Dihydropyridine Enhances the Antioxidant Capacities of Lactating Dairy Cows under Heat Stress Condition


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Statement
2.2. Determination of Anti-Oxidative Indexes
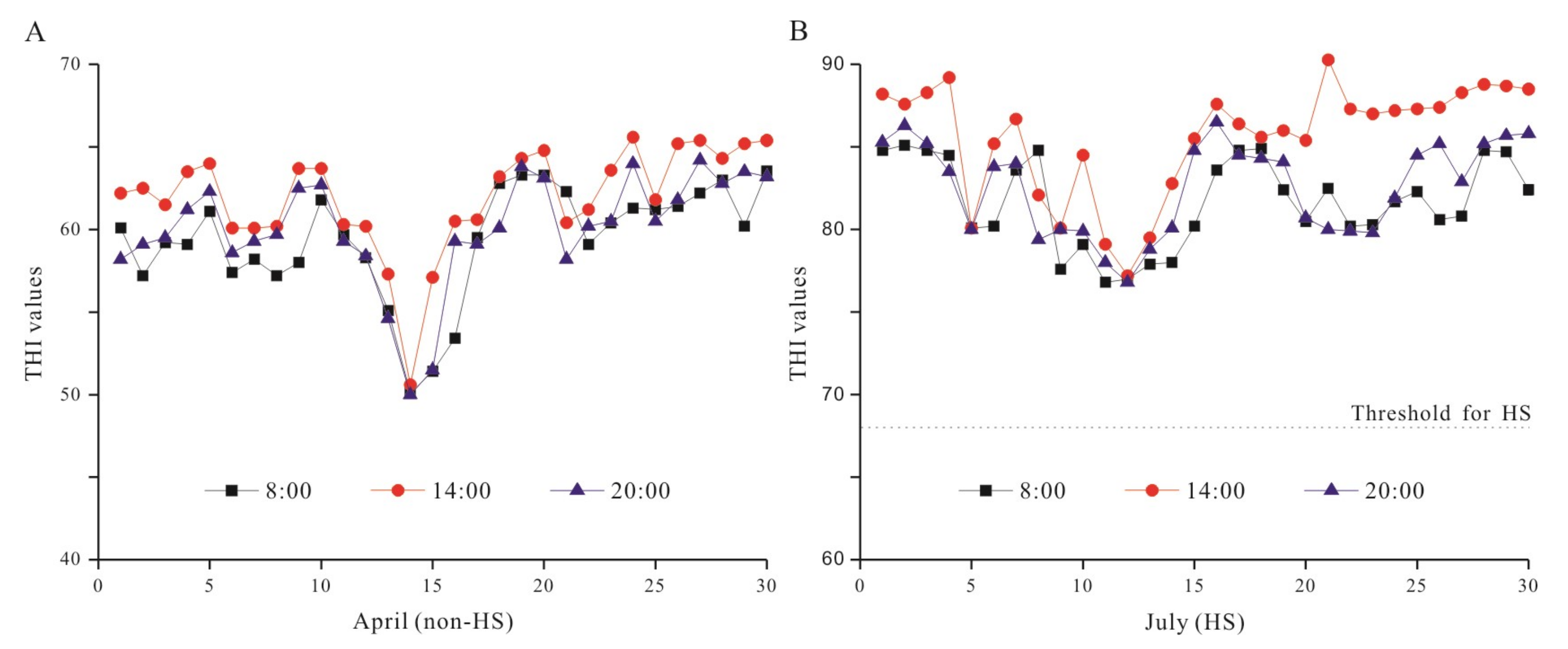
2.3. Detection of HS Using Temperature–Humidity Index (THI) Method
2.4. DHP Supplement Experiment
2.5. Blood Sample Collection
2.6. Ruminal Fluids Collection
2.7. Detection of Antioxidant Status of Dairy Cows
2.8. DNA Extraction
2.9. Real-Time PCR
2.10. Denaturing Gradient Gel Electrophoresis (DGGE)
2.11. DNA Sequencing and Phylogenetic Tree Analysis
2.12. Statistical Analysis
3. Results
3.1. THI Determination in Different Seasons
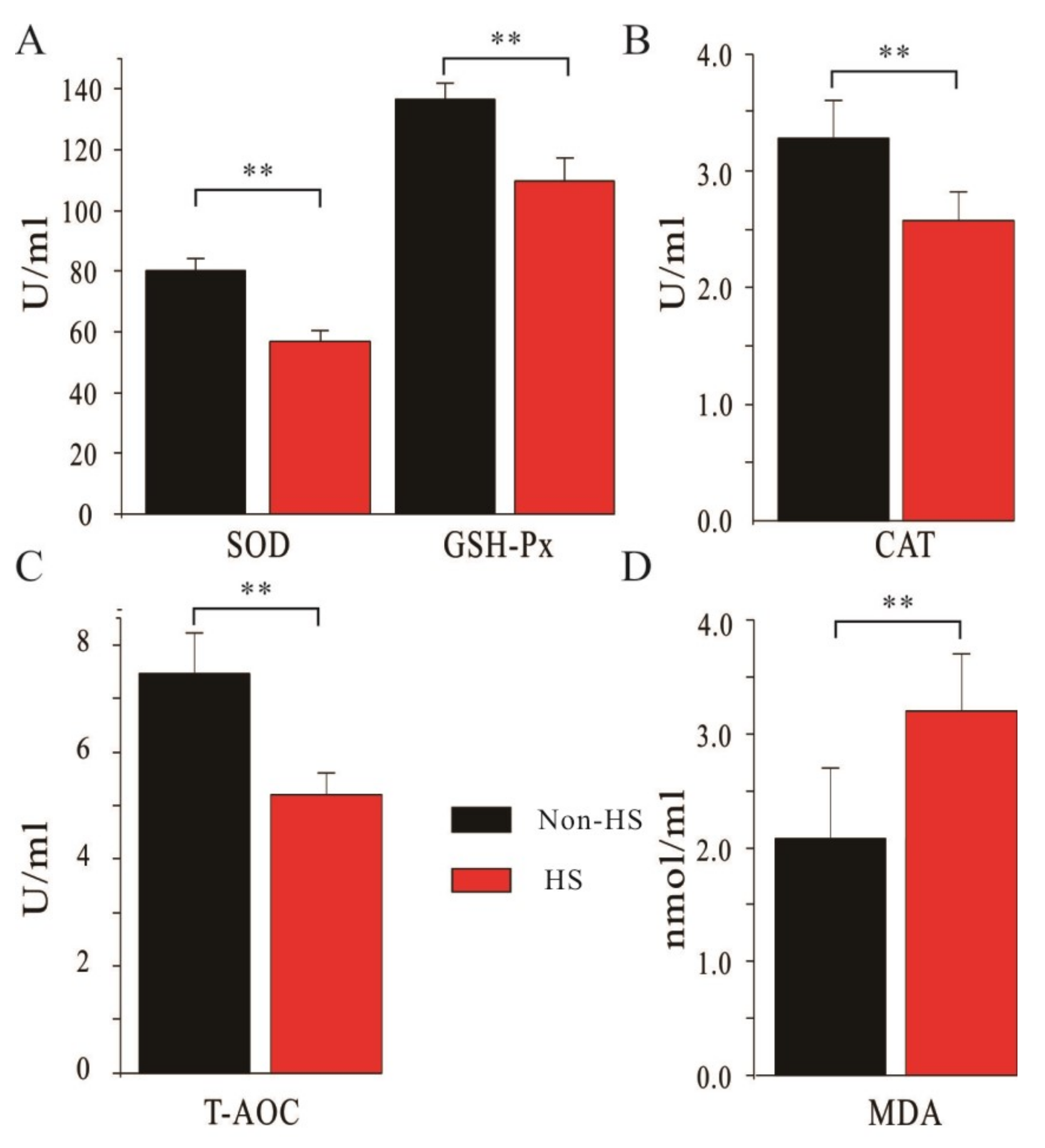
3.2. Changes in Serum Antioxidant Capacity of Dairy Cows in Spring (Non-HS) and Summer (HS)
3.3. Correlation Analysis between the Serum Antioxidant Capacity and Milk Yield
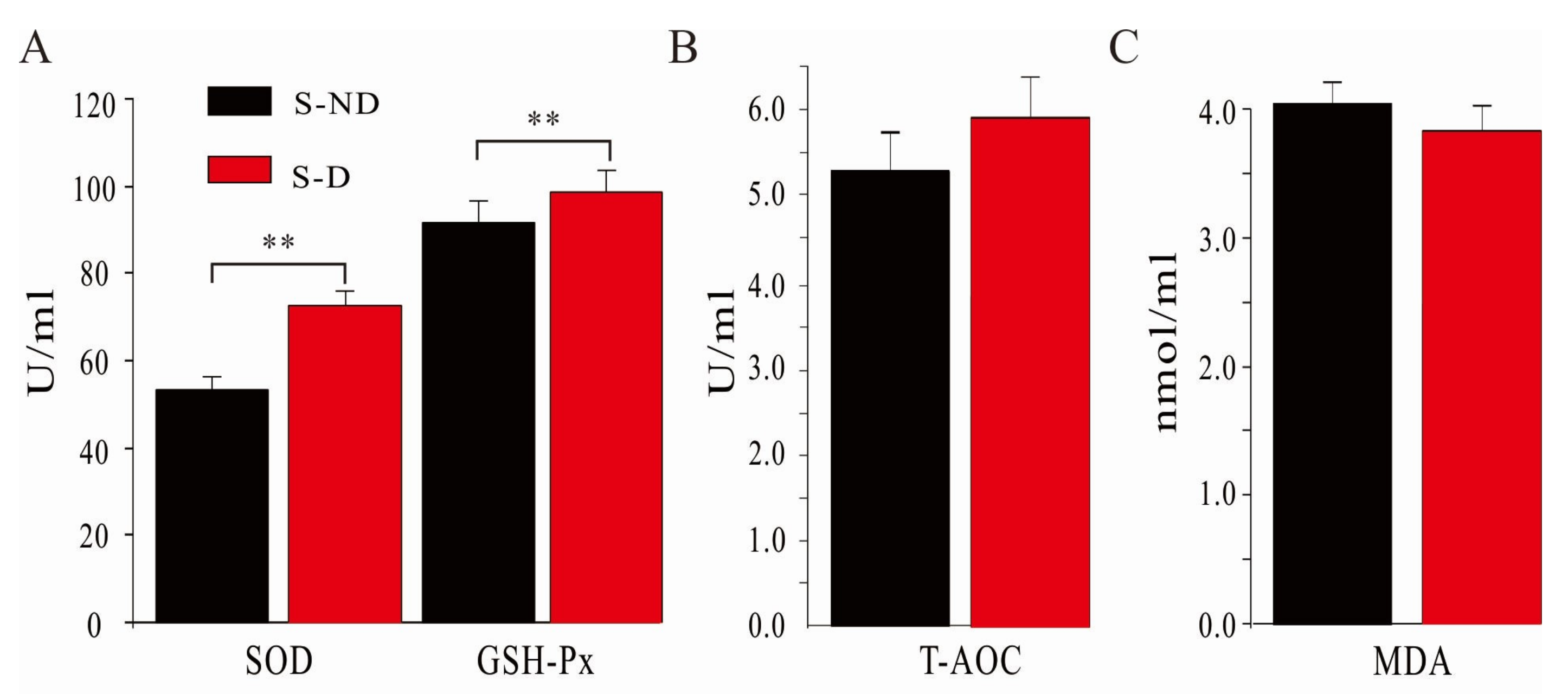
3.4. Effects of DHP on Serum Antioxidant Capacity of Dairy Cows under HS Conditions
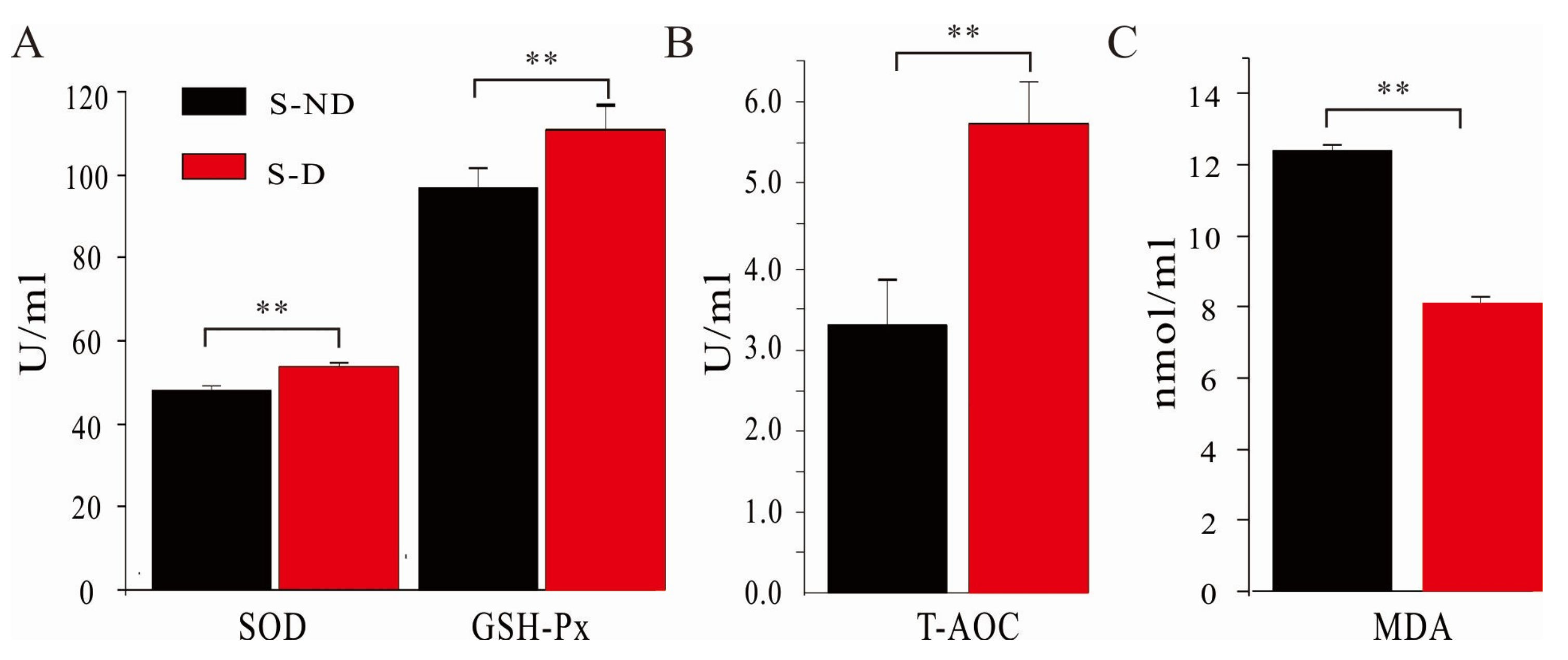
3.5. Effects of DHP on Ruminal Antioxidant Capacity of Dairy Cows under HS Condition
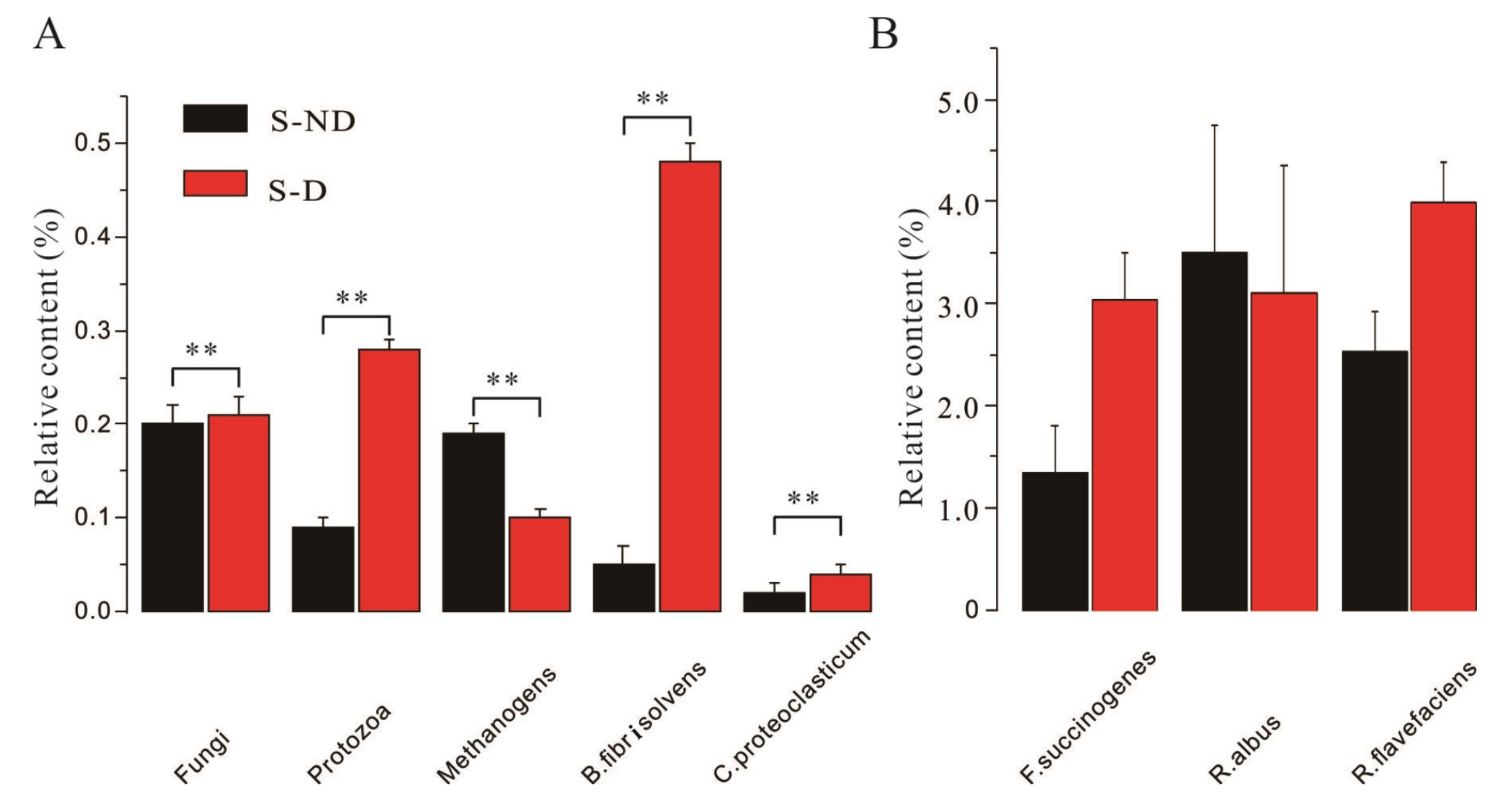
3.6. Effects of DHP on Ruminal Microbial Communities
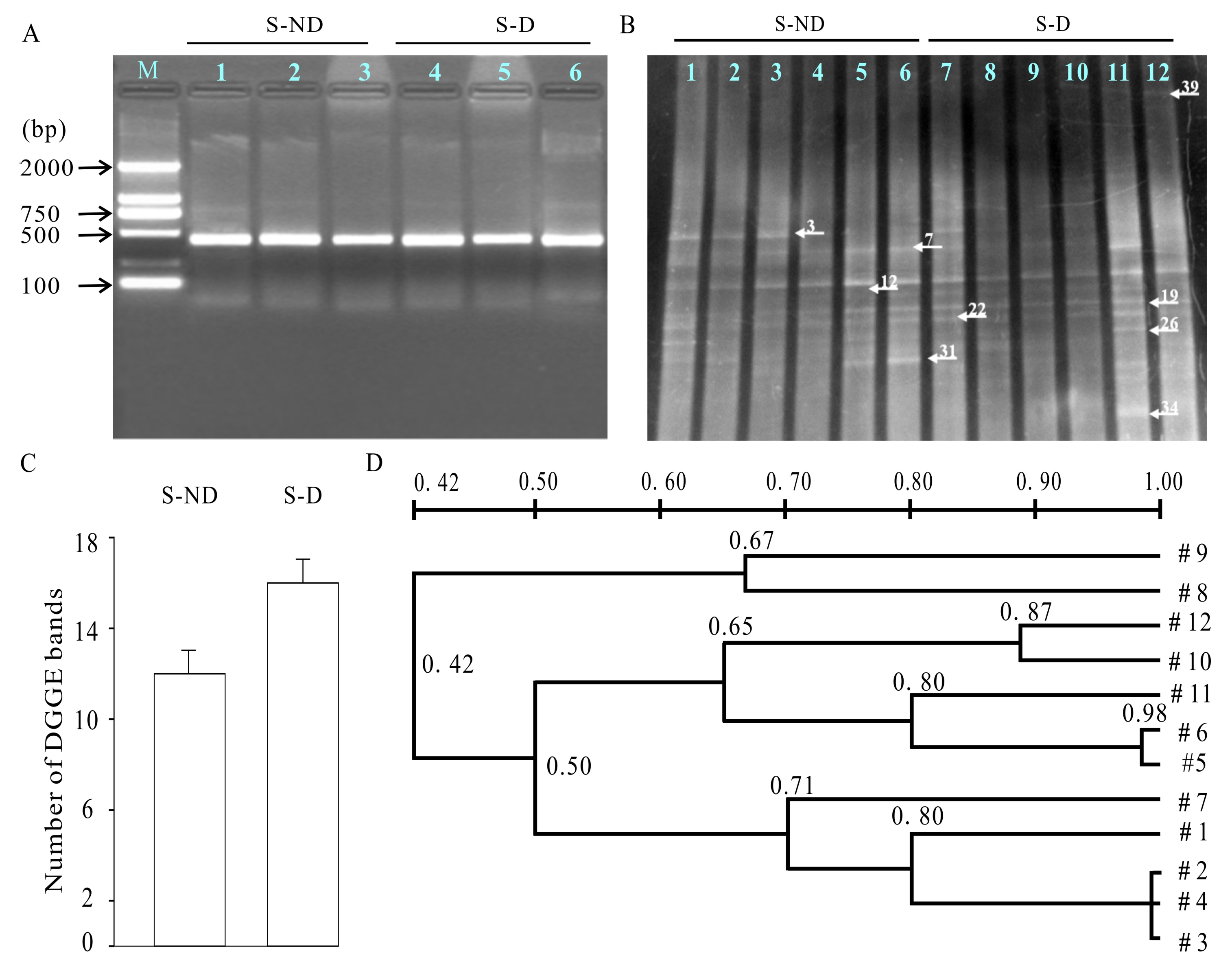
3.7. DGGE Analysis of Microbe Communities in Rumen
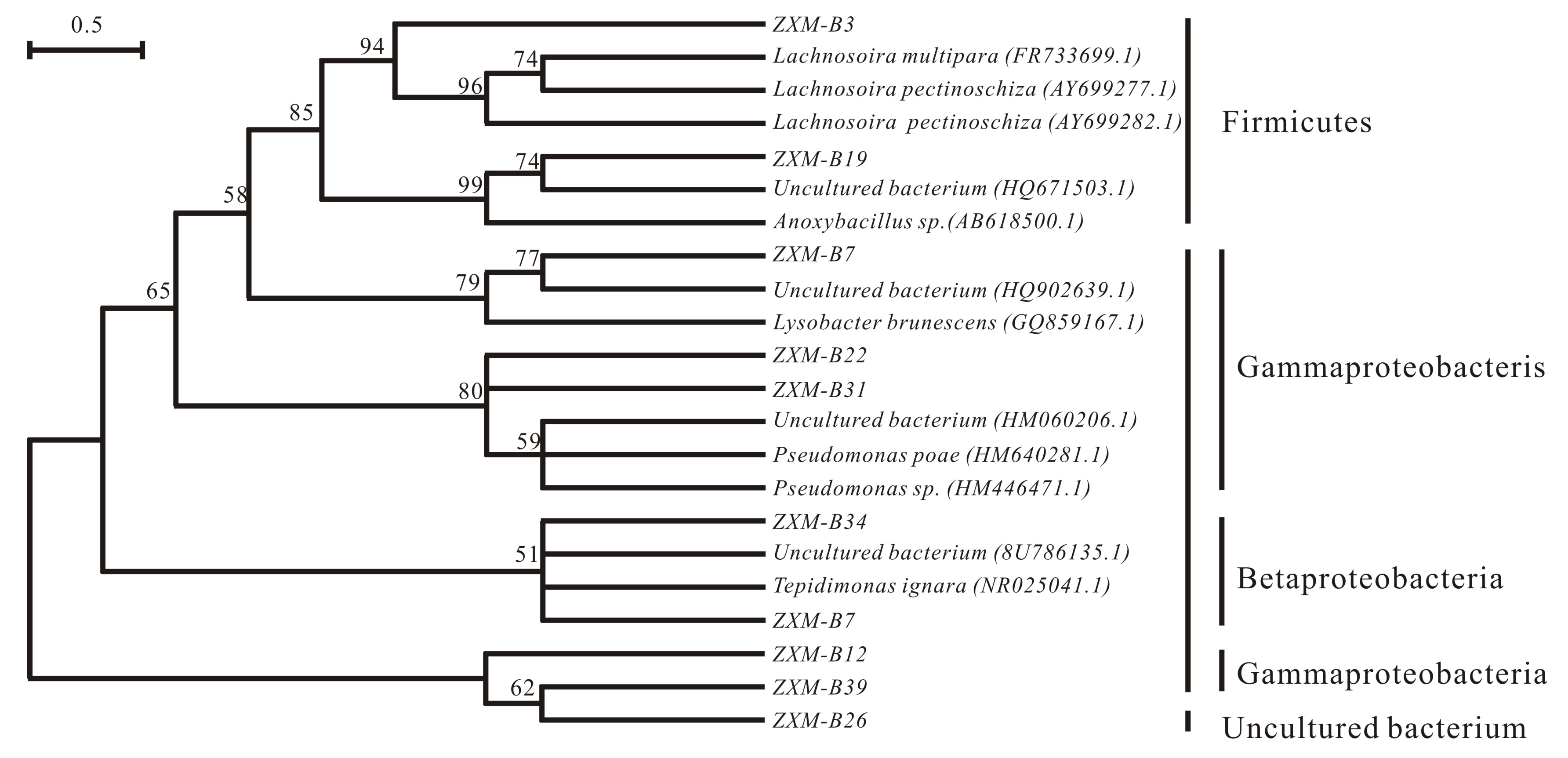
3.8. Phylogenic Tree Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aitken, S.; Karcher, E.; Rezamand, P.; Gandy, J.; VandeHaar, M.; Capuco, A.; Sordillo, L. Evaluation of antioxidant and proinflammatory gene expression in bovine mammary tissue during the periparturient period. J. Dairy Sci. 2009, 92, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Guo, J.; Quan, S.; Nan, X.; Sanz-Fernandez, M.V.; Baumgard, L.; Bu, D. The effects of heat stress on protein metabolism in lactating Holstein cows. J. Dairy Sci. 2017, 100, 5040–5049. [Google Scholar] [CrossRef] [PubMed]
