Fermentation Pattern of Several Carbohydrate Sources Incubated in An in Vitro Semicontinuous System with Inocula From Ruminants Given Either Forage or Concentrate-Based Diets

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Substrates and Inocula
2.2. Experimental Conditions
2.3. Chemical and Microbiological Analyses
2.4. Calculations And Statistical Analyses
3. Results
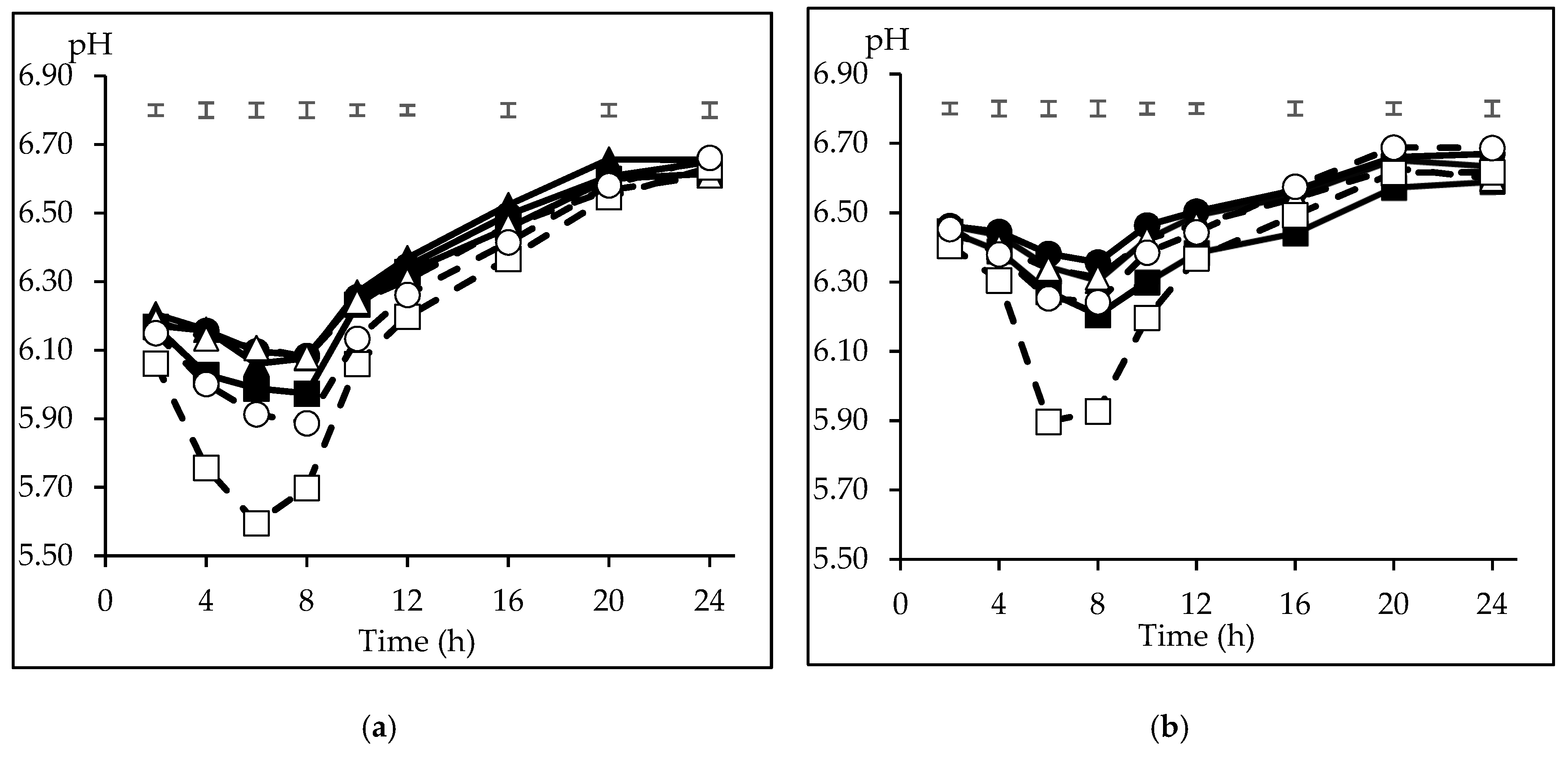
3.1. Pattern of Incubation pH
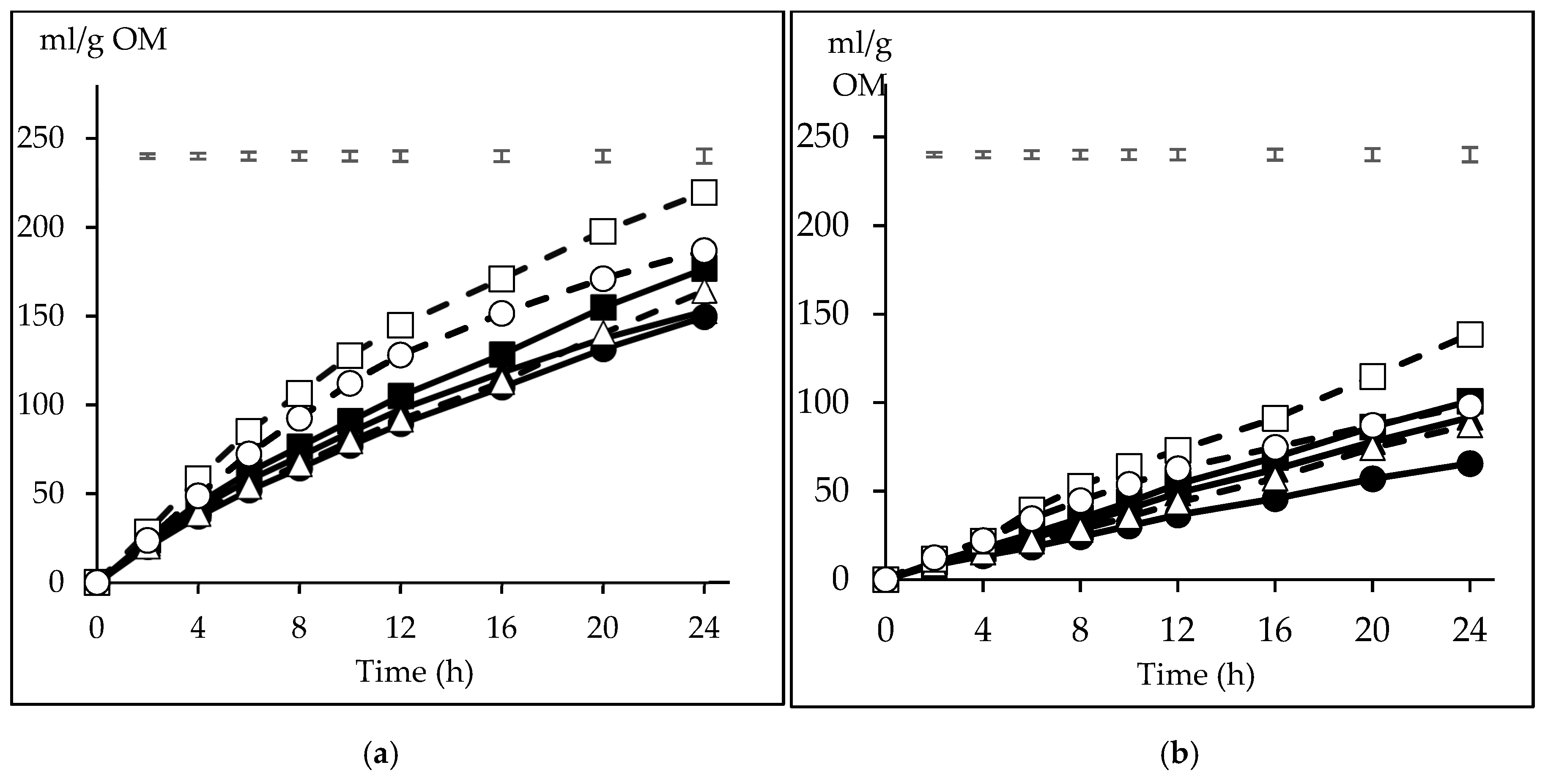
3.2. Pattern of in Vitro Gas Production
3.3. Dry matter Disappearance (DMd)
3.4. Volatile Fatty Acids and Lactic Acid Production
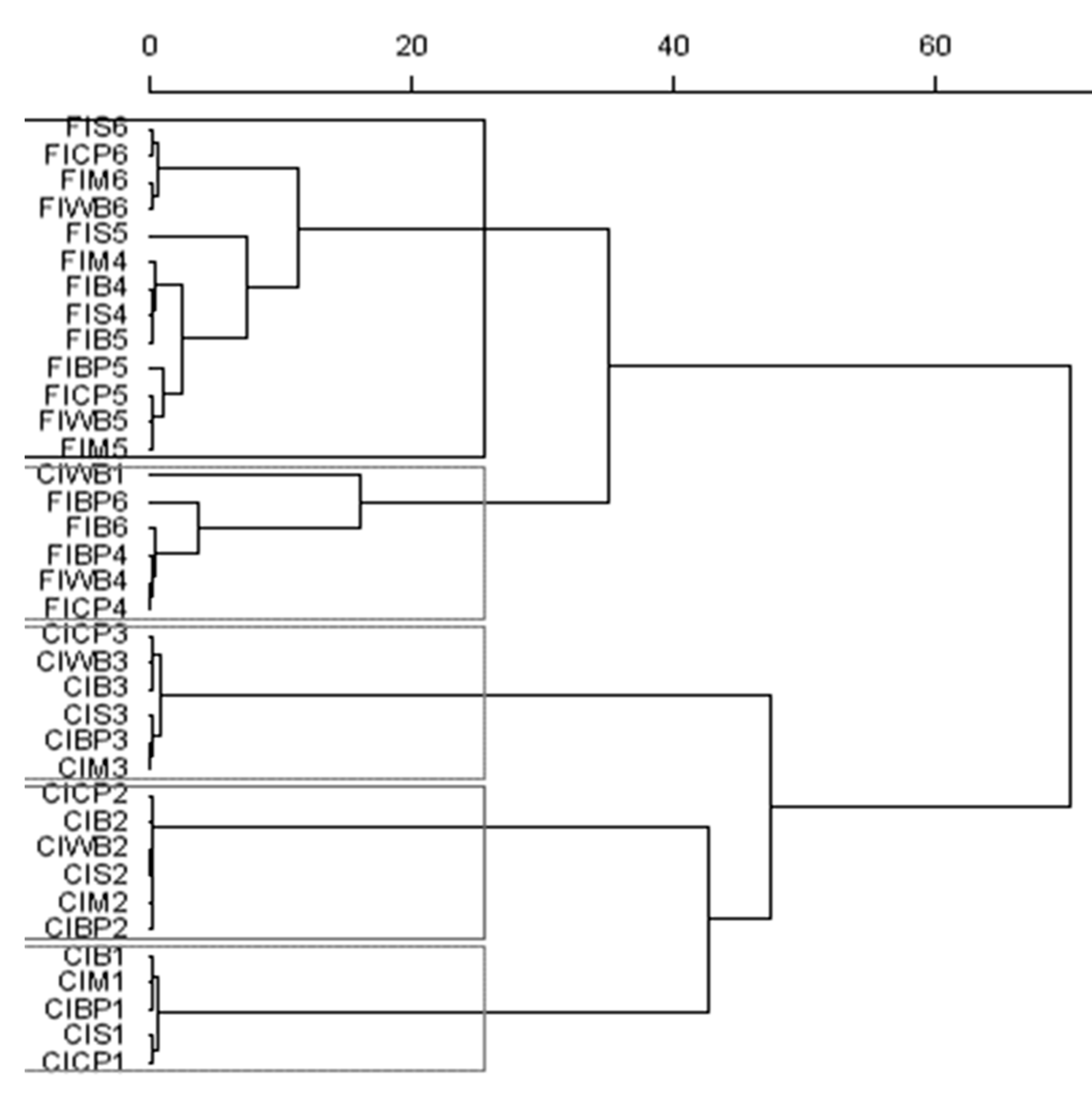
3.5. Bacterial Biodiversity
4. Discussion
4.1. Effect of the Inoculum Source on the in Vitro Fermentation Pattern
4.2. Effect of Different Substrates on the In Vitro Fermentation Pattern
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bevans, D.W.; Beauchemin, K.A.; Schwartzkopf-Genswein, K.S.; McKinnon, J.J.; McAllister, T.A. Effect of rapid or gradual grain adaptation on subacute acidosis and feed intake by feedlot cattle. J. Anim. Sci. 2005, 83, 1116–1132. [Google Scholar] [CrossRef] [PubMed]
- Fernando, S.C.; Purvis, H.T.; Najar, F.Z.; Sukharnikov, L.O.; Krehbiel, C.R.; Nagaraja, T.G.; Roe, B.A.; de Silva, U. Rumen microbial population dynamics during adaptation to a high-grain diet. Appl. Environ. Microbiol. 2010, 76, 7482–7490. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, L. Genotype and environment effects on feed grain quality. Aust. J. Agric. Res. 1999, 50, 703–719. [Google Scholar] [CrossRef]
- Offner, A.; Bach, A.; Sauvant, D. Quantitative review of in situ starch degradation in the rumen. Anim. Feed Sci. Technol. 2003, 106, 81–93. [Google Scholar] [CrossRef]
- De Peeters, E.J.; Fadel, J.G.; Arosemena, A. Digestion kinetics of neutral detergent fiber and chemical composition within some selected by-product feedstuffs. Anim. Feed Sci. Technol. 1997, 67, 127–140. [Google Scholar] [CrossRef]
- Maes, C.; Delcour, J.A. Alkaline hydrogen peroxide extraction of wheat bran non-starch polysaccharides. J. Cereal Sci. 2001, 34, 29–35. [Google Scholar] [CrossRef]
- Mould, F.L.; Kliem, K.E.; Morgan, R.; Mauricio, R.M. In vitro microbial inoculum: A review of its function and properties. Anim. Feed Sci. Technol. 2005, 123–124, 31–50. [Google Scholar] [CrossRef]
