
Sustainable Production of N-methylphenylalanine by Reductive Methylamination of Phenylpyruvate Using Engineered Corynebacterium glutamicum




Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Molecular Genetic Techniques and Strain Construction
2.3. Protein Analytics
2.4. Quantification of Amino Acids and Organic Acids
3. Results
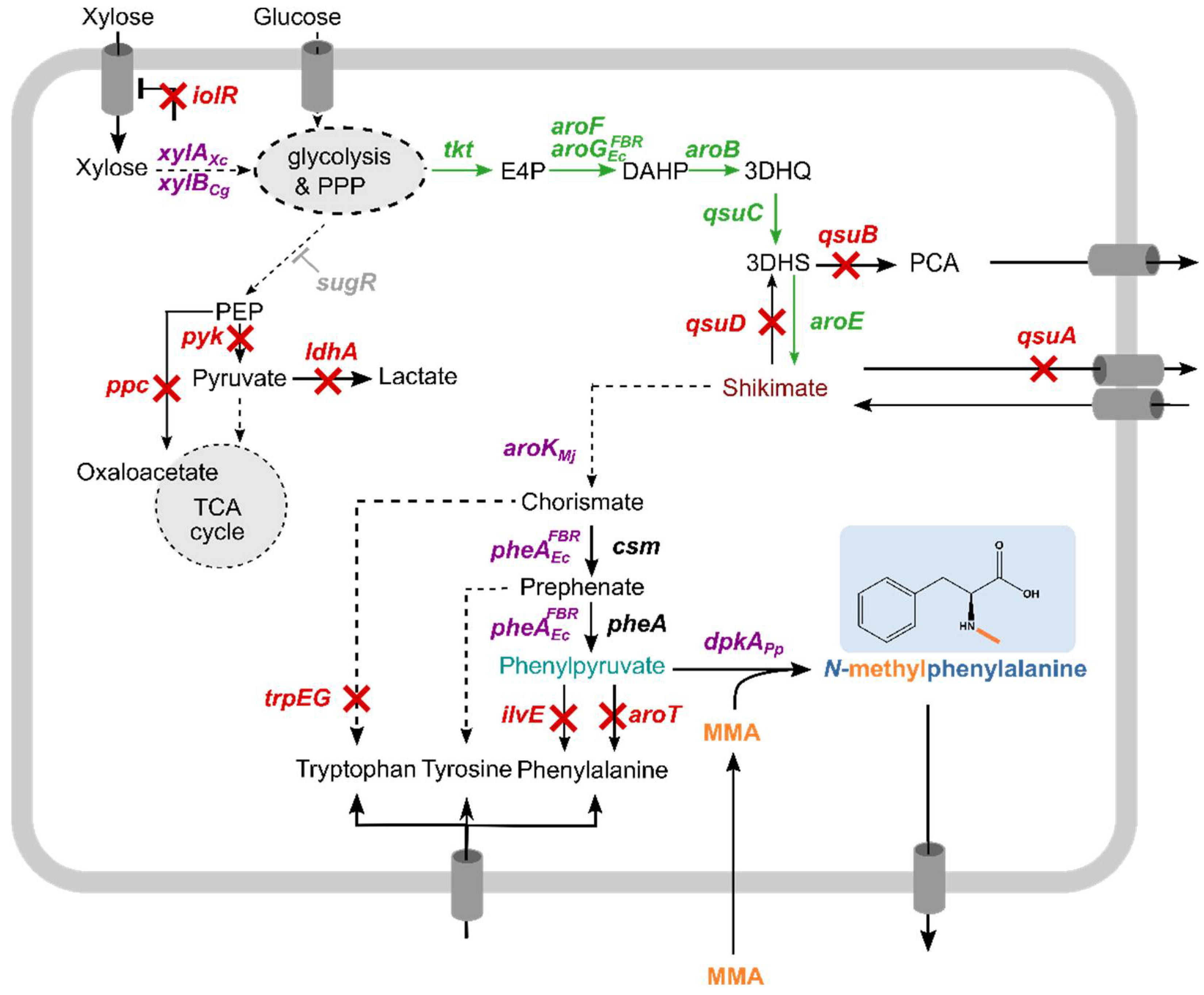
3.1. Assessing the Suitability of C. glutamicum for NMePhe Production
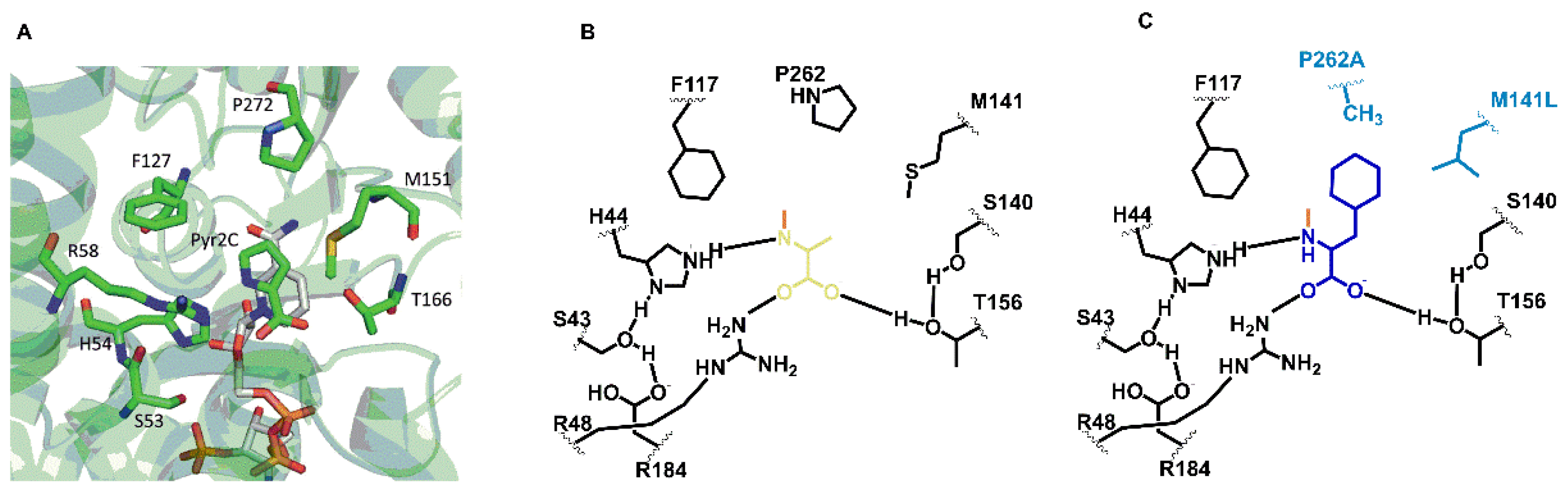
3.2. Exchanging Two Amino Acid Residues of DpkA to Affect Preference of the Substrates Pyruvate and Phenylpyruvate
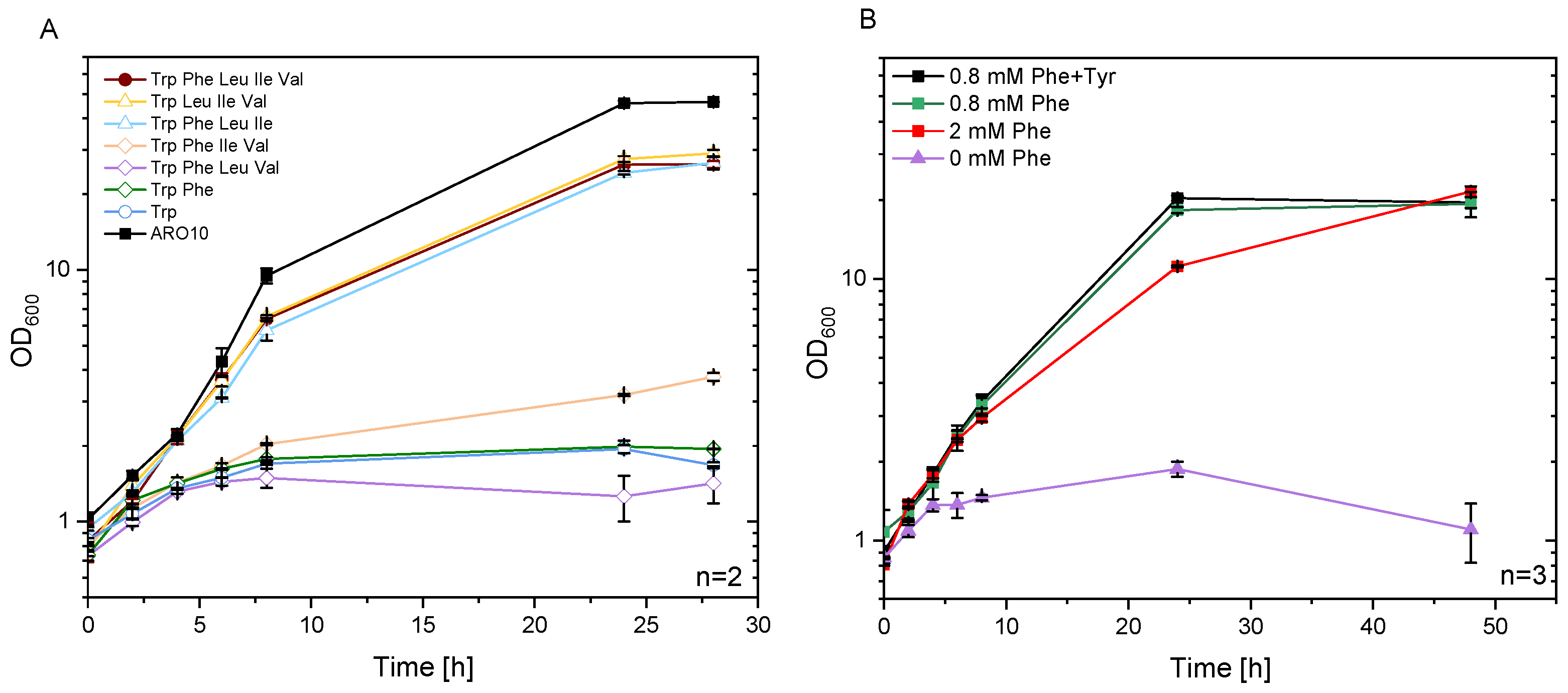
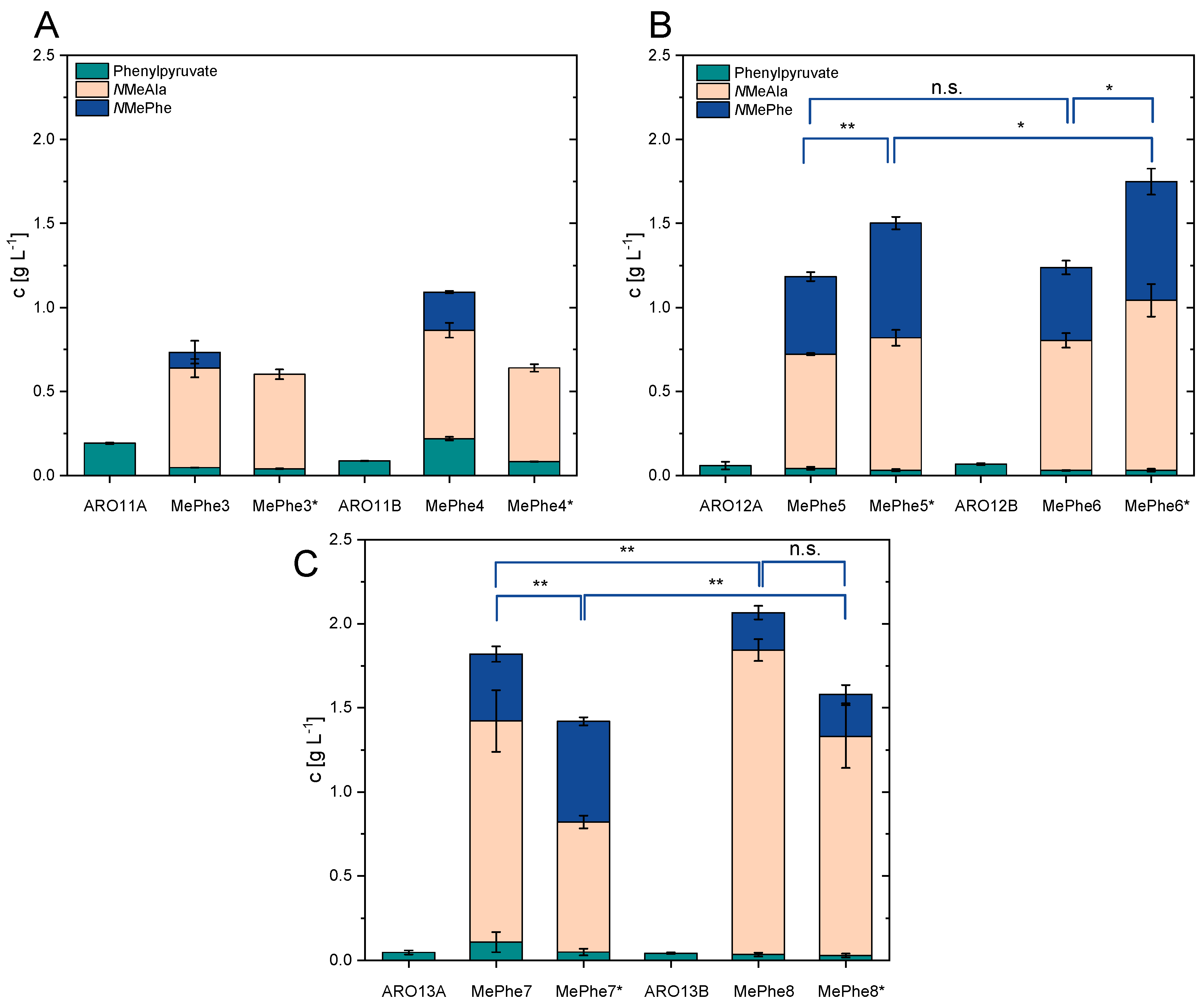
3.3. Metabolic Engineering of C. glutamicum for Efficient Provision of Phenylpyruvate as Precursor
3.4. DpkA Mediated Methylamination of Phenylpyruvate Yielded NMePhe
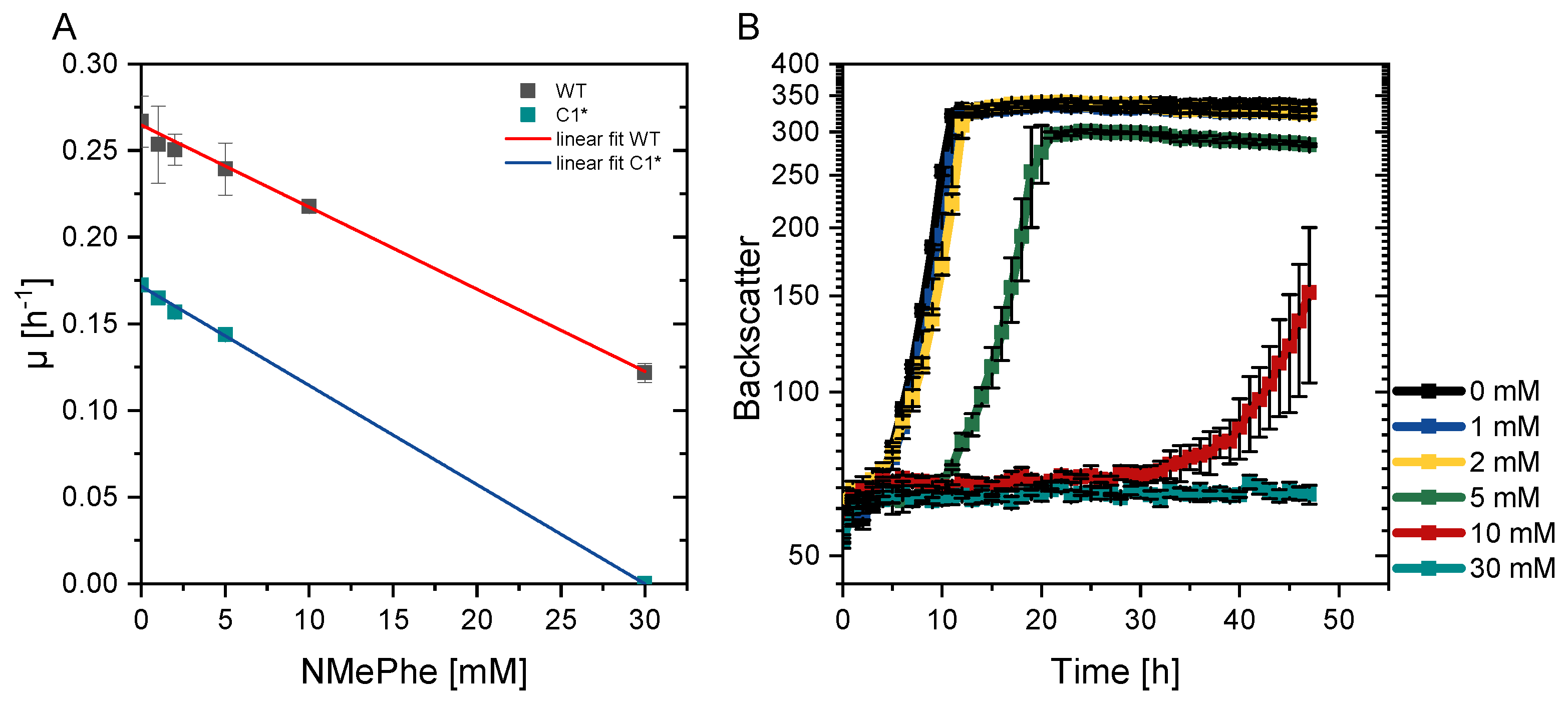
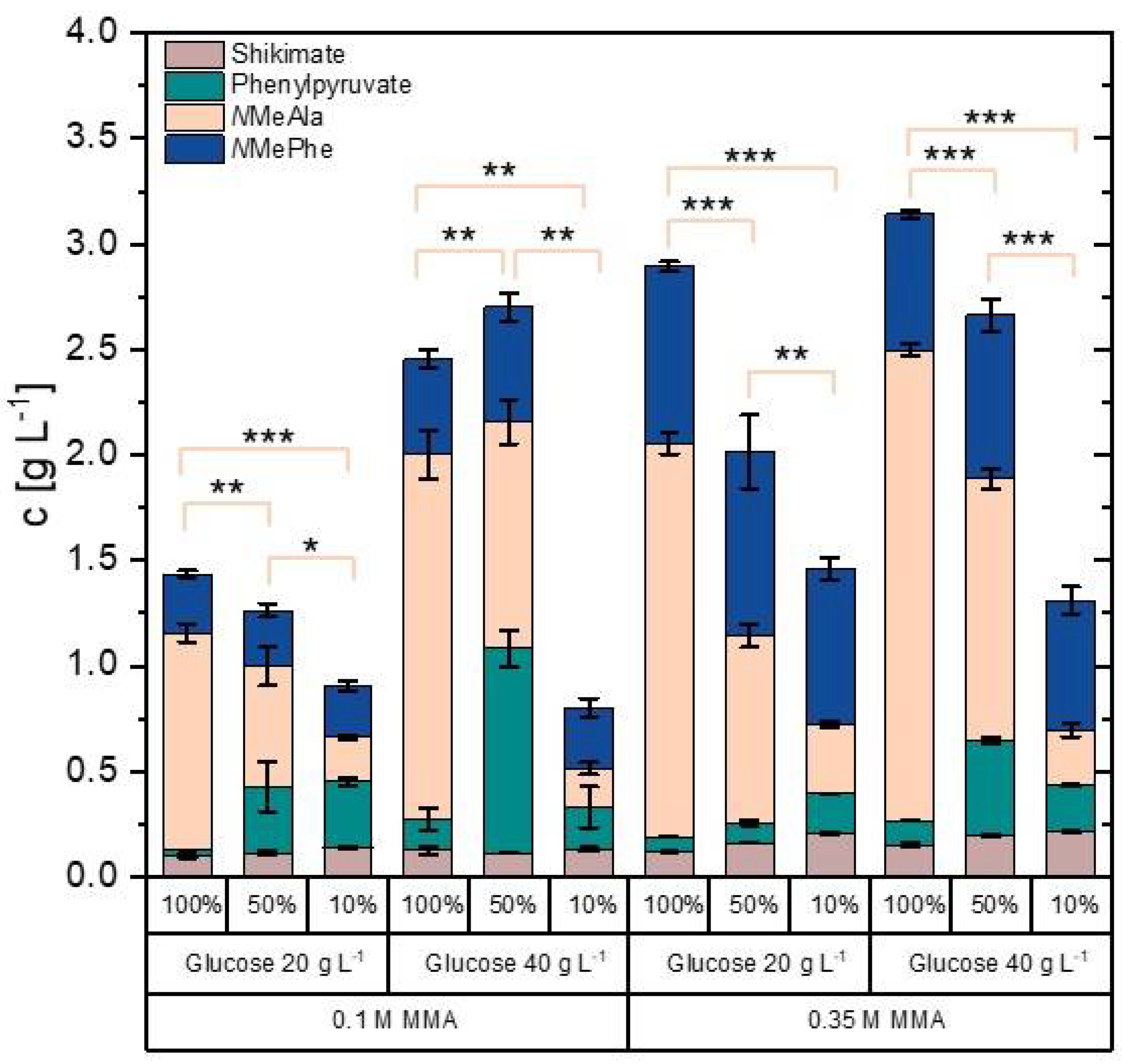
3.5. Improvement of NMePhe Production and Reduction of NMeAla
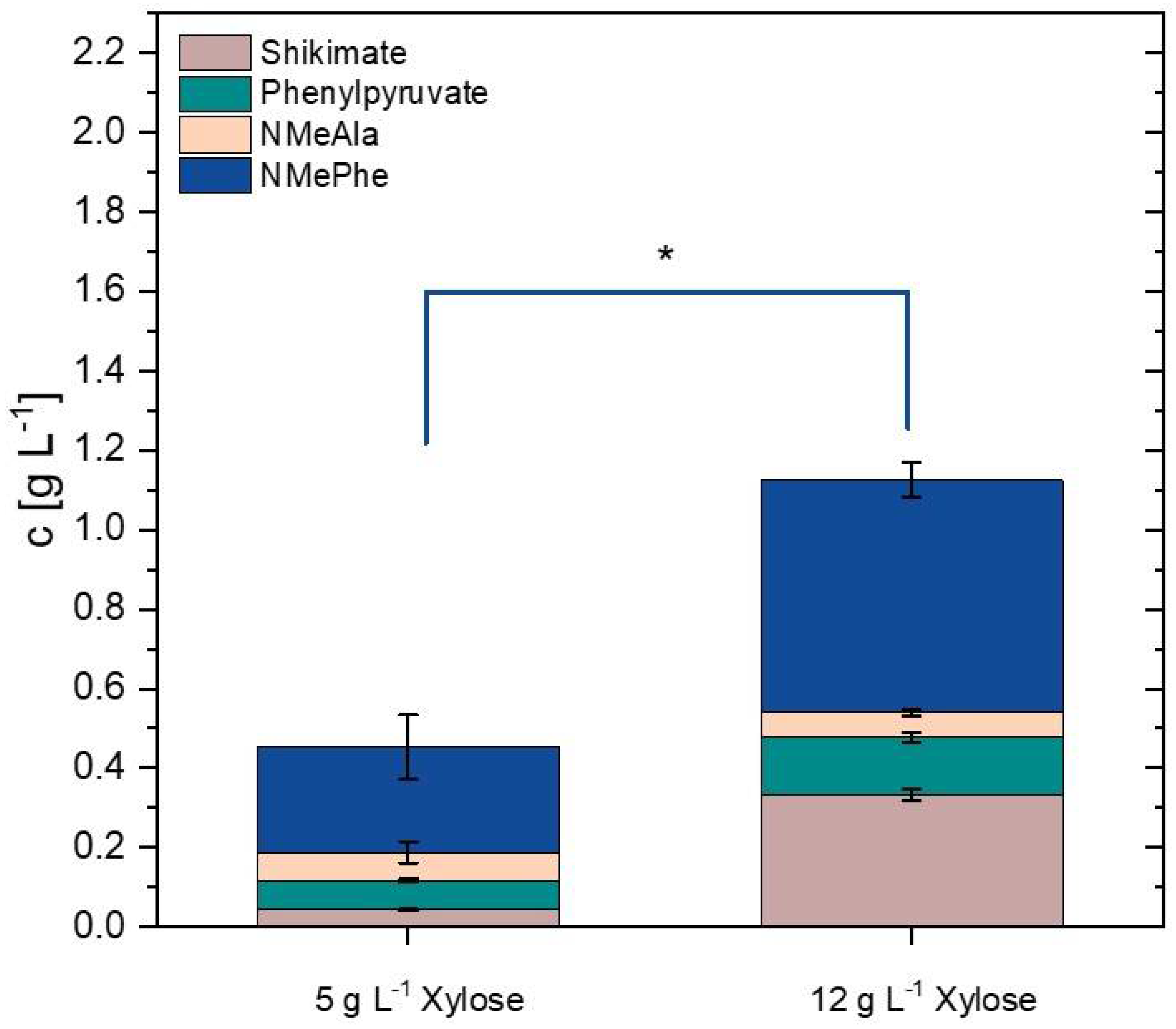
3.6. Establishing NMePhe Production from the Alternative Feedstock Xylose
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cartwright, R.A.; Roberts, E.A.H.; Wood, D.J. Theanine, an amino-acid n-ethyl amide present in tea. J. Sci. Food Agric. 1954, 5, 597–599. [Google Scholar] [CrossRef]
- Sakato, Y. The chemical constituents of tea; III. A new amide theanine. Nippon Nogeikagaku Kaishi 1949, 23, 262–267. [Google Scholar] [CrossRef]
- Luka, Z.; Mudd, S.H.; Wagner, C. Glycine N-Methyltransferase and Regulation of S-Adenosylmethionine Levels. J. Biol. Chem. 2009, 284, 22507–22511. [Google Scholar] [CrossRef]
- Ducker, G.S.; Rabinowitz, J.D. One-Carbon Metabolism in Health and Disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef]
- Wilcken, B. Therapeutic targets in homocystinuria due to cystathionine β-synthase deficiency: New European guidelines. Expert Opin. Orphan Drugs 2017, 5, 1–3. [Google Scholar] [CrossRef][Green Version]
- Latypova, E.; Yang, S.; Wang, Y.-S.; Wang, T.; Chavkin, T.A.; Hackett, M.; Schäfer, H.; Kalyuzhnaya, M.G. Genetics of the glutamate-mediated methylamine utilization pathway in the facultative methylotrophic beta-proteobacterium Methyloversatilis universalis FAM5. Mol. Microbiol. 2010, 75, 426–439. [Google Scholar] [CrossRef]
- Chatterjee, J.; Rechenmacher, F.; Kessler, H. N-Methylation of Peptides and Proteins: An Important Element for Modulating Biological Functions. Angew. Chem. Int. Ed. 2013, 52, 254–269. [Google Scholar] [CrossRef]