- Zachut, M.; Kra, G.; Livshitz, L.; Portnick, Y.; Yakoby, S.; Friedlander, G.; Levin, Y. Seasonal heat stress affects adipose tissue proteome toward enrichment of the Nrf2-mediated oxidative stress response in late-pregnant dairy cows. J. Proteom. 2017, 158, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Gantner, V.; Bobic, T.; Gantner, R.; Gregic, M.; Kuterovac, K.; Novakovic, J.; Potocnik, K. Differences in response to heat stress due to production level and breed of dairy cows. Int. J. Biometeorol. 2017, 92, 1675–1685. [Google Scholar] [CrossRef] [PubMed]
- Puppel, K.; Kapusta, A.; Kuczyńska, B. The etiology of oxidative stress in the various species of animals, a review. J. Sci. Food Agric. 2014, 95, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Sun, L.; Shu, S.; Zhu, K.; Xu, C.; Wang, J.; Wang, H. Nuclear magnetic resonance-based serum metabolic profiling of dairy cows with footrot. J. Veter-Med Sci. 2016, 78, 1421–1428. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.-C.; Chen, H.-R.; Chiu, C.-C.; Liou, S.-F.; Chen, I.-J.; Yeh, J.-L. Protective effect of labedipinedilol-A, a novel dihydropyridine-type calcium channel blocker, on myocardial apoptosis in ischemia–reperfusion injury. Life Sci. 2006, 79, 1248–1256. [Google Scholar] [CrossRef]
- Wang, J.-G.; Kario, K.; Lau, T.; Wei, Y.Q.; Park, C.G.; Kim, C.H.; Huang, J.; Zhang, W.; Li, Y.; Yan, P.; et al. Use of dihydropyridine calcium channel blockers in the management of hypertension in Eastern Asians: A scientific statement from the Asian Pacific Heart Association. Hypertens. Res. 2011, 34, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Robles, N.R.; Fici, F.; Grassı, G. Dihydropyridine calcium channel blockers and renal disease. Hypertens. Res. 2016, 40, 21–28. [Google Scholar] [CrossRef]