- McAllister, T.A.; Cheng, K.J.; Rode, L.M.; Forsberg, C.W. Digestion of barley, maize, and wheat by selected species of ruminal bacteria. Appl. Environ. Microbiol. 1990, 56, 3146–3153. [Google Scholar] [CrossRef]
- Klieve, A.V.; Hennessy, D.; Ouwerkerk, D.; Forster, R.J.; Mackie, R.I.; Attwood, G.T. Establishing populations of Megasphaera elsdenii YE34 and Butyrivibrio fibrisolvens YE44 in the rumen of cattle fed high grain diets. J. Appl. Microbiol. 2003, 95, 621–630. [Google Scholar] [CrossRef]
- Dijkstra, J.; Kebreab, E.; Bannink, A.; France, J.; López, S. Application of the gas production technique to feed evaluation systems for ruminants. Anim. Feed Sci. Technol. 2005, 123–124, 561–578. [Google Scholar] [CrossRef]
- Raab, L.; Cafantaris, B.; Jilg, T.; Menke, K.H. Rumen protein degradation and biosynthesis. A new method for determination of protein degradation in rumen fluid in vitro. Br. J. Nutr. 1983, 50, 569–582. [Google Scholar] [CrossRef]
- Sari, M.; Ferret, A.; Calsamiglia, S. Effect of pH on in vitro microbial fermentation and nutrient flow in diets containing barley straw or non-forage fiber sources. Anim. Feed Sci. Technol. 2015, 200, 17–24. [Google Scholar] [CrossRef]
- Bertipaglia, L.M.A.; Fondevila, M.; van Laar, H.; Castrillo, C. Effect of pelleting and pellet size of a concentrate for intensively reared beef cattle on in vitro fermentation by two different approaches. Anim. Feed Sci. Technol. 2010, 159, 88–95. [Google Scholar] [CrossRef]
- Amanzougarene, Z.; Fondevila, M. Fitting of pH conditions for the study of concentrate feeds fermentation by the in vitro gas production technique. Anim. Prod. Sci. 2018, 58, 1751–1757. [Google Scholar] [CrossRef]
- Amanzougarene, Z.; Yuste, S.; Castrillo, C.; Fondevila, M. In vitro acidification potential and fermentation pattern of different cereal grains incubated with inoculum from animals given forage or concentrate-based diets. Anim. Prod. Sci. 2018, 58, 2300–2307. [Google Scholar] [CrossRef]
- Amanzougarene, Z.; Yuste, S.; de Vega, A.; Fondevila, M. Differences in nutritional characteristics of three varieties of sorghum grain determine their in vitro rumen fermentation. Span. J. Agric. Res. 2018, 16. [Google Scholar] [CrossRef]