- Aurelio, L.; Brownlee, R.T.C.; Hughes, A.B. Synthetic Preparation of N-Methyl-α-amino Acids. Chem. Rev. 2004, 104, 5823–5846. [Google Scholar] [CrossRef] [PubMed]
- Cody, W.L.; He, J.X.; Reily, M.D.; Haleen, S.J.; Walker, D.M.; Reyner, E.L.; Stewart, B.H.; Doherty, A.M. Design of a Potent Combined Pseudopeptide Endothelin-A/Endothelin-B Receptor Antagonist, Ac-DBhg16-Leu-Asp-Ile-[NMe]Ile-Trp21 (PD 156252): Examination of Its Pharmacokinetic and Spectral Properties. J. Med. Chem. 1997, 40, 2228–2240. [Google Scholar] [CrossRef]
- Gordon, D.J.; Tappe, R.; Meredith, S.C. Design and characterization of a membrane permeable N-methyl amino acid-containing peptide that inhibits Aβ1–40 fibrillogenesis. J. Pept. Res. 2002, 60, 37–55. [Google Scholar] [CrossRef]
- Biron, E.; Chatterjee, J.; Ovadia, O.; Langenegger, D.; Brueggen, J.; Hoyer, D.; Schmid, H.A.; Jelinek, R.; Gilon, C.; Hoffman, A.; et al. Improving Oral Bioavailability of Peptides by Multiple N-Methylation: Somatostatin Analogues. Angew. Chem. Int. Ed. 2008, 47, 2595–2599. [Google Scholar] [CrossRef]
- Soto, C.; Kascsak, R.J.; Saborío, G.P.; Aucouturier, P.; Wisniewski, T.; Prelli, F.; Kascsak, R.; Mendez, E.; Harris, D.A.; Ironside, J.; et al. Reversion of prion protein conformational changes by synthetic beta-sheet breaker peptides. Lancet Lond. Engl. 2000, 355, 192–197. [Google Scholar] [CrossRef]
- Ghanta, J.; Shen, C.-L.; Kiessling, L.L.; Murphy, R.M. A Strategy for Designing Inhibitors of β-Amyloid Toxicity. J. Biol. Chem. 1996, 271, 29525–29528. [Google Scholar] [CrossRef]
- Hughes, E.; Burke, R.M.; Doig, A.J. Inhibition of Toxicity in the β-Amyloid Peptide Fragment β-(25–35) Using N-Methylated Derivatives: A General Strategy to Prevent Amyloid Formation. J. Biol. Chem. 2000, 275, 25109–25115. [Google Scholar] [CrossRef]
- Doig, A.J.; Hughes, E.; Burke, R.M.; Su, T.J.; Heenan, R.K.; Lu, J. Inhibition of toxicity and protofibril formation in the amyloid-β peptide β-(25–35) using N-Methylated derivatives. Biochem. Soc. Trans. 2002, 30, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Kapurniotu, A.; Schmauder, A.; Tenidis, K. Structure-based design and study of non-amyloidogenic, double N-methylated IAPP amyloid core sequences as inhibitors of IAPP amyloid formation and cytotoxicity. J. Mol. Biol. 2002, 315, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Frost, L.S.; Carpenter, M.; Paranchych, W. N-methylphenylalanine at the N-terminus of pilin isolated from Pseudomonas aeruginosa K. Nature 1978, 271, 87–89. [Google Scholar] [CrossRef]
