Polymorphisms of the ACSL1 Gene Influence Milk Production Traits and Somatic Cell Score in Chinese Holstein Cows

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data and Animal Sample Collection
2.2. SNP Discovery and Genotyping
2.3. Statistical Analyses
3. Results
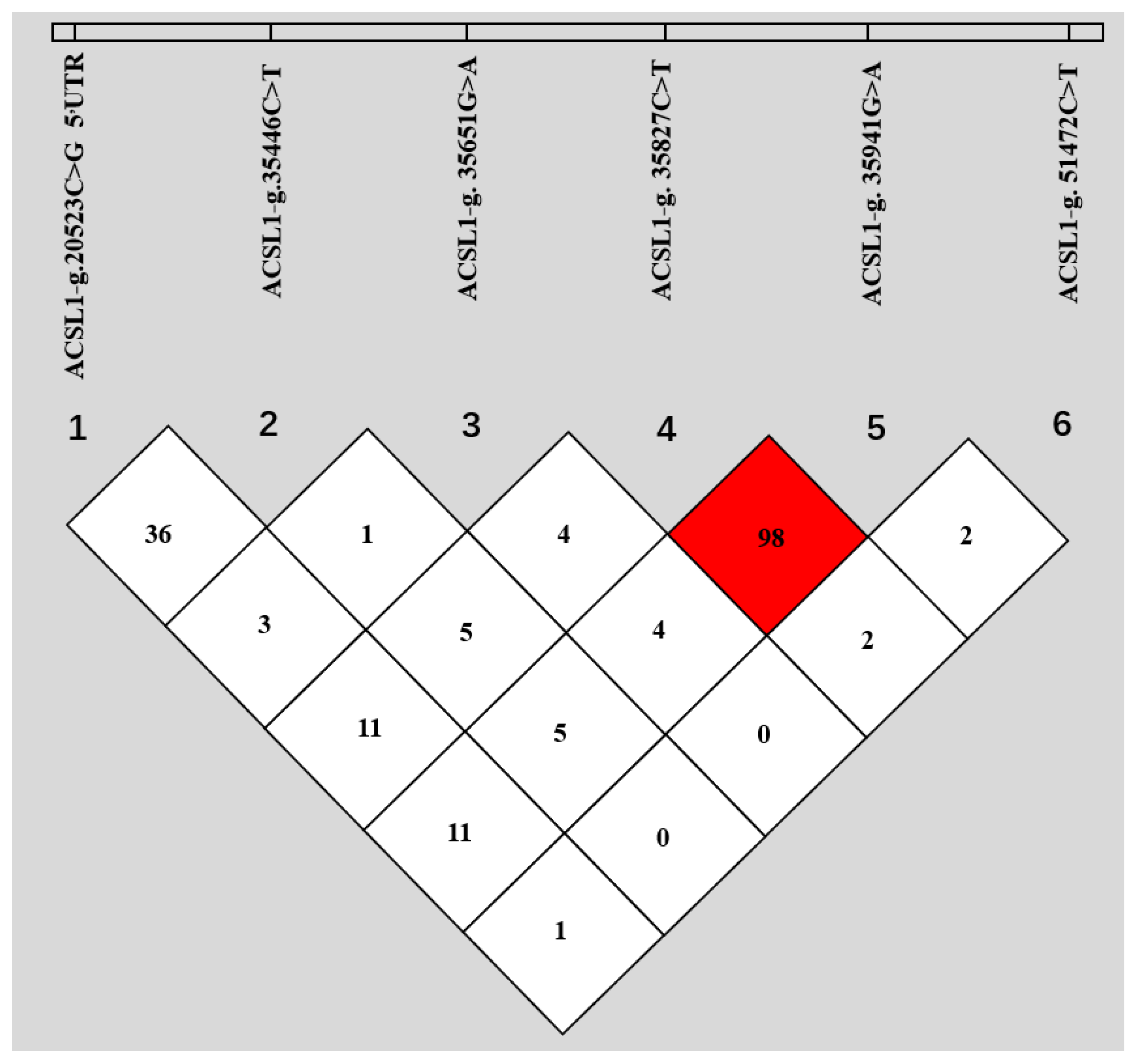
3.1. SNPs within ACSL1
3.2. Effects of Different Non-Genetical Factors on Milking Traits and SCS of Holstein Cows
3.3. Associations of SNPs in ACSL1 with Milking Traits and SCS
3.4. Associations of Haplotypes for SNPs in ACSL1 with Milking Traits and SCS
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Oliveira, H.R.; Cant, J.P.; Brito, L.F.; Feitosa, F.L.B.; Chud, T.C.S.; Fonseca, P.A.S.; Jamrozik, J.; Silva, F.F.; Lourenco, D.A.L.; Schenkel, F.S. Genome-wide association for milk production traits and somatic cell score in different lactation stages of Ayrshire, Holstein, and Jersey dairy cattle. J. Dairy Sci. 2019, 102, 8159–8174. [Google Scholar] [CrossRef] [PubMed]
- Jattawa, D.; Koonawootrittriron, S.; Elzo, M.A.; Suwanasopee, T. Somatic cells count and its genetic association with milk yield in dairy cattle raised under Thai tropical environmental conditions. Asian Australas J. Anim. 2012, 25, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Gussmann, M.; Steeneveld, W.; Kirkeby, C.; Hogeveen, H.; Farre, M.; Halasa, T. Economic and epidemiological impact of different intervention strategies for subclinical and clinical mastitis. Prev. Vet. Med. 2019, 166, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Rupp, R.; Boichard, D. Genetic parameters for clinical mastitis, somatic cell score, production, udder type and milking easy in first lactation Holsteins. J. Dairy Sci. 1999, 82, 2198–2204. [Google Scholar] [CrossRef]
- Akers, R.M. A 100-Year Review: Mammary development and lactation. J. Dairy Sci. 2017, 100, 10332–10352. [Google Scholar] [CrossRef]
- Clancey, E.; Kiser, J.N.; Moraes, J.G.N.; Dalton, J.C.; Spencer, T.E.; Neibergs, H.L. Genome-wide association analysis and gene set enrichment analysis with SNP data identify genes associated with 305-day milk yield in Holstein dairy cows. Anim. Genet. 2019, 50, 254–258. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine mammary protein synthesis during the lactation cycle. Bioinform. Biol. Insights 2011, 5, 83–98. [Google Scholar] [CrossRef]