- Krause, K.M.; Combs, D.K. Effects of forage particle size, forage source and grain fermentability on performance and ruminal pH in mid lactation cows. J. Dairy Sci. 2003, 86, 1382–1397. [Google Scholar] [CrossRef]
- Fondevila, M.; Pérez-Espés, B. A new in vitro system to study the effect of liquid phase turnover and pH on microbial fermentation of concentrate diets for ruminants. Anim. Feed Sci. Technol. 2008, 144, 196–211. [Google Scholar] [CrossRef]
- Prates, A.; de Oliveira, J.A.; Abecia, L.; Fondevila, M. Effects of preservation procedures of rumen inoculum on in vitro microbial diversity and fermentation. Anim. Feed Sci. Technol. 2010, 155, 186–193. [Google Scholar] [CrossRef]
- Mould, F.L.; Morgan, R.; Kliem, K.E.; Krystallidou, E. A review and simplification of the in vitro incubation medium. Anim. Feed Sci. Technol. 2005, 123–124, 155–172. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; Horwitz, W., Latimer, G.W., Eds.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Mertens, D.R. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beakers or crucibles: Collaborative study. J. AOAC Int. 2002, 85, 1217–1240. [Google Scholar] [PubMed]
- Robertson, J.B.; Van Soest, P.J. The detergent system of analysis and its application to human foods. In The Analysis of Dietary Fiber in Foods; James, W.P.T., Theander, O., Eds.; Marcel Dekker: New York, NY, USA, 1981; pp. 123–158. [Google Scholar]
- Hall, M.B.; Lewis, B.A.; Van Soest, P.J.; Chase, L.E. A simple method for estimation of neutral detergent-soluble fibre. J. Sci.Food Agric. 1997, 74, 441–449. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Blummel, M.; Borowy, N.K.; Becker, K. Gravimetric determination of tannins and their correlations with chemical and protein precipitation methods. J. Sci. Food Agric. 1993, 61, 161–165. [Google Scholar] [CrossRef]
- Barker, S.B.; Summerson, W.H. The colorimetric determination of lactic acid in biological material. J. Biol. Chem. 1941, 138, 535–554. [Google Scholar]
- Hongoh, Y.; Yuzawa, H.; Ohkuma, M.; Kudo, T. Evaluation of primers and PCR conditions for the analysis of 16S rRNA genes from a natural environment. FEMS Microbiol. Lett. 2003, 221, 299–304. [Google Scholar] [CrossRef]