- Wu, G. Important roles of dietary taurine, creatine, carnosine, anserine and 4-hydroxyproline in human nutrition and health. Amino Acids 2020, 52, 329–360. [Google Scholar] [CrossRef]
- Niu, Z.; Liu, F.; Min, Y.; Li, W. Effects of dietary dihydropyridine supplementation on growth performance and lipid metabolism of broiler chickens. Czech J. Anim. Sci. 2010, 55, 116–122. [Google Scholar] [CrossRef]
- Bian, L.Q.; Yuan, D.; Liu, X.J.; Chen, J.; Liao, S.W.; Li, L. Effects of dihydropyridine on the growth performance, meat quality and anti-oxidant properties of muslce tissues in fattening pigs. Henan J. Anim. Husban Vet. Med. 2010, 30, 5–7. [Google Scholar] [CrossRef]
- West, J.; Mullinix, B.; Bernard, J. Effects of Hot, Humid Weather on Milk Temperature, Dry Matter Intake, and Milk Yield of Lactating Dairy Cows. J. Dairy Sci. 2003, 86, 232–242. [Google Scholar] [CrossRef]
- Ministry of Agriculture of China. Veterinary Drug Quality Standards; China Agriculture Press: Beijing, China, 2003; pp. 1–2.
- Guo, J.; Gao, S.; Quan, S.; Zhang, Y.; Bu, D.; Wang, J. Blood amino acids profile responding to heat stress in dairy cows. Asian-Australas. J. Anim. Sci. 2017, 31, 47–53. [Google Scholar] [CrossRef]
- Weimer, P.; Waghorn, G.; Odt, C.; Mertens, D. Effect of Diet on Populations of Three Species of Ruminal Cellulolytic Bacteria in Lactating Dairy Cows. J. Dairy Sci. 1999, 82, 122–134. [Google Scholar] [CrossRef]
- Jin, D.; Zhao, S.; Zheng, N.; Bu, D.; Beckers, Y.; Wang, J. Urea nitrogen induces changes in rumen microbial and host metabolic profiles in dairy cows. Livest. Sci. 2018, 210, 104–110. [Google Scholar] [CrossRef]