- Li, C.; Sun, D.X.; Zhang, S.L.; Wang, S.; Wu, X.P.; Zhang, Q.; Liu, L.; Li, Y.H.; Qiao, L. Genome Wide Association Study Identifies 20 Novel Promising Genes Associated with Milk Fatty Acid Traits in Chinese Holstein. PLoS ONE 2014, 9, e96186. [Google Scholar] [CrossRef]
- Soupene, E.; Dinh, N.P.; Siliakus, M.; Kuypers, F.A. Activity of the acyl-CoA synthetase ACSL6 isoforms: Role of the fatty acid Gate-domains. BMC Biochem. 2010, 11, 18. [Google Scholar] [CrossRef]
- Ellis, J.M.; Li, L.O.; Wu, P.C.; Koves, T.; Ilkayeva, O.; Stevens, R.D.; Watkins, S.M.; Muoio, D.M.; Coleman, R.A. Adipose Acyl-CoA Synthetase-1 Directs Fatty Acids toward β-Oxidation and Is Required for Cold Thermogenesis. Cell Metab. 2010, 12, 64. [Google Scholar] [CrossRef]
- Harris, C.A.; Haas, J.; Streeper, R.S.; Stone, S.J.; Kumari, M.; Yang, K.; Han, X.L.; Brownell, N.; Gross, R.W.; Zechner, R.; et al. DGAT enzymes are required for triacylglycerol synthesis and lipid droplets in adipocytes. J. Lipid Res. 2011, 52, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Bernard, L.; Rouel, J.; Leroux, C.; Ferlay, A.; Faulconnier, Y.; Legrand, P.; Chilliard, Y. Mammary lipid metabolism and milk fatty acid secretion in alpine goats fed vegetable lipids. J. Dairy Sci. 2005, 88, 1478–1489. [Google Scholar] [CrossRef]
- Widmann, P.; Nuernberg, K.; Kuehn, C.; Weikard, R. Association of an ACSL1 gene variant with polyunsaturated fatty acids in bovine skeletal muscle. BMC Genet. 2011, 12, 96. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.S.; Pang, C.Y.; Deng, T.X.; Ma, X.Y.; Lu, X.R.; Duan, A.Q.; Liang, X.W. Expression of ACSL1 and Its Effect on Expression Involved in Fatty Acid Metabolism in Buffalo. Chin. J. Anim. Sci. 2020, 56, 41–45. [Google Scholar] [CrossRef]
- Winfrey, M.R.; Rott, M.A.; Wortman, A. Unraveling DNA: Molecular Biology for the Laboratory, 1st ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1997; pp. 234–248. [Google Scholar]
- Hill, W.G.; Robertson, A. Linkage disequilibrium in finite populations. Theor. Appl. Genet. 1968, 38, 226–231. [Google Scholar] [CrossRef]
- Shi, Y.Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetics association at polymorphism loci. Cell Res. 2005, 15, 97–98. [Google Scholar] [CrossRef]
- Browning, B.L.; Zhou, Y.; Browning, S.R. A one-penny imputed genome from next generation reference panels. Am. J. Hum. Genet. 2018, 103, 338–348. [Google Scholar] [CrossRef]
- Mao, Y.J.; Zhu, X.R.; Xing, S.Y.; Zhang, M.R.; Zhang, H.M.; Wang, X.L.; Karrow, N.; Yang, L.G.; Yang, Z.P. Polymorphisms in the promoter region of the bovine lactoferrin gene influence milk somatic cell score and milk production traits in Chinese Holstein cows. Res. Vet. Sci. 2015, 103, 107–112. [Google Scholar] [CrossRef]