- De la Fuente, G.; Belanche, A.; Girwood, S.E.; Pinloche, E.; Wilkinson, T.; Newbold, C.J. Pros and cons of ion-torrent next generation sequencing versus terminal restriction fragment length polymorphism T-RFLP for studying the rumen bacterial community. PLoS ONE 2014, 9, e101435. [Google Scholar] [CrossRef]
- Analytical Software. Statistix 10 for Windows; Analytical Software: Tallahasee, FL, USA, 2010. [Google Scholar]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef]
- Broudiscou, L.P.; Offner, A.; Sauvant, D. Effects of inoculum source, pH, redox potential and headspace di-hidrogen on rumen in vitro fermentation yields. Animal 2014, 8, 931–937. [Google Scholar] [CrossRef]
- Kim, S.H.; Mamuad, L.L.; Kim, E.J.; Sung, H.G.; Bae, G.S.; Cho, K.K.; Lee, C.; Lee, S.S. Effect of different concentrate diet levels on rumen fluid inoculum used for determination of in vitro rumen fermentation, methane concentration, and methanogen abundance and diversity. Ital. J. Anim. Sci. 2018, 17, 359–367. [Google Scholar] [CrossRef]
- Hiltner, P.; Dehority, B.A. Effect of soluble carbohydrates on digestion of cellulose by pure cultures of rumen bacteria. Appl. Environ. Microbiol. 1983, 46, 642–648. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H. Estimation of the energy feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Nagadi, S.; Herrero, M.; Jessop, N.S. The influence of diet of the donor animal on the initial bacterial concentration of ruminal fluid and in vitro gas production degradability parameters. Anim. Feed Sci. Technol. 2000, 87, 231–239. [Google Scholar] [CrossRef]
- Hatfield, R.D.; Weimer, P.J. Degradation characteristics of isolated and in situ cell wall lucerne pectic polysaccharides by mixed ruminal microbes. J. Sci. Food Agric. 1995, 69, 185–196. [Google Scholar] [CrossRef]
- Barrios-Urdaneta, A.; Fondevila, M.; Castrillo, C. Effect of supplementation with different proportions of barley grain or citrus pulp on the digestive utilization of ammonia-treated straw by sheep. Anim. Sci. 2003, 76, 309–317. [Google Scholar] [CrossRef]
- Mould, F.L.; Ørskov, E.R. Manipulation of rumen fluid pH and its influence on cellulolysis in sacco, dry matter degradation and the ruminal microflora of sheep offered either hay or concentrate. Anim. Feed Sci. Technol. 1983, 10, 1–14. [Google Scholar] [CrossRef]
