianzeng Gao, Xiaowei Dang, and Fushan Lu. 2020. "Dietary Supplementation with Compound Probiotics and Berberine Alters Piglet Production Performance and Fecal Microbiota" Animals 10, no. 3: 511. https://doi.org/10.3390/ani10030511
APA StyleXu, X., Yang, C., Chang, J., Wang, P., Yin, Q., Liu, C., Gao, T., Dang, X., & Lu, F. (2020). Dietary Supplementation with Compound Probiotics and Berberine Alters Piglet Production Performance and Fecal Microbiota. Animals, 10(3), 511. https://doi.org/10.3390/ani10030511