- Muoio, D.M.; Lewin, T.M.; Wiedmer, P.; Coleman, R.A. Acyl-CoAs are functionally channeled in liver: Potential role of acyl-CoA synthetase. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E1366–E1373. [Google Scholar] [CrossRef]
- Hoashi, S.; Hinenoya, T.; Tanaka, A.; Ohsaki, H.; Mannen, H. Association between fatty acid compositions and genotypes of FABP4 and LXR-α in Japanese black cattle. BMC Genet. 2008, 9, 84. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. ACSL1, AGPAT6, FABP3, LPINI, and SLC27A6 are the most abundant isoforms in bovine mammary tissue and their expression is affected by stage of lactation. J. Nutr. 2008, 138, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.D.; Tian, H.S.; Jiang, Y.Y.; Shi, B.G.; Liu, X.; Li, X.P.; Wang, D.Z.; Chen, J.L.; Hu, J. Association analysis of ACSL1 gene promoter polymorphism and dairy quality traits in yak. J. Agric. Biol. 2019, 27, 1596–1630. [Google Scholar]
- Zhao, Z.D. Transcriptional Regulation Study of the Bovine ACSL1 Gene. Ph.D. Thesis, Northwest A & F University, Xianyang, China, 2016; pp. 1–2. [Google Scholar]
- Cullen, B.R. Nuclear RNA Export Pathways. Mol. Cell. Biol. 2000, 20, 4181–4187. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.B. Intron-Mediated Regulation of Gene Expression. Curr. Top. Microbiol. 2008, 326, 277–290. [Google Scholar]
- Casas, E.; White, S.N.; Riley, D.G.A.; Smith, T.; Brenneman, R.; Olson, T.A.; Johnson, D.; Coleman, S.; Bennett, G.L.; Chase, C.C. Assessment of single nucleotide polymorphisms in genes residing on chromosomes 14 and 29 for association with carcass composition traits in Bos indicus cattle. J. Anim. Sci. 2005, 83, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Ringnér, M.; Krogh, M. Folding Free Energies of 5’UTRs Impact Post-Transcriptional Regulation on a Genomic Scale in Yeast. PLoS Comput. Biol. 2005, 1, 72. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′→3′) | Size(bp) | Exon | Position | Tm (°C) |
|---|---|---|---|---|---|
| P1 | F1: AACCCAGCGTGACCTGTTTCACCAG | 963 | 5’UTR + Exon 1 | −484~+479 | 69 |
| R1: ATGAGCCTCTGCTCCGTGTGTAACG | |||||
| P2 | F2: GTCCATGCAGCAAACACTCACCC | 1070 | Exon 2,3 + Intron 2,3 | +14,601~+15,648 | 64 |
| R2: CAACCTACAGAGGCTCCAGAAA | |||||
| P3 | F3: ACTGGGCAAGTGTTTTGTTCATTAG | 1034 | Exon 5,6 + Intron 5,6 | +21,364~+22,373 | 63 |