- Jafari, S.; Ebrahimi, M.; Meng, G.Y.; Rajion, M.A.; Jahromi, M.F. Dietary supplementation of papaya (Carica papaya l.) leaf affects abundance of Butyrivibrio fibrisolvens and modulates biohydrogenation of C18 polyunsaturated fatty acids in the rumen of goats. Ital. J. Anim. Sci. 2017, 17, 326–335. [Google Scholar] [CrossRef]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef]
- Denman, S.E.; Tomkins, N.W.; McSweeney, C.S. Quantitation and diversity analysis of ruminal methanogenic populations in response to the antimethanogenic compound bromochloromethane. FEMS Microbiol. Ecol. 2007, 62, 313–322. [Google Scholar] [CrossRef]
- Alekkhahi, M.; Tahmasbi, A.; Naserian, A.A.; Mesgaran, M.D.; Kleen, J.; AlZahal, O.; Ghaffari, M. Effects of supplementation of active dried yeast and malate during sub-acute ruminal acidosis on rumen fermentation, microbial population, selected blood metabolites, and milk production in dairy cows. Anim. Feed. Sci. Technol. 2016, 213, 29–43. [Google Scholar] [CrossRef]
- Zhang, T.; Kucharzyk, K.H.; Kim, B.; Deshusses, M.A.; Hsu-Kim, H. Net Methylation of Mercury in Estuarine Sediment Microcosms Amended with Dissolved, Nanoparticulate, and Microparticulate Mercuric Sulfides. Environ. Sci. Technol. 2014, 48, 9133–9141. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Kamra, D.N.; Chatterjee, P.N.; Kumar, R.; Chaudhary, L.C. In vitro Methanogenesis, Microbial Profile and Fermentation of Green Forages with Buffalo Rumen Liquor as Influenced by 2-Bromoethanesulphonic Acid. Asian-Australas. J. Anim. Sci. 2008, 21, 818–823. [Google Scholar] [CrossRef]
- Ma, L.; He, F.; Huang, T.; Zhou, Q.; Zhang, Y.; Wu, Z. Nitrogen and phosphorus transformations and balance in a pond-ditch circulation system for rural polluted water treatment. Ecol. Eng. 2016, 94, 117–126. [Google Scholar] [CrossRef]
