The Prophylactic Use of Bovine Colostrum in a Murine Model of TNBS-Induced Colitis

 ,
,  ,
,  ,
,  ,
,  ,
,  , , ,
, , , 
Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Bovine Colostrum
2.3. Induction of Colitis
2.4. Clinical Evaluation of Colitis (Disease Activity Index, DAI)
2.5. Tissue Processing
2.6. Histological Analyses and Scoring
2.7. Microbiota Analysis
2.8. Detection and Quantification of Gene Expression
2.9. Statistical Analysis
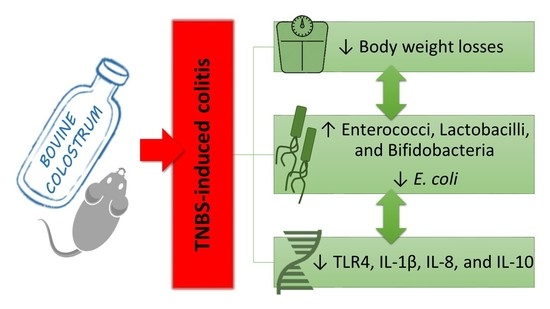
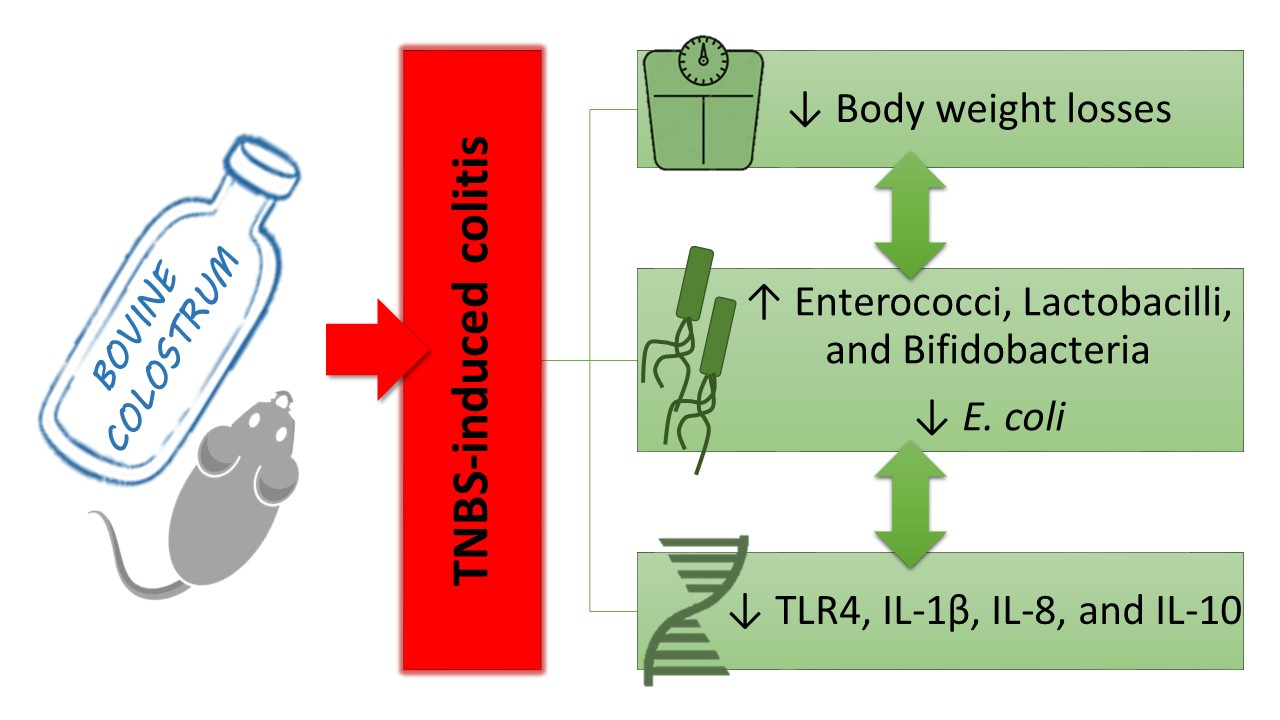
3. Results
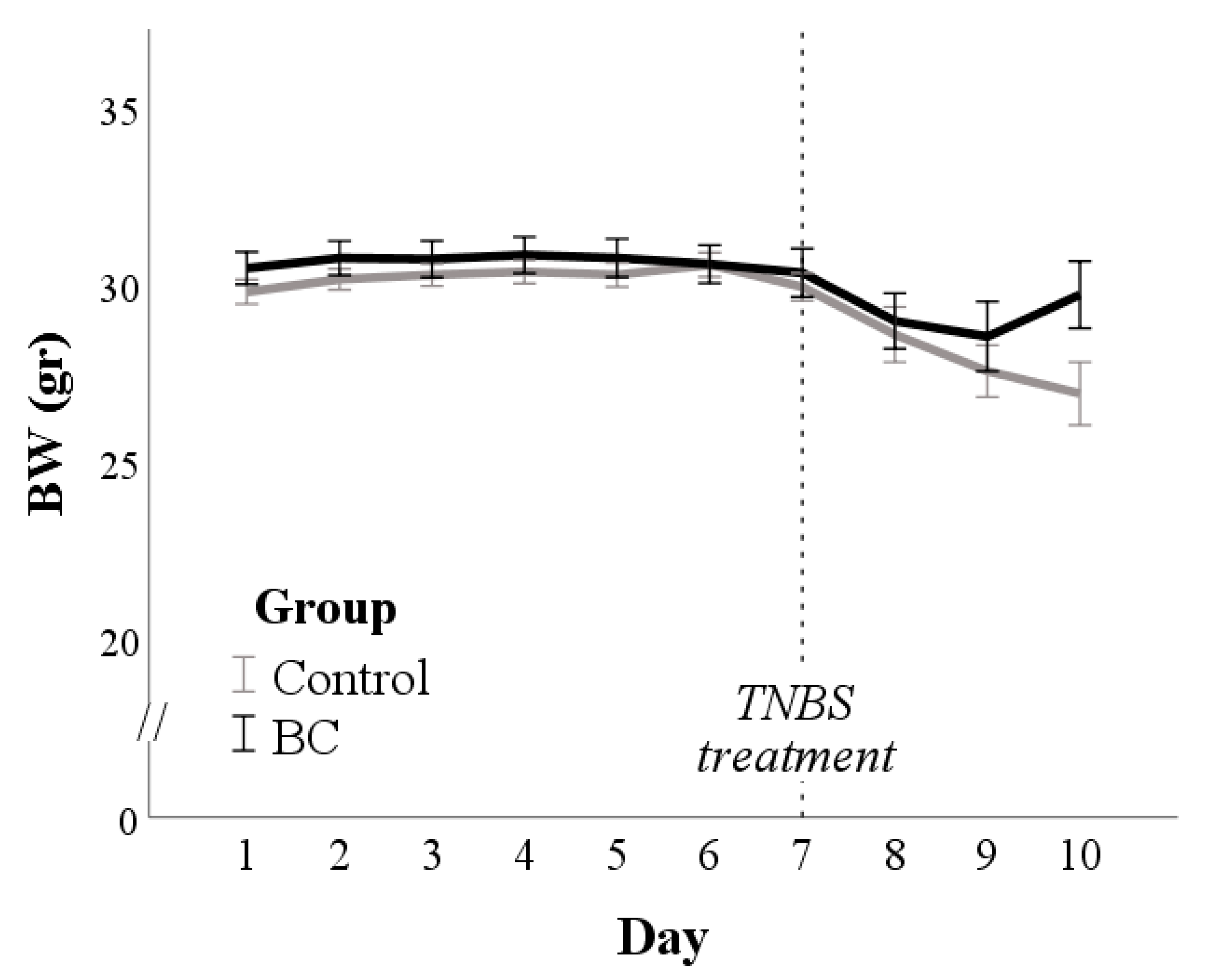
3.1. Body Weight (BW) and Disease Activity Index (DAI)
3.2. Macroscopic Evaluation
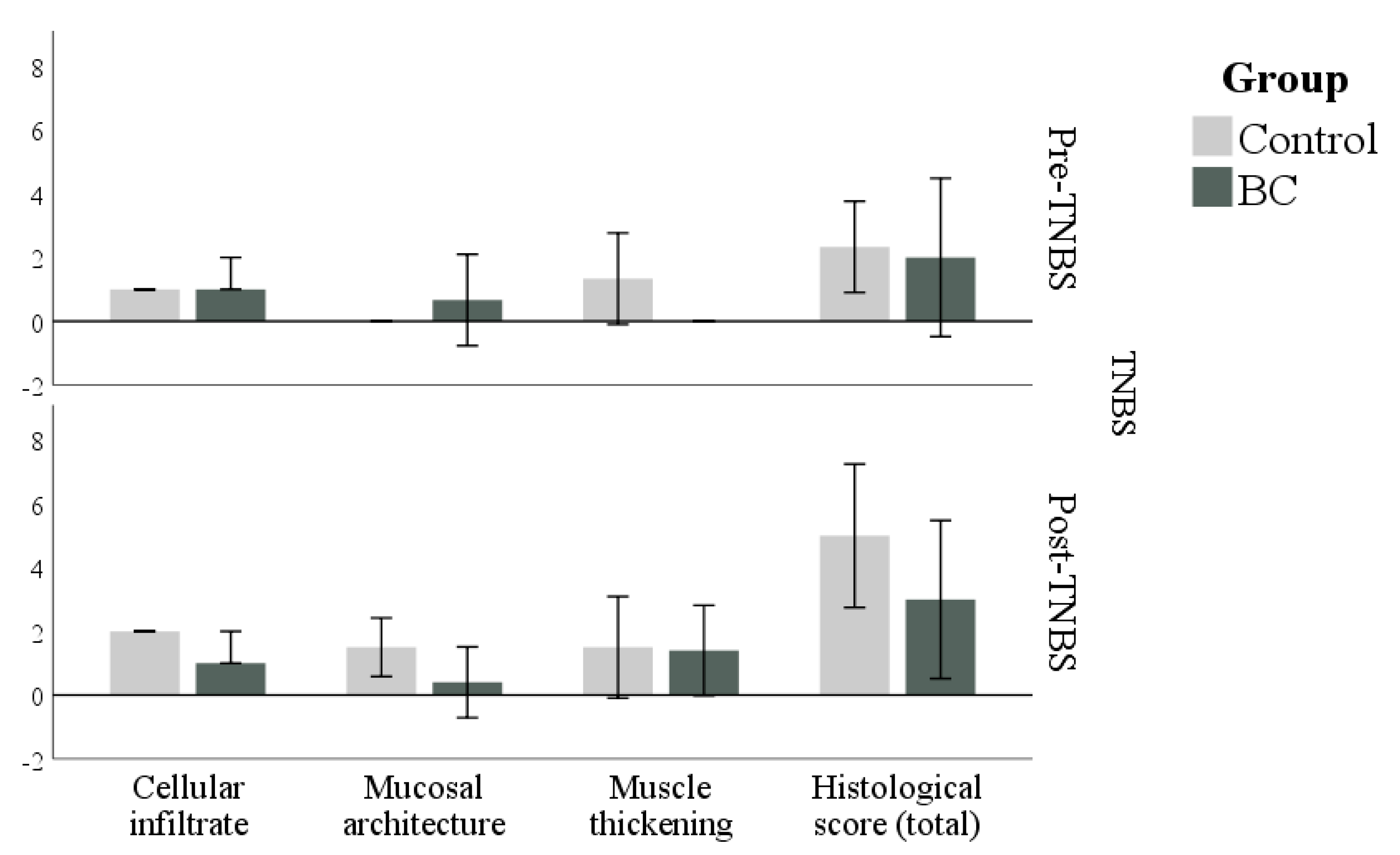
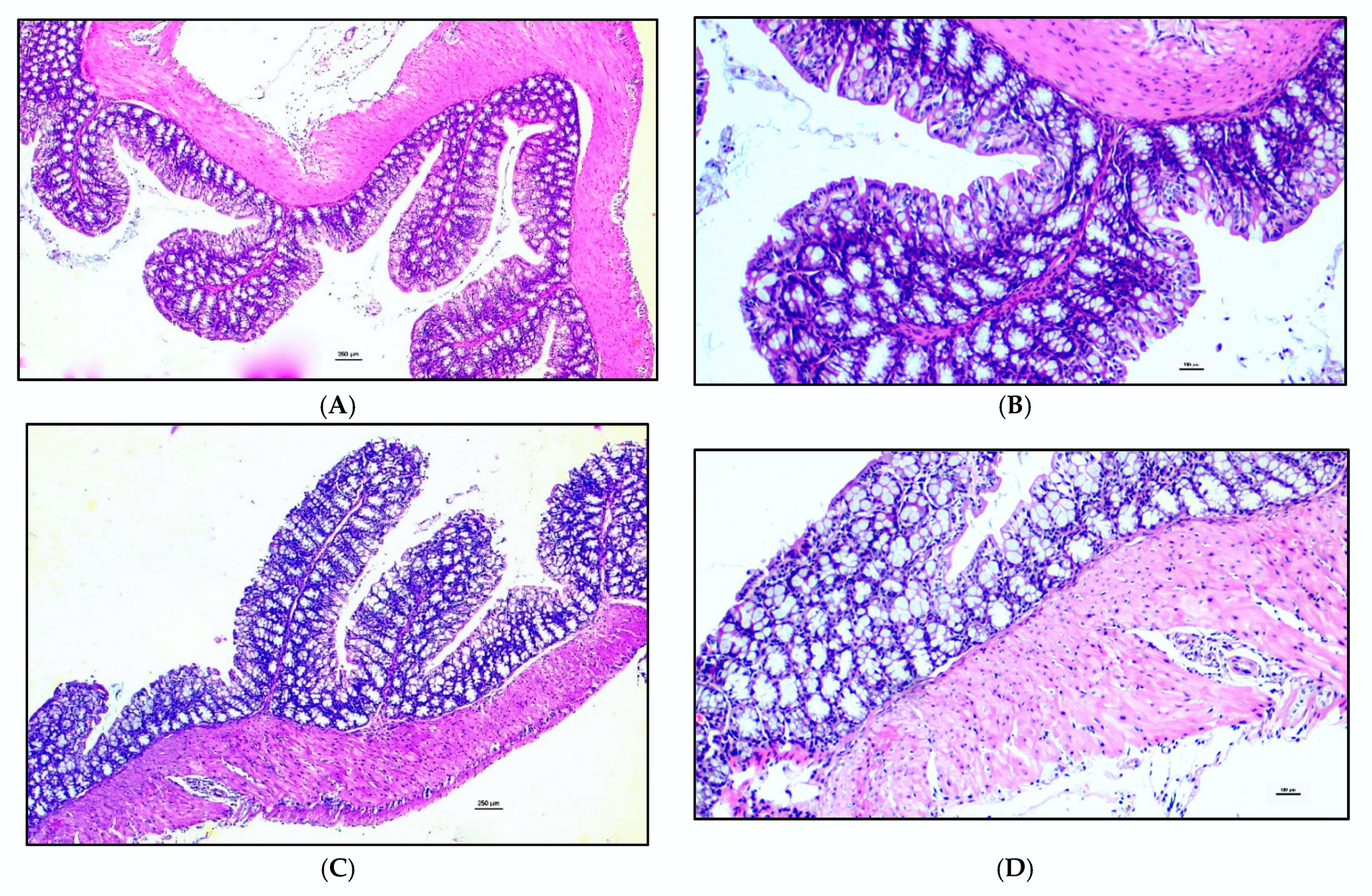
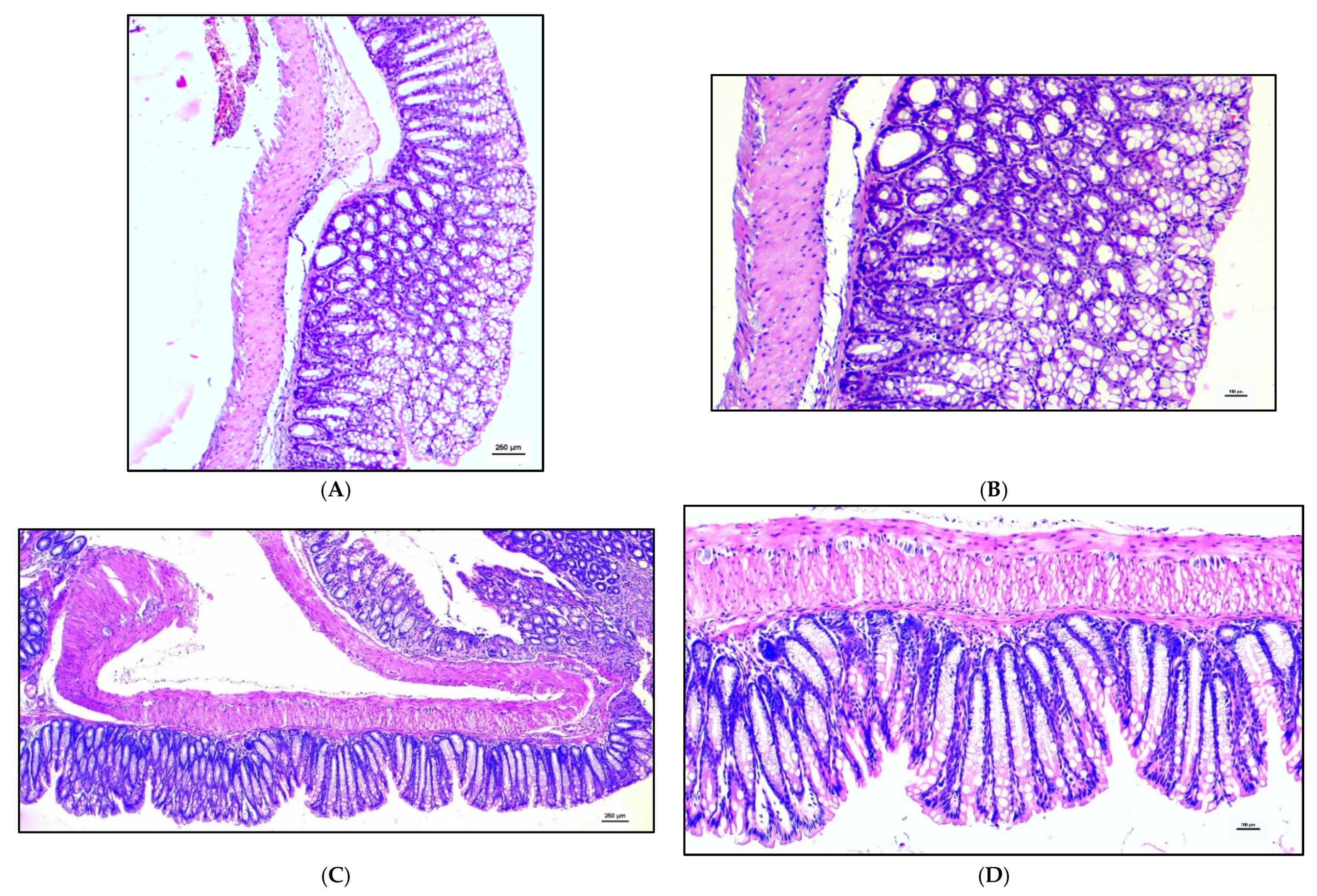
3.3. Histology
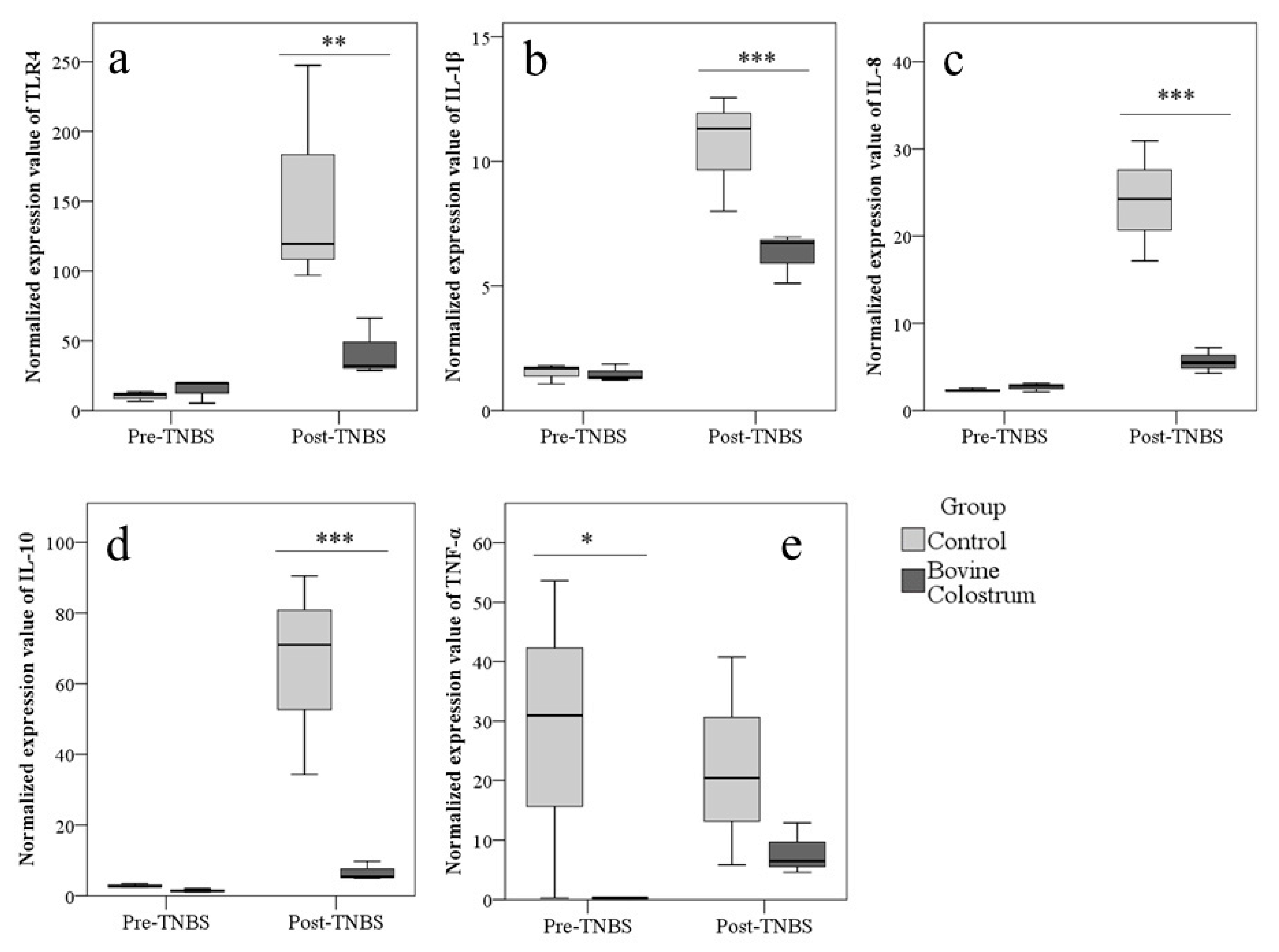
3.4. TLR4 and Cytokines Expression in Colon Evaluated by Real-Time PCR
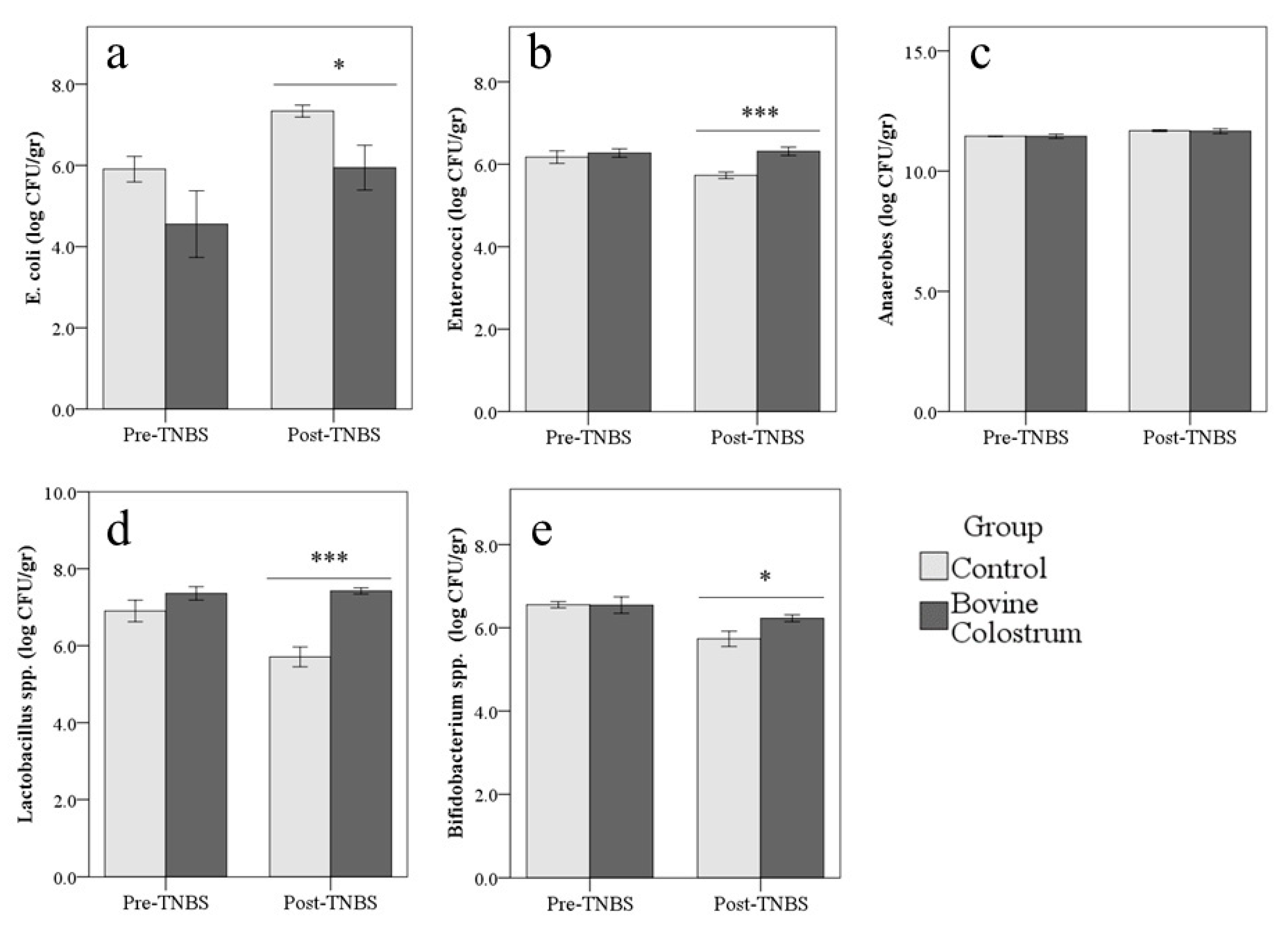
3.5. Gut Microflora
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Langer, P. Differences in the Composition of Colostrum and Milk in Eutherians Reflect Differences in Immunoglobulin Transfer. J. Mammal. 2009, 90, 332–339. [Google Scholar] [CrossRef]
- Ballard, O.; Ardythe, L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. North Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Purup, S.; Vestergaard, M.; Pedersen, L.O.; Sejrsen, K. Biological activity of bovine milk on proliferation of human intestinal cells. J. Dairy Res. 2007, 74, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Playford, R.J. Peptide therapy and the gastroenterologist: Colostrum and milk-derived growth factors. Clin. Nutr. 2001, 20, 101–106. [Google Scholar] [CrossRef]