- Malakoutikhah, M.; Prades, R.; Teixidó, M.; Giralt, E. N-Methyl Phenylalanine-Rich Peptides as Highly Versatile Blood-Brain Barrier Shuttles. J. Med. Chem. 2010, 53, 2354–2363. [Google Scholar] [CrossRef]
- Freidinger, R.M.; Hinkle, J.S.; Perlow, D.S. Synthesis of 9-fluorenylmethyloxycarbonyl-protected N-alkyl amino acids by reduction of oxazolidinones. J. Org. Chem. 1983, 48, 77–81. [Google Scholar] [CrossRef]
- De Marco, R.; Leggio, A.; Liguori, A.; Marino, T.; Perri, F.; Russo, N. Site-Selective Methylation of Nβ-Nosyl Hydrazides of N-Nosyl Protected α-Amino Acids. J. Org. Chem. 2010, 75, 3381–3386. [Google Scholar] [CrossRef] [PubMed]
- Hyslop, J.F.; Lovelock, S.L.; Watson, A.J.B.; Sutton, P.W.; Roiban, G.-D. N-Alkyl-α-amino acids in Nature and their biocatalytic preparation. J. Biotechnol. 2019, 293, 56–65. [Google Scholar] [CrossRef]
- Mindt, M.; Walter, T.; Risse, J.M.; Wendisch, V.F. Fermentative Production of N-Methylglutamate from Glycerol by Recombinant Pseudomonas putida. Front. Bioeng. Biotechnol. 2018, 6. [Google Scholar] [CrossRef]
- Goto, M.; Muramatsu, H.; Mihara, H.; Kurihara, T.; Esaki, N.; Omi, R.; Miyahara, I.; Hirotsu, K. Crystal Structures of Δ1-Piperideine-2-carboxylate/Δ1-Pyrroline-2-carboxylate Reductase Belonging to a New Family of NAD(P)H-dependent Oxidoreductases: Conformational Change, Substrate Recognition, and Stereochemistry of the Reaction. J. Biol. Chem. 2005, 280, 40875–40884. [Google Scholar] [CrossRef] [PubMed]
- Wieschalka, S.; Blombach, B.; Eikmanns, B.J. Engineering Corynebacterium glutamicum for the production of pyruvate. Appl. Microbiol. Biotechnol. 2012, 94, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Krause, F.S.; Blombach, B.; Eikmanns, B.J. Metabolic Engineering of Corynebacterium glutamicum for 2-Ketoisovalerate Production. Appl. Environ. Microbiol. 2010, 76, 8053–8061. [Google Scholar] [CrossRef] [PubMed]
- Bückle-Vallant, V.; Krause, F.S.; Messerschmidt, S.; Eikmanns, B.J. Metabolic engineering of Corynebacterium glutamicum for 2-ketoisocaproate production. Appl. Microbiol. Biotechnol. 2014, 98, 297–311. [Google Scholar] [CrossRef]