- Wang, W.; Li, C.; Li, F.; Wang, X.; Zhang, X.; Liu, T.; Nian, F.; Yue, X.; Li, F.; Pan, X.; et al. Effects of early feeding on the host rumen transcriptome and bacterial diversity in lambs. Sci. Rep. 2016, 6, 32479. [Google Scholar] [CrossRef] [PubMed]
- Zimbelman, R.; Rhoads, R.; Rhoads, M.; Duff, G.; Baumguard, L.; Collier, R. A re-evaluation of the impact of temperature humidity index (THI) and black globe humidity index (BGHI) on milk production in high producing dairy cows. In Proceedings of the 24th Annu. Southwest Nutrition Management Conference, Tempe, AZ, USA, 26–27 February 2009; pp. 158–169. [Google Scholar]
- Ravagnolo, O.; Misztal, I.; Hoogenboom, G. Genetic Component of Heat Stress in Dairy Cattle, Development of Heat Index Function. J. Dairy Sci. 2000, 83, 2120–2125. [Google Scholar] [CrossRef]
- Wang, Y.-M. Effects of Diatery Fat and Engergy Density on Oxidative Stress and Lactation Performance in Dairy Cows, in the Absence or Presence of Antioxidant. Ph.D Thesis, Zhejiang University, Hangzhou, China, 2010. [Google Scholar]
- Gobert, M.; Gruffat, D.; Habeanu, M.; Parafita, E.; Bauchart, D.; Durand, D. Plant extracts combined with vitamin E in PUFA-rich diets of cull cows protect processed beef against lipid oxidation. Meat Sci. 2010, 85, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Anon, M.; Nocek, J.; Bowman, G.; Hampton, T.; Atwell, C.; Vázquez, P.; Jenkins, T. Effects of Feeding a Dietary Antioxidant in Diets with Oxidized Fat on Lactation Performance and Antioxidant Status of the Cow. J. Dairy Sci. 2008, 91, 3165–3172. [Google Scholar] [CrossRef] [PubMed]
- Polsky, L.; Von Keyserlingk, M. Invited review: Effects of heat stress on dairy cattle welfare. J. Dairy Sci. 2017, 100, 8645–8657. [Google Scholar] [CrossRef]