- Grant, R.H.; Mertens, D.R. Influence of buffer pH and raw corn starch addition on in vitro fiber digestion kinetics. J. Dairy Sci. 1992, 75, 2762–2768. [Google Scholar] [CrossRef]
- Cerneau, P.; Michalet-Doreau, B. In situ starch degradation of different feeds in the rumen. Reprod. Nutr. Dev. 1991, 31, 65–72. [Google Scholar] [CrossRef]
- Demarquilly, C.; Andrieu, J. Les fourrages. In Alimentation des Bovins Ovins et Caprins; INRA: Paris, France, 1988; Volume 3, pp. 57–74. [Google Scholar]
- Amanzougarene, Z.; Yuste, S.; de Vega, A.; Fondevila, M. In vitro fermentation pattern and acidification, potential of different sources of carbohydrates by ruminants giving high concentrate diets. Span. J. Agric. Res. 2017, 15, e0602. [Google Scholar] [CrossRef]
- Calsamiglia, S.; Cardozo, P.W.; Ferret, A.; Bach, A. Changes in rumen microbial fermentation are due to a combined effect of type of diet and pH. J. Anim. Sci. 2008, 86, 702–711. [Google Scholar] [CrossRef]
- Saro, C.; Ranilla, M.J.; Tejido, M.L.; Carro, M.D. Influence of forage type in the diet of sheep on rumen microbiota and fermentation characteristics. Livest. Sci. 2014, 160, 52–59. [Google Scholar] [CrossRef]
- Tapio, I.; Fischer, D.; Blasco, L.; Tapio, M.; Wallace, R.J.; Bayat, A.R.; Ventto, L.; Kahala, M.; Negussie, E.; Shingfield, K.J.; et al. Taxon abundance, diversity, co-occurrence and network analysis of the ruminal microbiota in response to dietary changes in dairy cows. PLoS ONE 2017, 12, e0180260. [Google Scholar] [CrossRef] [PubMed]
- Nagata, R.; Kim, Y.H.; Ohkubo, A.; Kushibiki, S.; Ichijo, T.; Sato, S. Effects of repeated subacute ruminal acidosis challenges on the adaptation of the rumen bacterial community in Holstein bulls. J. Dairy Sci. 2018, 101, 4424–4436. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Hino, T. Regulation of lactate production in streptococcus bovis—A spiraling effect that contributes to rumen acidosis. J. Dairy Sci. 1985, 68, 1712–1721. [Google Scholar] [CrossRef]
- Sauvant, D.; Giger-Reverdin, S.; Meschy, F. Le contrôle de l’acidose ruminale latente. INRA Prod. Anim. 2006, 19, 69–78. [Google Scholar]
- Krause, K.M.; Oetzel, G.R. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Technol. 2006, 126, 215–236. [Google Scholar] [CrossRef]
- Hall, M.B.; Pell, A.N.; Chase, L.E. Characteristics of neutral detergent-soluble fiber fermentation by mixed ruminal microbes. Anim. Feed Sci. Technol. 1998, 70, 23–39. [Google Scholar] [CrossRef]