- Menchetti, L.; Traina, G.; Tomasello, G.; Casagrande-Proietti, P.; Leonardi, L.; Barbato, O.; Brecchia, G. Potential benefits of colostrum in gastrointestinal diseases. Front. Biosci. (Schol. Ed). 2016, 8, 331–351. [Google Scholar]
- Lönnerdal, B. Bioactive proteins in breast milk. J. Paediatr. Child Health 2013, 49, 1–7. [Google Scholar] [CrossRef]
- Khan, Z.; Macdonald, C.; Wicks, A.C.; Holt, M.P.; Floyd, D.; Ghosh, S.; Wright, N.A.; Playford, R.J. Use of the “nutriceutical”, bovine colostrum, for the treatment of distal colitis: Results from an initial study. Aliment. Pharmacol. Ther. 2002, 16, 1917–1922. [Google Scholar] [CrossRef]
- Spalinger, M.R.; Atrott, K.; Baebler, K.; Schwarzfischer, M.; Melhem, H.; Peres, D.R.; Lalazar, G.; Rogler, G.; Scharl, M.; Frey-Wagner, I. Administration of the Hyper-immune Bovine Colostrum Extract IMM-124E Ameliorates Experimental Murine Colitis. J. Crohns. Colitis 2019, 13, 785–797. [Google Scholar] [CrossRef]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef]
- Wallace, K.L.; Zheng, L.; Kanazawa, Y.; Shih, D.Q. Immunopathology of inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 6–21. [Google Scholar] [CrossRef]
- Dragasevic, S.; Stankovic, B.; Milosavljevic, T.; Sokic-Milutinovic, A.; Alempijevic, T.; Zukic, B.; Kotur, N.; Nikcevic, G.; Pavlovic, S.; Popovic, D. Genetic and environmental factors significant fot the presentation and development of inflammatory bowel disease. Eur J Gastroenterol Hepatol 2017, 29, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Triantafillidis, J.K.; Merikas, E.; Georgopoulos, F. Current and emerging drugs for the treatment of inflammatory bowel disease. Drug Des. Devel. Ther. 2011, 5, 185–210. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Kordjazy, N.; Haj-Mirzaian, A.; Haj-Mirzaian, A.; Rohani, M.M.; Gelfand, E.W.; Rezaei, N.; Abdolghaffari, A.H. Role of toll-like receptors in inflammatory bowel disease. Pharmacol. Res. 2017, 129, 204–215. [Google Scholar] [CrossRef]