- Miron, J.; Ben-Ghedalia, D.; Morrison, M. Invited Review: Adhesion Mechanisms of Rumen Cellulolytic Bacteria. J. Dairy Sci. 2001, 84, 1294–1309. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Components | Formulation (%) | Nutritional Ingredients | Content (%) |
|---|---|---|---|
| Corn silage | 55.6 | Dry matter (DM, %) | 51.4 |
| Peanut hay | 13.9 | Dry matter intake (DMI, kg/d) | 18.5 |
| Corn | 15.6 | Net energy for lactation (NEL, MJ/d) | 29.7 |
| Wheat bran | 6.1 | Crude protein (CP, %) | 15.37 |
| Soybean meal | 4.6 | Acid detergent fiber (ADF, %) | 24.09 |
| Cotton seed meal | 2.4 | Neutral detergent fiber (NDF, %) | 37.42 |
| CaHPO4 | 0.6 | Calcium (Ca, %) | 0.91 |
| NaHCO3 | 0.3 | Phosphorus (P, %) | 0.48 |
| Premix | 0.9 | ||
| Total | 100.0 |
| Target Species | Primer | Sequence (5′-3′) | Reference |
|---|---|---|---|
| Total bacteria | F R | CGGCAACGAGCGCAACC CCATTGTAGCACGTGTGTAGCC | [19] |
| Fungi | F R | GAGGAAGTAAAAGTCGTAACAAGGTTTC CAAATTCACAAAGGGTAGGATGATT | [19] |
| Protozoa | F R | GCTTTCGWTGGTAGTGTATT CTTGCCCTCYAATCGTWCT | [20] |
| Methanogens | F R | TTCGGTGGATCDCARAGRGC GBARGTCGWAWCCGTAGAATCC | [20] |
| B. fibrisolvens | F R | ACACACCGCCCGTCACA TCCTTACGGTTGGGTCACAGA | |
| C.proteoclasticum | F R | TCCGGTGGTATGAGATGGGC GTCGCTGCATCAGAGTTTCCT | |
| F. succinogenes | F R | GTTCGGAATTACTGGGCGTAAA CGCCTGCCCCTGAACTATC | [21] |
| R. albus | F R | CGGCAACGAGCGCAACCC CCATTGTAGCACGTGTGTAGCC | [22] |
| R. flavefaciens | F R | CGAACGGAGATAATTTGAGTTTACTTAGG CGGTCTCTGTATGTTATGAGGTATTACC | [23] |
| Items | Milk Yield Decrease | |
|---|---|---|
| Correlation Coefficient (r Values) | p Values | |
| SOD (U/mL) | −0.462 | p < 0.01 |
| GSH-Px (U/mL) | −0.619 | p < 0.01 |
| CAT (U/mL) | −0.291 | p > 0.05 |
| T-AOC (U/mL) | −0.515 | p < 0.01 |