- Mindt, M.; Heuser, M.; Wendisch, V.F. Xylose as preferred substrate for sarcosine production by recombinant Corynebacterium glutamicum. Bioresour. Technol. 2019, 281, 135–142. [Google Scholar] [CrossRef]
- Mindt, M.; Risse, J.M.; Gruß, H.; Sewald, N.; Eikmanns, B.J.; Wendisch, V.F. One-step process for production of N-methylated amino acids from sugars and methylamine using recombinant Corynebacterium glutamicum as biocatalyst. Sci. Rep. 2018, 8, 12895. [Google Scholar] [CrossRef]
- Mindt, M.; Hannibal, S.; Heuser, M.; Risse, J.M.; Sasikumar, K.; Nampoothiri, K.M.; Wendisch, V.F. Fermentative Production of N-Alkylated Glycine Derivatives by Recombinant Corynebacterium glutamicum Using a Mutant of Imine Reductase DpkA from Pseudomonas putida. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef]
- Pérez-García, F.; Risse, J.M.; Friehs, K.; Wendisch, V.F. Fermentative production of l-pipecolic acid from glucose and alternative carbon sources. Biotechnol. J. 2017, 12, 1600646. [Google Scholar] [CrossRef]
- Pérez-García, F.; Peters-Wendisch, P.; Wendisch, V.F. Engineering Corynebacterium glutamicum for fast production of l-lysine and l-pipecolic acid. Appl. Microbiol. Biotechnol. 2016, 100, 8075–8090. [Google Scholar] [CrossRef]
- Yi, Y.; Sheng, H.; Li, Z.; Ye, Q. Biosynthesis of trans-4-hydroxyproline by recombinant strains of Corynebacterium glutamicum and Escherichia coli. BMC Biotechnol. 2014, 14, 44. [Google Scholar] [CrossRef]
- Falcioni, F.; Bühler, B.; Schmid, A. Efficient hydroxyproline production from glucose in minimal media by Corynebacterium glutamicum. Biotechnol. Bioeng. 2015, 112, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Veldmann, K.H.; Dachwitz, S.; Risse, J.M.; Lee, J.-H.; Sewald, N.; Wendisch, V.F. Bromination of L-tryptophan in a Fermentative Process with Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2019, 7, 219. [Google Scholar] [CrossRef]
- Veldmann, K.H.; Minges, H.; Sewald, N.; Lee, J.-H.; Wendisch, V.F. Metabolic engineering of Corynebacterium glutamicum for the fermentative production of halogenated tryptophan. J. Biotechnol. 2019, 291, 7–16. [Google Scholar] [CrossRef]
- Walter, T.; Al Medani, N.; Burgardt, A.; Cankar, K.; Ferrer, L.; Kerbs, A.; Lee, J.-H.; Mindt, M.; Risse, J.M.; Wendisch, V.F. Fermentative N-Methylanthranilate Production by Engineered Corynebacterium glutamicum. Microorganisms 2020, 8, 866. [Google Scholar] [CrossRef]