- Bocker, U.; Yezerskyy, O.; Feick, P.; Manigold, T.; Panja, A.; Kalina, U.; Herweck, F.; Rossol, S.; Singer, M. V Responsiveness of intestinal epithelial cell lines to lipopolysaccharide is correlated with Toll-like receptor 4 but not Toll-like receptor 2 or CD14 expression. Int. J. Colorectal Dis. 2003, 18, 25–32. [Google Scholar] [CrossRef]
- Menchetti, L.; Barbato, O.; Sforna, M.; Vigo, D.; Mattioli, S.; Curone, G.; Tecilla, M.; Riva, F. Brecchia Effects of Diets Enriched in Linseed and Fish Oil on the Expression Pattern of Toll-Like Receptors 4 and Proinflammatory Cytokines on Gonadal Axis and Reproductive Organs in Rabbit Buck. Oxid. Med. Cell. Longev. 2020, 2020, 4327470. [Google Scholar] [CrossRef]
- Brecchia, G.; Menchetti, L.; Cardinali, R.; Castellini, C.; Polisca, A.; Zerani, M.; Maranesi, M.; Boiti, C. Effects of a bacterial lipopolysaccharide on the reproductive functions of rabbit does. Anim. Reprod. Sci. 2014, 147, 128–134. [Google Scholar] [CrossRef]
- Menchetti, L.; Barbato, O.; Filipescu, I.E.; Traina, G.; Leonardi, L.; Polisca, A.; Troisi, A.; Guelfi, G.; Piro, F.; Brecchia, G. Effects of local lipopolysaccharide administration on the expression of Toll-like receptor 4 and pro-inflammatory cytokines in uterus and oviduct of rabbit does. Theriogenology 2018, 107, 162–174. [Google Scholar] [CrossRef]
- Nuding, S.; Fellermann, K.; Wehkamp, J.; Stange, E.F. Reduced mucosal antimicrobial activity in Crohn’s disease of the colon. Gut 2007, 56, 1240–1247. [Google Scholar] [CrossRef]
- Chassaing, B.; Darfeuillemichaud, A. The commensal microbiota and enteropathogens in the pathogenesis of inflammatory bowel diseases. Gastroenterology 2011, 140, 1720–1728. [Google Scholar] [CrossRef]