| MDA (nmol/mL) | 0.371 | p < 0.05 |
| Lane | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 100.0 | 80.1 | 79.8 | 80.3 | 69.2 | 70.8 | 60.5 | 40.1 | 20.0 | 19.2 | 52.9 | 32.7 |
| 2 | 80.1 | 100.0 | 99.3 | 99.3 | 68.2 | 68.4 | 74.8 | 50.1 | 24.9 | 24.0 | 62.4 | 40.3 |
| 3 | 79.8 | 99.3 | 100.0 | 99.3 | 68.3 | 68.5 | 74.2 | 50.0 | 24.9 | 24.0 | 62.5 | 40.4 |
| 4 | 80.3 | 99.3 | 99.3 | 100.0 | 68.1 | 68.3 | 74.7 | 49.6 | 24.8 | 23.9 | 62.3 | 40.2 |
| 5 | 69.2 | 68.2 | 68.3 | 68.1 | 100.0 | 98.4 | 50.8 | 50.6 | 34.3 | 55.1 | 80.4 | 67.6 |
| 6 | 70.8 | 68.4 | 68.5 | 68.3 | 98.4 | 100.0 | 50.9 | 50.7 | 34.3 | 55.0 | 80.3 | 67.5 |
| 7 | 60.5 | 74.8 | 74.2 | 74.7 | 50.8 | 50.9 | 100.0 | 49.8 | 24.8 | 23.9 | 46.4 | 40.2 |
| 8 | 40.1 | 50.1 | 50.0 | 49.6 | 50.6 | 50.7 | 49.8 | 100.0 | 67.2 | 47.7 | 47.1 | 41.0 |
| 9 | 20.0 | 24.9 | 24.9 | 24.8 | 34.3 | 34.3 | 24.8 | 67.2 | 100.0 | 72.1 | 46.9 | 61.6 |
| 10 | 19.2 | 24.0 | 24.0 | 23.9 | 55.1 | 55.0 | 23.9 | 47.7 | 72.1 | 100.0 | 66.6 | 87.2 |
| 11 | 52.9 | 62.4 | 62.5 | 62.3 | 80.4 | 80.3 | 46.4 | 47.1 | 46.9 | 66.6 | 100.0 | 77.4 |
| 12 | 32.7 | 40.3 | 40.4 | 40.2 | 67.6 | 67.5 | 40.2 | 41.0 | 61.6 | 87.2 | 77.4 | 100.0 |
| Bands | Closest Known Species | Sequence Similarity | Length (bp) | Taxonomic Group | Acc. Number |
|---|---|---|---|---|---|
| B3 | Lachnospira pectinoschiza AY699277.1 | 99% | 433 | Bacteria; Firmicutes; Clostridia; Clostridiales; Lachnospiraceae; Lachnospira. | JF798509 |
| B7 | Lysobacter brunescens GQ859167.1 | 99% | 434 | Bacteria; Proteobacteria; Gammaproteobacteria; Xanthomonadales; Xanthomonadaceae; Lysobacter. | JF798510 |
| B12 | Acinetobacter sp. JFAN2 HQ693555.1 | 99% | 435 | Bacteria; Proteobacteria; Gammaproteobacteria; Pseudomonadales; Moraxellaceae; Acinetobacter. | JF798511 |