- Shimizu, H.; Hirasawa, T. Production of Glutamate and Glutamate-Related Amino Acids: Molecular Mechanism Analysis and Metabolic Engineering. In Amino Acid Biosynthesis—Pathways, Regulation and Metabolic Engineering; Microbiology Monographs; Wendisch, V.F., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–38. ISBN 978-3-540-48596-4. [Google Scholar] [CrossRef]
- Wendisch, V.F. L-Lysine. Industrial Biotechnology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 361–390. ISBN 978-3-527-80783-3. [Google Scholar] [CrossRef]
- Wendisch, V.F. Metabolic engineering advances and prospects for amino acid production. Metab. Eng. 2020, 58, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Priefer, U.; Pühler, A. A Broad Host Range Mobilization System for in vivo Genetic Engineering: Transposon Mutagenesis in Gram Negative Bacteria. Bio/Technology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Baumgart, M.; Unthan, S.; Kloß, R.; Radek, A.; Polen, T.; Tenhaef, N.; Müller, M.F.; Küberl, A.; Siebert, D.; Brühl, N.; et al. Corynebacterium glutamicum Chassis C1*: Building and Testing a Novel Platform Host for Synthetic Biology and Industrial Biotechnology. ACS Synth. Biol. 2018, 7, 132–144. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012; ISBN 978-1-936113-41-5. [Google Scholar]
- Eggeling, L.; Bott, M.; Bott, M. Handbook of Corynebacterium glutamicum; CRC Press: Boca Raton, FL, USA, 2005; ISBN 978-1-4200-3969-6. [Google Scholar] [CrossRef]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Heider, S.A.E.; Peters-Wendisch, P.; Netzer, R.; Stafnes, M.; Brautaset, T.; Wendisch, V.F. Production and glucosylation of C50 and C40 carotenoids by metabolically engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2014, 98, 1223–1235. [Google Scholar] [CrossRef]
- Johnson, K.A.; Goody, R.S. The Original Michaelis Constant: Translation of the 1913 Michaelis-Menten Paper. Biochemistry 2011, 50, 8264–8269. [Google Scholar] [CrossRef]
- Schneider, J.; Eberhardt, D.; Wendisch, V.F. Improving putrescine production by Corynebacterium glutamicum by fine-tuning ornithine transcarbamoylase activity using a plasmid addiction system. Appl. Microbiol. Biotechnol. 2012, 95, 169–178. [Google Scholar] [CrossRef]