- Segain, J.-P. Butyrate inhibits inflammatory responses through NFkappa B inhibition: Implications for Crohn’s disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Juhl, S.M.; Ye, X.; Shen, R.L.; Iyore, E.O.; Dai, Y.; Sangild, P.T.; Greisen, G.O. A Stepwise, Pilot Study of Bovine Colostrum to Supplement the First Enteral Feeding in Preterm Infants (Precolos): Study Protocol and Initial Results. Front. Pediatr. 2017, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bodammer, P.; Zirzow, E.; Klammt, S.; Maletzki, C.; Kerkhoff, C. Alteration of DSS-mediated immune cell redistribution in murine colitis by oral colostral immunoglobulin. BioMedCentral Immunol. 2013, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, J.R.; Kanwar, R.K.; Stathopoulos, S.; Haggarty, N.W.; MacGibbon, A.K.H.; Palmano, K.P.; Roy, K.; Rowan, A.; Krissansen, G.W. Comparative activities of milk components in reversing chronic colitis. J. Dairy Sci. 2016, 99, 2488–2501. [Google Scholar] [CrossRef]
- Filipescu, I.E.; Leonardi, L.; Menchetti, L.; Guelfi, G.; Traina, G.; Casagrande-Proietti, P.; Piro, F.; Quattrone, A.; Barbato, O.; Brecchia, G. Preventive effects of bovine colostrum supplementation in TNBS-induced colitis in mice. PLoS One 2018, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Traina, G.; Casagrande-Proietti, P.; Menchetti, L.; Leonardi, L.; Tomasello, G.; Barbato, O.; Piro, F.; Brecchia, G. Colon microbial composition is correlated with the severity of colitis induced by 2,4,6-trinitrobenzenesulfonic acid in mice. EuroMediterranean Biomed. J. 2016, 11, 165–175. [Google Scholar]