| B19 | Anoxybacillus sp. IP-3 AB618500.1 | 99% | 436 | Bacteria; Firmicutes; Bacillales; Bacillaceae; Anoxybacillus. | JF798512 |
| B22 | Pseudomonas sp. SUT 19 HM446471.1 | 99% | 434 | Bacteria; Proteobacteria; Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas. | JF798513 |
| B26 | Uncultured bacterium EU843886.1 | 99% | 437 | Bacteria; environmental samples | JF798514 |
| B31 | Pseudomonas sp. SUT 19 HM446471.1 | 99% | 434 | Bacteria; Proteobacteria; Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas. | JF798515 |
| B34 | Tepidimonas aquatica NR_025755.1 | 99% | 430 | Bacteria; Proteobacteria; Betaproteobacteria; Burkholderiales; Tepidimonas. | JF798516 |
| B39 | Xanthomonas axonopodis AB101447.1 | 99% | 433 | Bacteria; Proteobacteria; Gammaproteobacteria; Xanthomonadales; Xanthomonadaceae; Xanthomonas. | JF798517 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.-F.; Zhao, X.-M.; Cai, H.; Yi, J.-M.; Hua, G.-H. Dihydropyridine Enhances the Antioxidant Capacities of Lactating Dairy Cows under Heat Stress Condition. Animals 2020, 10, 1812. https://doi.org/10.3390/ani10101812
Yu M-F, Zhao X-M, Cai H, Yi J-M, Hua G-H. Dihydropyridine Enhances the Antioxidant Capacities of Lactating Dairy Cows under Heat Stress Condition. Animals. 2020; 10(10):1812. https://doi.org/10.3390/ani10101812
Chicago/Turabian StyleYu, Meng-Fei, Xin-Mao Zhao, Hang Cai, Jian-Ming Yi, and Guo-Hua Hua. 2020. "Dihydropyridine Enhances the Antioxidant Capacities of Lactating Dairy Cows under Heat Stress Condition" Animals 10, no. 10: 1812. https://doi.org/10.3390/ani10101812
APA StyleYu, M.-F., Zhao, X.-M., Cai, H., Yi, J.-M., & Hua, G.-H. (2020). Dihydropyridine Enhances the Antioxidant Capacities of Lactating Dairy Cows under Heat Stress Condition. Animals, 10(10), 1812. https://doi.org/10.3390/ani10101812