- Mihara, H.; Muramatsu, H.; Kakutani, R.; Yasuda, M.; Ueda, M.; Kurihara, T.; Esaki, N. N-Methyl-l-amino acid dehydrogenase from Pseudomonas putida. FEBS J. 2005, 272, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Meiswinkel, T.M.; Gopinath, V.; Lindner, S.N.; Nampoothiri, K.M.; Wendisch, V.F. Accelerated pentose utilization by Corynebacterium glutamicum for accelerated production of lysine, glutamate, ornithine and putrescine. Microb. Biotechnol. 2013, 6, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, V.; Meiswinkel, T.M.; Wendisch, V.F.; Nampoothiri, K.M. Amino acid production from rice straw and wheat bran hydrolysates by recombinant pentose-utilizing Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2011, 92, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, H.; Mihara, H.; Kakutani, R.; Yasuda, M.; Ueda, M.; Kurihara, T.; Esaki, N. Enzymatic synthesis of N-methyl-l-phenylalanine by a novel enzyme, N-methyl-l-amino acid dehydrogenase, from Pseudomonas putida. Tetrahedron Asymmetry 2004, 15, 2841–2843. [Google Scholar] [CrossRef]
- Ikeda, M. Lysine Fermentation: History and Genome Breeding. In Amino Acid Fermentation; Advances in Biochemical Engineering/Biotechnology; Yokota, A., Ikeda, M., Eds.; Springer: Tokyo, Japan, 2017; pp. 73–102. ISBN 978-4-431-56520-8. [Google Scholar] [CrossRef]
- Hermann, T. Industrial production of amino acids by coryneform bacteria. J. Biotechnol. 2003, 104, 155–172. [Google Scholar] [CrossRef]
- Farinas, E.T.; Schwaneberg, U.; Glieder, A.; Arnold, F.H. Directed Evolution of a Cytochrome P450 Monooxygenase for Alkane Oxidation. Adv. Synth. Catal. 2001, 343, 601–606. [Google Scholar] [CrossRef]
- Reetz, M.T.; Carballeira, J.D. Iterative saturation mutagenesis (ISM) for rapid directed evolution of functional enzymes. Nat. Protoc. 2007, 2, 891–903. [Google Scholar] [CrossRef]
- Gupta, R.D.; Tawfik, D.S. Directed enzyme evolution via small and effective neutral drift libraries. Nat. Methods 2008, 5, 939–942. [Google Scholar] [CrossRef]
- Borlinghaus, N.; Nestl, B.M. Switching the Cofactor Specificity of an Imine Reductase. ChemCatChem 2018, 10, 183–187. [Google Scholar] [CrossRef]
- Muramatsu, H.; Mihara, H.; Goto, M.; Miyahara, I.; Hirotsu, K.; Kurihara, T.; Esaki, N. A new family of NAD(P)H-dependent oxidoreductases distinct from conventional Rossmann-fold proteins. J. Biosci. Bioeng. 2005, 99, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Katsumata, R. Hyperproduction of Tryptophan by Corynebacterium glutamicum with the Modified Pentose Phosphate Pathway. Appl. Environ. Microbiol. 1999, 65, 2497–2502. [Google Scholar] [CrossRef] [PubMed]