- Murano, M.; Maemura, K.; Hirata, I.; Toshina, K.; Nishikawa, T.; Hamamoto, N.; Sasaki, S. Therapeutic effect of intracolonically administered nuclear factor kB (p65) antisense oligonucleotide on mouse dextran sulphate sodium (DSS)-induced colitis. Blackwell Sci. Clin Exp Immunol 2000, 120, 51–58. [Google Scholar] [CrossRef]
- Bellavia, M.; Rappa, F.; Bello, M.L.O.; Brecchia, G.; Tomasello, G.; Leone, A.; Spatola, G.; Uzzo, M.L.; Bonaventura, G.; David, S.; et al. Lactobacillus casei and Bifidobacterium lactis supplementation reduces tissue damage of intestinal mucosa and liver after 2,4,6-trinitrobenzenesulfonic acid treatment in mice. J. Biol. Regul. Homeost. Agents 2014, 28, 251–261. [Google Scholar]
- Traina, G.; Menchetti, L.; Rappa, F.; Casagrande-Proietti, P.; Barbato, O.; Leonardi, L.; Carini, F.; Piro, F.; Brecchia, G. Probiotic mixture supplementation in the preventive management of trinitrobenzenesulfonic acid-induced inflammation in a murine model. J. Biol. Regul. Homeost. Agents 2016, 30, 895–901. [Google Scholar]
- Ya’acov, A.B.; Lichtenstein, Y.; Zolotarov, L.; Ilan, Y. The gut microbiome as a target for regulatory T cell-based immunotherapy: Induction of regulatory lymphocytes by oral administration of anti-LPS enriched colostrum alleviates immune mediated colitis. BMC Gastroenterol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Merga, Y.; Campbell, B.J.; Rhodes, J.M. Mucosal barrier, bacteria and inflammatory bowel disease: Possibilities for therapy. Dig. Dis. 2014, 32, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S.; Poulsen, A.S.R.; Canibe, N.; Lauridsen, C. Effect of bovine colostrum feeding in comparison with milk replacer and natural feeding on the immune responses and colonisation of enterotoxigenic Escherichia coli in the intestinal tissue of piglets. Br. J. Nutr. 2015, 113, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Pontoppidan, P.E.; Shen, R.L.; Cilieborg, M.S.; Jiang, P.; Kissow, H.; Petersen, B.L.; Thymann, T.; Heilmann, C.; Muller, K.; Sangild, P.T. Bovine Colostrum Modulates Myeloablative Chemotherapy-Induced Gut Toxicity in Piglets. J. Nutr. 2015, 145, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.O.; Martin, L.; Østergaard, M.V.; Rudloff, S.; Li, Y.; Roggenbuck, M.; Bering, S.B.; Sangild, P.T. Bovine colostrum improves neonatal growth, digestive function, and gut immunity relative to donor human milk and infant formula in preterm pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G480–G491. [Google Scholar] [CrossRef]
- Zheng, B.; Morgan, M.E.; van de Kant, H.J.G.; Garssen, J.; Folkerts, G.; Kraneveld, A.D. Transcriptional modulation of pattern recognition receptors in chronic colitis in mice is accompanied with Th1 and Th17 response. Biochem. Biophys. Reports 2017, 12, 29–39. [Google Scholar] [CrossRef]
- Tozawa, K.; Hanai, H.; Sugimoto, K.; Baba, S.; Sugimura, H.; Aoshi, T.; Uchijima, M.; Nagata, T.; Koide, Y. Evidence for the critical role of interleukin-12 but not interferon-γ in the pathogenesis of experimental colitis in mice. J. Gastroenterol. Hepatol. 2003, 18, 578–587. [Google Scholar] [CrossRef]
- Støy, A.C.F.; Heegaard, P.M.H.; Thymann, T.; Bjerre, M.; Skovgaard, K.; Boye, M.; Stoll, B.; Schmidt, M.; Jensen, B.B.; Sangild, P.T. Bovine colostrum improves intestinal function following formula-induced gut inflammation in preterm pigs. Clin. Nutr. 2014, 33, 322–329. [Google Scholar] [CrossRef]
- Lopez-Posadas, R.; Requena, P.; Gonzalez, R.; Suarez, M.D.; Zarzuelo, A.; Sanchez de Medina, F.; Martinez-Augustin, O. Bovine Glycomacropeptide Has Intestinal Antiinflammatory Effects in Rats with Dextran Sulfate-Induced Colitis. J. Nutr. 2010, 140, 2014–2019. [Google Scholar] [CrossRef]