| R3: TCGCTCAGTCATGTCCGACTCTTAG | |||||
| P4 | F4: TCAGCTTGAACTGACTTGATGTGAC | 1382 | Exon 7–9 + Intron 7–9 | +24,637~+25,994 | 64 |
| R4: ATAGTCCGGGCCTAACATGATGGTG | |||||
| P5 | F5: AAGTCCTGCATGGATTTACTTTGTC | 485 | Exon 10 + Intron 10 | +28,070~+28,530 | 63 |
| R5: GAACTGCCTACGGGGAAGATGG | |||||
| P6 | F6: AAGCCACATTCCCTAGTTGCTG | 1028 | Exon 11 + Intron 11 | +30,241~+31,247 | 60 |
| R6: GCACTATGAAAGTGGAGGCATC | |||||
| P7 | F7: TTCCAGTTTCTCCACATCTTCAC | 1291 | Exon 12,13 + Intron 12,13 | +32,505~+33,773 | 59 |
| R7: ACACATCCGAAGAAAAGAAGGG | |||||
| P8 | F8: GGGTCTTTATCCCTTCAGAGGC | 1291 | Exon 19 + 3’UTR | +42,584~+43,853 | 60 |
| R8: CCTATGTCCTAGAATTTTGGCTTG |
| SNP Locus | Location | Number | Genotypes | Genotype Frequency | Allele | Allele Frequency | Chi-Square Value for HWE Test |
|---|---|---|---|---|---|---|---|
| 5’UTR-g.20523C>G | 581 | CC | 0.590 | C | 0.76 | 4.700 | |
| 5’UTR | 334 | CG | 0.339 | G | 0.24 | ||
| 69 | GG | 0.070 | |||||
| g.35446C>T | 770 | CC | 0.780 | C | 0.871 | 34.399 | |
| Intron 2 | 180 | CT | 0.182 | T | 0.129 | ||
| 37 | TT | 0.037 | |||||
| g.35651G>A | 12 | GG | 0.012 | G | 0.107 | 0.042 * | |
| Intron 2 | 188 | GA | 0.190 | A | 0.893 | ||
| 787 | AA | 0.797 | |||||
| g.35827C>T | 540 | CC | 0.548 | C | 0.729 | ||
| Intron 2 | 357 | CT | 0.362 | T | 0.271 | 7.00 | |
| 89 | TT | 0.090 | |||||
| g.35941G>A | 525 | GG | 0.542 | G | 0.727 | ||
| Intron 2 | 357 | GA | 0.369 | A | 0.273 | 4.93 | |
| 87 | AA | 0.089 | |||||
| g.51472C>T | 838 | CC | 0.852 | C | 0.922 | ||
| Intron 11 | 139 | CT | 0.141 | T | 0.078 | 0.219 | |
| 7 | TT | 0.007 |
| Haplotypes | Number | Frequencies |
|---|---|---|
| CCGCGC | 598 | 0.31 |
| CCGTAC | 517 | 0.268 |
| GTGCGC | 230 | 0.119 |
| GCGCGC | 226 | 0.117 |
| CCACGC | 206 | 0.107 |
| CCGCGT | 116 | 0.06 |
| CTGCGT | 19 | 0.01 |
| CCGTAC | 8 | 0.004 |
| GCGCGT | 5 | 0.003 |
| CCACAC | 2 | 0.001 |
| CCACGT | 1 | 0.001 |
| CTGCGC | 1 | 0.001 |
| GCGCAC | 1 | 0.001 |
| GTGCAC | 1 | 0.001 |
| GTGCGT | 1 | 0.001 |
| Total | 1932 | 1 |
| Factors | Milking Traits and SCS | F Value | Sig |
|---|---|---|---|
| Test year | TDMY | 9.63 ** | 0.00 |
| FC | 4.39 * | 0.01 | |
| PC | 9.02 ** | 0.00 | |
| SCS | 9.67 ** | 0.00 | |
| Test season | TDMY | 69.27 ** | 0.00 |
| FC | 99.59 ** | 0.00 | |
| PC | 159.50 ** | 0.00 | |
| SCS | 7.26 ** | 0.00 | |
| Parity | TDMY | 188.99 ** | 0.00 |
| FC | 0.52 | 0.60 | |
| PC | 10.80 ** | 0.00 | |
| SCS | 83.49 ** | 0.00 | |
| Calving season | TDMY | 23.83 ** | 0.00 |