- Kitade, Y.; Hashimoto, R.; Suda, M.; Hiraga, K.; Inui, M. Production of 4-Hydroxybenzoic Acid by an Aerobic Growth-Arrested Bioprocess Using Metabolically Engineered Corynebacterium glutamicum. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Syukur Purwanto, H.; Kang, M.-S.; Ferrer, L.; Han, S.-S.; Lee, J.-Y.; Kim, H.-S.; Lee, J.-H. Rational engineering of the shikimate and related pathways in Corynebacterium glutamicum for 4-hydroxybenzoate production. J. Biotechnol. 2018, 282, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.-S.; Lee, D.; Lee, S.J.; Chun, G.-T.; Choi, S.-S.; Kim, E.-S.; Kim, S. Characterization of a non-phosphotransferase system for cis,cis-muconic acid production in Corynebacterium glutamicum. Biochem. Biophys. Res. Commun. 2018, 499, 279–284. [Google Scholar] [CrossRef]
- Kallscheuer, N.; Marienhagen, J. Corynebacterium glutamicum as platform for the production of hydroxybenzoic acids. Microb. Cell Factories 2018, 17, 70. [Google Scholar] [CrossRef]
- Milke, L.; Mutz, M.; Marienhagen, J. Synthesis of the character impact compound raspberry ketone and additional flavoring phenylbutanoids of biotechnological interest with Corynebacterium glutamicum. Microb. Cell Factories 2020, 19, 92. [Google Scholar] [CrossRef]
- Kogure, T.; Suda, M.; Hiraga, K.; Inui, M. Protocatechuate overproduction by Corynebacterium glutamicum via simultaneous engineering of native and heterologous biosynthetic pathways. Metab. Eng. 2020. [Google Scholar] [CrossRef]
- Toivari, M.H.; Salusjärvi, L.; Ruohonen, L.; Penttilä, M. Endogenous Xylose Pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2004, 70, 3681–3686. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Rao, C.V. Microbial conversion of xylose into useful bioproducts. Appl. Microbiol. Biotechnol. 2018, 102, 9015–9036. [Google Scholar] [CrossRef]
- Jeffries, T.W. Engineering yeasts for xylose metabolism. Curr. Opin. Biotechnol. 2006, 17, 320–326. [Google Scholar] [CrossRef]
- Zhang, M.; Eddy, C.; Deanda, K.; Finkelstein, M.; Picataggio, S. Metabolic Engineering of a Pentose Metabolism Pathway in Ethanologenic Zymomonas mobilis. Science 1995, 267, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Weimberg, R. Pentose Oxidation by Pseudomonas fragi. J. Biol. Chem. 1961, 236, 629–635. [Google Scholar] [CrossRef]