- Ariza, P.; Bach, A.; Stern, M.D.; Hall, M.B. Effects of carbohydrates from citrus pulp and hominy feed on microbial fermentation in continuous culture. J. Anim. Sci. 2001, 79, 2713–2718. [Google Scholar] [CrossRef]
- Bampidis, V.A.; Robinson, P.H. Citrus by-products as ruminant feeds: A review. Anim. Feed Sci. Technol. 2006, 128, 175–217. [Google Scholar] [CrossRef]
- Strobel, H.J.; Russel, J.B. Effects of pH and energy spilling on bacterial protein synthesis by carbohydrate-limited cultures of mixed rumen bacteria. J. Dairy Sci. 1986, 69, 2941–2947. [Google Scholar] [CrossRef]
- Sauvant, D.; Dorleans, M.; Delacour, C.; Bertrand, D.; Giger, S. La modélisation des cinétiques de dégradation des constituants pariétaux et cellulaires des aliments dans le rumen. Reprod. Nutr. Dev. 1986, 26, 303–304. [Google Scholar] [CrossRef]
- Nocek, J.E.; Tamminga, S. Site of digestion of starch in the gastrointestinal tract of dairy cows and its effect on milk yield and composition. J. Dairy Sci. 1991, 74, 3598–3629. [Google Scholar] [CrossRef]
- McAllister, T.A.; Philippe, R.C.; Rode, L.M.; Cheng, K.J. Effect of the protein matrix on the digestion of cereal grains by ruminal microorganisms. J. Anim. Sci. 1993, 71, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Overton, T.R.; Cameron, M.R.; Elliott, J.P.; Clark, J.H.; Nelson, D.R. Ruminal fermentation and passage of nutrients to the duodenum of lactating cows fed mixtures of corn and barley. J. Dairy Sci. 1995, 78, 1981–1998. [Google Scholar] [CrossRef]
- Firkins, J.L.; Eastridge, M.L.; St-Pierre, N.R.; Noftsger, S.M. Effects of grain variability and processing on starch utilization by lactating dairy cattle. J. Anim. Sci. 2001, 79, E218–E238. [Google Scholar] [CrossRef]
- Lanzas, C.; Fox, D.G.; Pell, A.N. Digestion kinetics of dried cereal grains. Anim. Feed Sci. Technol. 2007, 136, 265–280. [Google Scholar] [CrossRef]
- Opatpatanakit, Y.; Kellaway, R.C.; Lean, I.J.; Annison, G.; Kirby, A. Microbial fermentation of cereal grains in vitro. Aust. J. Agric. Res. 1994, 45, 1247–1263. [Google Scholar] [CrossRef]