| FC | 5.50 ** | 0.00 | |
| PC | 6.94 ** | 0.00 | |
| SCS | 8.40 ** | 0.00 | |
| Days in milk | TDMY | 471.99 ** | 0.00 |
| FC | 97.49 ** | 0.00 | |
| PC | 745.58 ** | 0.00 | |
| SCS | 35.48 ** | 0.00 | |
| Farm | TDMY | 49.75 ** | 0.00 |
| FC | 65.90 ** | 0.00 | |
| PC | 187.06 ** | 0.00 | |
| SCS | 85.29 ** | 0.00 |
| SNP Locus | Genotypes | DHI Record Number | TDMY (kg) | FC (%) | PC (%) | SCS |
|---|---|---|---|---|---|---|
| 5’UTR-ACSL1-g.20523C>G | CC | 5070 | 34.65 ± 0.15 | 3.63 ± 0.01 | 3.22 ± 0.01 b | 2.84 ± 0.03 a |
| CG | 3204 | 35.40 ± 0.19 | 3.66 ± 0.02 | 3.25 ± 0.01 a | 2.68 ± 0.04 b | |
| GG | 774 | 35.70 ± 0.41 | 3.62 ± 0.03 | 3.27 ± 0.01a | 2.57 ± 0.07 b | |
| Total | 9048 | 34.97 ± 0.11 | 3.64 ± 0.01 | 3.24 ± 0.00 | 2.76 ± 0.02 | |
| F value | 2.789 | 1.734 | 14.797 ** | 4.463 ** | ||
| Sig | 0.06 | 0.18 | 0.00 | 0.01 | ||
| ACSL1-g.35446C>T | CC | 6898 | 34.82 ± 0.13 b | 3.63 ± 0.01 a | 3.23 ± 0.00 b | 2.79 ± 0.03 a |
| CT | 1768 | 35.42 ± 0.26 a,b | 3.70 ± 0.02 a | 3.24 ± 0.01 b | 2.66 ± 0.05 a,b | |
| TT | 410 | 35.84 ± 0.52 a | 3.53 ± 0.05 b | 3.28 ± 0.02 a | 2.75 ± 0.10 b | |
| Total | 9076 | 34.97 ± 0.11 | 3.64 ± 0.01 | 3.24 ± 0.00 | 2.76 ± 0.02 | |
| F value | 4.220 * | 5.002 ** | 6.279 ** | 8.532 ** | ||
| Sig | 0.02 | 0.01 | 0.00 | 0.00 | ||
| ACSL1-g.35651G>A | AA | 71 | 35.01 ± 1.43 | 3.75 ± 0.11 | 3.22 ± 0.04 b | 2.62 ± 0.25 |
| GA | 1660 | 34.45 ± 0.27 | 3.64 ± 0.02 | 3.22 ± 0.01 b | 2.78 ± 0.05 | |
| GG | 7338 | 35.08 ± 0.13 | 3.64 ± 0.01 | 3.24 ± 0.00 a | 2.76 ± 0.02 | |
| Total | 9069 | 34.97 ± 0.11 | 3.64 ± 0.01 | 3.24 ± 0.00 | 2.76 ± 0.02 | |
| F value | 1.262 | 0.704 | 7.016 ** | 0.007 | ||
| Sig | 0.28 | 0.50 | 0.00 | 0.99 | ||
| ACSL1-g.35827C>T | CC | 5143 | 35.19 ± 0.15 a | 3.67 ± 0.01 a | 3.25 ± 0.01 a | 2.83 ± 0.03 a |
| TC | 3180 | 35.03 ± 0.19 a | 3.62 ± 0.02 a | 3.22 ± 0.01 b | 2.67 ± 0.04 b | |
| TT | 740 | 33.24 ± 0.38 b | 3.56 ± 0.03 b | 3.22 ± 0.01 b | 2.67 ± 0.08 b | |
| Total | 9063 | 34.97 ± 0.11 | 3.64 ± 0.01 | 3.24 ± 0.00 | 2.76 ± 0.02 | |
| F value | 4.869 ** | 5.301 ** | 3.984 * | 14.045 ** | ||
| Sig | 0.01 | 0.01 | 0.02 | 0.00 | ||
| ACSL1-g.51472C>T | CC | 7592 | 35.04 ± 0.13 b | 3.63 ± 0.01 b | 3.23 ± 0.00 | 2.78 ± 0.02 a |
| TC | 1376 | 34.56 ± 0.28 b | 3.70 ± 0.02 b | 3.27 ± 0.01 | 2.66 ± 0.06 b | |
| TT | 76 | 38.69 ± 1.23 a | 3.92 ± 0.10 a | 3.26 ± 0.04 | 2.81 ± 0.22 a,b | |
| Total | 9044 | 34.97 ± 0.11 | 3.64 ± 0.01 | 3.24 ± 0.00 | 2.76 ± 0.02 | |