- Blais, M.; Fortier, M.; Pouliot, Y.; Gauthier, S.F.; Boutin, Y.; Asselin, C.; Lessard, M. Colostrum whey down-regulates the expression of early and late inflammatory response genes induced by Escherichia coli and Salmonella enterica Typhimurium components in intestinal epithelial cells. Br. J. Nutr. 2018, 113, 200–211. [Google Scholar] [CrossRef]
- An, M.J.; Cheon, J.H.; Kim, S.W.; Park, J.J.; Moon, C.M.; Han, S.Y.; Kim, E.S.; Kim, T.I.; Kim, W.H. Bovine colostrum inhibits nuclear factor κB–mediated proinflammatory cytokine expression in intestinal epithelial cells. Nutr. Res. 2009, 29, 275–280. [Google Scholar] [CrossRef]
- Sun, X.; He, S.; Lv, C.; Sun, X.; Wang, J.; Zheng, W.; Wang, D. Analysis of murine and human Treg subsets in inflammatory bowel disease. Mol. Med. Rep. 2017, 16, 2893–2898. [Google Scholar] [CrossRef] [PubMed]
- Shouval, D.S.; Konnikova, L.; Griffith, A.E.; Wall, S.M.; Biswas, A.; Werner, L.; Nunberg, M.; Kammermeier, J.; Goettel, J.A.; Anand, R.; et al. Enhanced TH17 Responses in Patients with IL10 Receptor Deficiency and Infantile-onset IBD. Inflamm. Bowel Dis. 2017, 23, 1950–1961. [Google Scholar] [CrossRef] [PubMed]
- Godefroy, E.; Alameddine, J.; Montassier, E.; Mathé, J.; Desfrançois-Noël, J.; Marec, N.; Bossard, C.; Jarry, A.; Bridonneau, C.; Le Roy, A.; et al. Expression of CCR6 and CXCR6 by Gut-Derived CD4+/CD8α+ T-Regulatory Cells, Which Are Decreased in Blood Samples From Patients With Inflammatory Bowel Diseases. Gastroenterology 2018, 155, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Schäffler, H.; Kaschitzki, A.; Alberts, C.; Bodammer, P.; Bannert, K.; Köller, T.; Warnke, P.; Kreikemeyer, B.; Lamprecht, G. Alterations in the mucosa-associated bacterial composition in Crohn’s disease: A pilot study. Int. J. Colorectal Dis. 2016, 31, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Sartor, R.B. Microbial Influences in Inflammatory Bowel Diseases. Gastroenterology 2008, 134, 577–594. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- D’Hennezel, E.; Abubucker, S.; Murphy, L.O.; Cullen, T.W. Total Lipopolysaccharide from the Human Gut Microbiome Silences Toll-Like Receptor Signaling. mSystems 2017, 2, e00046-17. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menchetti, L.; Curone, G.; Filipescu, I.E.; Barbato, O.; Leonardi, L.; Guelfi, G.; Traina, G.; Casagrande-Proietti, P.; Riva, F.; Casano, A.B.; et al. The Prophylactic Use of Bovine Colostrum in a Murine Model of TNBS-Induced Colitis. Animals 2020, 10, 492. https://doi.org/10.3390/ani10030492
Menchetti L, Curone G, Filipescu IE, Barbato O, Leonardi L, Guelfi G, Traina G, Casagrande-Proietti P, Riva F, Casano AB, et al. The Prophylactic Use of Bovine Colostrum in a Murine Model of TNBS-Induced Colitis. Animals. 2020; 10(3):492. https://doi.org/10.3390/ani10030492
Chicago/Turabian StyleMenchetti, Laura, Giulio Curone, Iulia Elena Filipescu, Olimpia Barbato, Leonardo Leonardi, Gabriella Guelfi, Giovanna Traina, Patrizia Casagrande-Proietti, Federica Riva, Anna Beatrice Casano, and et al. 2020. "The Prophylactic Use of Bovine Colostrum in a Murine Model of TNBS-Induced Colitis" Animals 10, no. 3: 492. https://doi.org/10.3390/ani10030492
APA StyleMenchetti, L., Curone, G., Filipescu, I. E., Barbato, O., Leonardi, L., Guelfi, G., Traina, G., Casagrande-Proietti, P., Riva, F., Casano, A. B., Piro, F., Vigo, D., Quattrone, A., & Brecchia, G. (2020). The Prophylactic Use of Bovine Colostrum in a Murine Model of TNBS-Induced Colitis. Animals, 10(3), 492. https://doi.org/10.3390/ani10030492