- Taxis, T.M.; Wolff, S.; Gregg, S.J.; Minton, N.O.; Zhang, C.; Dai, J.; Schnabel, R.D.; Taylor, J.F.; Kerley, M.S.; Pires, J.C.; et al. The players may change but the game remains: Network analyses of ruminal microbiomes suggest taxonomic differences mask functional similarity. Nucleic Acids Res. 2015, 43, 9600–9612. [Google Scholar] [CrossRef]
- Söllinger, A.; Tveit, A.T.; Poulsen, M.; Noel, S.J.; Bengtsson, M.; Bernhardt, J.; Frydendahl Hellwing, A.L.; Lund, P.; Riedel, K.; Schleper, C.; et al. Holistic assessment of rumen microbiome dynamics through quantitative metatranscriptomics reveals multifunctional redundancy during key steps of anaerobic feed degradation. mSystems 2018, 3. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Component | B | M | S | BP | CP | WB |
|---|---|---|---|---|---|---|
| OM | 978 | 986 | 979 | 953 | 940 | 944 |
| CP | 105 | 75 | 113 | 107 | 59 | 161 |
| EE | 24 | 34 | 11 | 5 | 14 | 31 |
| Starch | 672 | 706 | 647 | - | - | 245 |
| aNDFom | 173 | 91 | 97 | 437 | 207 | 499 |
| ADF | 56 | 25 | 60 | 272 | 192 | 145 |
| ADL | 18 | 2 | 5 | 75 | 21 | 37 |
| NDSF | 4 | 77 | 110 | 457 | 423 | 155 |
| Sugars | 1.6 | 13 | 1.3 | 9 | 243 | 31 |
| TP | - | - | 2.6 | - | - | - |
| TT | - | - | 1.3 | - | - | - |
| Substrates | VFA | Acetate | Propionate | Butyrate | Valerate | BCVFA | Lactic acid |
|---|---|---|---|---|---|---|---|
| with CI | |||||||
| B | 22.34 ab | 0.570 | 0.240 | 0.153 | 0.022 | 0.014 | 3.83 b |
| M | 20.18 bc | 0.578 | 0.235 | 0.149 | 0.023 | 0.016 | 2.35 c |
| S | 21.72 abc | 0.588 | 0.245 | 0.131 | 0.022 | 0.015 | 1.93 c |
| BP | 21.66 abc | 0.593 | 0.235 | 0.135 | 0.021 | 0.016 | 0.90 c |
| CP | 26.55 a | 0.595 | 0.238 | 0.135 | 0.020 | 0.012 | 8.70 a |
| WB | 26.94 a | 0.590 | 0.245 | 0.132 | 0.020 | 0.013 | 2.95 c |
| With FI | |||||||
| B | 10.31 xyz | 0.633 | 0.226 | 0.102 | 0.009 | 0.030 y | 3.05 y |
| M | 8.83 yz | 0.632 | 0.229 | 0.095 | 0.010 | 0.034 xy | 2.69 y |
| S | 8.31 z | 0.642 | 0.216 | 0.094 | 0.009 | 0.039 x | 0.84 y |
| BP | 9.52 yx | 0.665 | 0.207 | 0.088 | 0.008 | 0.032 y | 1.64 y |
| CP | 12.12 yz | 0.686 | 0.209 | 0.075 | 0.008 | 0.023 z | 8.67 x |
| WB | 9.83 yz | 0.653 | 0.225 | 0.083 | 0.009 | 0.030 y | 2.97 y |
| SEM | 1.065 | 0.0156 | 0.0076 | 0.0085 | 0.0010 | 0.0009 | 0.588 |
| p-Value | |||||||
| Inoculum | 0.002 | 0.077 | NS | NS | 0.005 | <0.001 | NS |
| Substrate | <0.001 | NS | NS | NS | NS | <0.001 | <0.001 |
| Inoc. × Subs. | NS | NS | NS | NS | NS | <0.001 | NS |