| F value | 3.169 * | 4.359 * | 0.371 | 3.412 * | ||
| Sig | 0.04 | 0.01 | 0.69 | 0.03 |
| Haplotypes | DHI Records Number | TDMY (kg) | FC (%) | PC (%) | SCS |
|---|---|---|---|---|---|
| CCGCGC | 4116 | 34.89 ± 0.17 b,c | 3.64 ± 0.01 b,c | 3.23 ± 0.01 b | 2.91 ± 0.03 a,b |
| GCGCGC | 1335 | 35.56 ± 0.30 a,b,c | 3.69 ± 0.02 b,c | 3.26 ± 0.01 b | 2.48 ± 0.05 a,b,c,d |
| CCGCAC | 1182 | 35.63 ± 0.30 a,b,c | 3.59 ± 0.03 b,c | 3.21 ± 0.01 b | 2.69 ± 0.06 a,b,c |
| CCGTAC | 1133 | 34.05 ± 0.32 b,c | 3.59 ± 0.03 b,c | 3.22 ± 0.01 b | 2.73 ± 0.06 a,b,c |
| GTGCGC | 355 | 36.10 ± 0.56 a,b | 3.49 ± 0.05 c | 3.29 ± 0.02 ab | 2.58 ± 0.10 a,b,c,d |
| CCGTGC | 354 | 33.77 ± 0.51 b,c | 3.67 ± 0.05 b,c | 3.24 ± 0.02 b | 2.65 ± 0.12 a,b,c |
| CCACGC | 225 | 34.12 ± 0.73 b,c | 3.71 ± 0.06 b,c | 3.26 ± 0.02 b | 3.00 ± 0.17 a |
| CTGCGC | 198 | 34.27 ± 0.71 b,c | 3.88 ± 0.06 b | 3.24 ± 0.03 b | 3.07 ± 0.16 a |
| CCGCGT | 85 | 38.91 ± 1.11 a | 3.81 ± 0.10 b | 3.26 ± 0.04 b | 2.64 ± 0.20 a,b,c |
| GCGTGC | 61 | 32.31 ± 1.49 c | 3.61 ± 0.10 b,c | 3.30 ± 0.03 a,b | 1.92 ± 0.20 d |
| GCGCAC | 55 | 33.41 ± 1.15 b,c | 4.15 ± 0.10 a | 3.38 ± 0.05 a | 2.17 ± 0.27 c,d |
| Total | 9099 | 34.97 ± 0.11 | 3.64 ± 0.01 | 3.24 ± 0.00 | 2.76 ± 0.02 |
| F value | 3.979 ** | 5.136 ** | 3.201 ** | 6.749 ** | |
| Sig | 0.00 | 0.00 | 0.00 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; Gao, Q.; Zhang, Q.; Arbab, A.A.I.; Li, M.; Yang, Z.; Karrow, N.A.; Mao, Y. Polymorphisms of the ACSL1 Gene Influence Milk Production Traits and Somatic Cell Score in Chinese Holstein Cows. Animals 2020, 10, 2282. https://doi.org/10.3390/ani10122282
Liang Y, Gao Q, Zhang Q, Arbab AAI, Li M, Yang Z, Karrow NA, Mao Y. Polymorphisms of the ACSL1 Gene Influence Milk Production Traits and Somatic Cell Score in Chinese Holstein Cows. Animals. 2020; 10(12):2282. https://doi.org/10.3390/ani10122282
Chicago/Turabian StyleLiang, Yan, Qisong Gao, Qiang Zhang, Abdelaziz Adam Idriss Arbab, Mingxun Li, Zhangping Yang, Niel A. Karrow, and Yongjiang Mao. 2020. "Polymorphisms of the ACSL1 Gene Influence Milk Production Traits and Somatic Cell Score in Chinese Holstein Cows" Animals 10, no. 12: 2282. https://doi.org/10.3390/ani10122282
APA StyleLiang, Y., Gao, Q., Zhang, Q., Arbab, A. A. I., Li, M., Yang, Z., Karrow, N. A., & Mao, Y. (2020). Polymorphisms of the ACSL1 Gene Influence Milk Production Traits and Somatic Cell Score in Chinese Holstein Cows. Animals, 10(12), 2282. https://doi.org/10.3390/ani10122282