| Substrates | VFA | Acetate | Propionate | Butyrate | Valerate | BCVFA |
|---|---|---|---|---|---|---|
| With CI | ||||||
| B | 21.16 b | 0.561 | 0.225 | 0.172 | 0.028 | 0.014 |
| M | 19.42 b | 0.548 | 0.227 | 0.178 | 0.031 | 0.017 |
| S | 19.87 b | 0.557 | 0.249 | 0.152 | 0.026 | 0.017 |
| BP | 20.25 b | 0.604 | 0.229 | 0.130 | 0.022 | 0.016 |
| CP | 27.32 a | 0.553 | 0.240 | 0.164 | 0.031 | 0.012 |
| WB | 25.10 ab | 0.537 | 0.261 | 0.158 | 0.029 | 0.015 |
| With FI | ||||||
| B | 10.46 | 0.597 | 0.256 xy | 0.117 | 0.009 | 0.022 |
| M | 8.51 | 0.620 | 0.236 xy | 0.109 | 0.009 | 0.026 |
| S | 8.48 | 0.620 | 0.222 y | 0.120 | 0.009 | 0.029 |
| BP | 8.34 | 0.611 | 0.227 y | 0.126 | 0.009 | 0.027 |
| CP | 10.47 | 0.579 | 0.267 x | 0.124 | 0.010 | 0.019 |
| WB | 9.76 | 0.575 | 0.274 x | 0.117 | 0.010 | 0.025 |
| SEM | 1.137 | 0.0233 | 0.0078 | 0.0169 | 0.0019 | 0.0024 |
| p-Value | ||||||
| Inoculum | 0.011 | NS | NS | NS | 0.008 | 0.008 |
| Substrate | 0.001 | NS | <0.001 | NS | NS | 0.046 |
| Inoc. × Subs. | 0.058 | NS | 0.017 | NS | NS | NS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amanzougarene, Z.; Yuste, S.; Fondevila, M. Fermentation Pattern of Several Carbohydrate Sources Incubated in An in Vitro Semicontinuous System with Inocula From Ruminants Given Either Forage or Concentrate-Based Diets. Animals 2020, 10, 261. https://doi.org/10.3390/ani10020261
Amanzougarene Z, Yuste S, Fondevila M. Fermentation Pattern of Several Carbohydrate Sources Incubated in An in Vitro Semicontinuous System with Inocula From Ruminants Given Either Forage or Concentrate-Based Diets. Animals. 2020; 10(2):261. https://doi.org/10.3390/ani10020261
Chicago/Turabian StyleAmanzougarene, Zahia, Susana Yuste, and Manuel Fondevila. 2020. "Fermentation Pattern of Several Carbohydrate Sources Incubated in An in Vitro Semicontinuous System with Inocula From Ruminants Given Either Forage or Concentrate-Based Diets" Animals 10, no. 2: 261. https://doi.org/10.3390/ani10020261
APA StyleAmanzougarene, Z., Yuste, S., & Fondevila, M. (2020). Fermentation Pattern of Several Carbohydrate Sources Incubated in An in Vitro Semicontinuous System with Inocula From Ruminants Given Either Forage or Concentrate-Based Diets. Animals, 10(2), 261. https://doi.org/10.3390/ani10020261