- Radek, A.; Krumbach, K.; Gätgens, J.; Wendisch, V.F.; Wiechert, W.; Bott, M.; Noack, S.; Marienhagen, J. Engineering of Corynebacterium glutamicum for minimized carbon loss during utilization of d-xylose containing substrates. J. Biotechnol. 2014, 192, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, H.; Vertès, A.A.; Okino, S.; Inui, M.; Yukawa, H. Engineering of a Xylose Metabolic Pathway in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2006, 72, 3418–3428. [Google Scholar] [CrossRef]
- Pérez-García, F.; Wendisch, V.F. Transport and metabolic engineering of the cell factory Corynebacterium glutamicum. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Brüsseler, C.; Radek, A.; Tenhaef, N.; Krumbach, K.; Noack, S.; Marienhagen, J. The myo-inositol/proton symporter IolT1 contributes to d-xylose uptake in Corynebacterium glutamicum. Bioresour. Technol. 2018, 249, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Jojima, T.; Kawaguchi, H.; Inui, M.; Yukawa, H. Engineering of pentose transport in Corynebacterium glutamicum to improve simultaneous utilization of mixed sugars. Appl. Microbiol. Biotechnol. 2009, 85, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Niermann, K.; Wendisch, V.F. Production of the amino acids l-glutamate, l-lysine, l-ornithine and l-arginine from arabinose by recombinant Corynebacterium glutamicum. J. Biotechnol. 2011, 154, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, D.; Lindner, S.N.; Wendisch, V.F. Engineering of a glycerol utilization pathway for amino acid production by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2008, 74, 6216–6222. [Google Scholar] [CrossRef] [PubMed]
- Meiswinkel, T.M.; Rittmann, D.; Lindner, S.N.; Wendisch, V.F. Crude glycerol-based production of amino acids and putrescine by Corynebacterium glutamicum. Bioresour. Technol. 2013, 145, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Uhde, A.; Youn, J.-W.; Maeda, T.; Clermont, L.; Matano, C.; Krämer, R.; Wendisch, V.F.; Seibold, G.M.; Marin, K. Glucosamine as carbon source for amino acid-producing Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2013, 97, 1679–1687. [Google Scholar] [CrossRef]
- Matano, C.; Uhde, A.; Youn, J.-W.; Maeda, T.; Clermont, L.; Marin, K.; Krämer, R.; Wendisch, V.F.; Seibold, G.M. Engineering of Corynebacterium glutamicum for growth and L-lysine and lycopene production from N-acetyl-glucosamine. Appl. Microbiol. Biotechnol. 2014, 98, 5633–5643. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Source |
|---|---|---|
| Escherichia coli | ||
| S17-1 | recA pro hsdR RP4-2-Tc::Mu-Km::Tn7 | [40] |
| DH5α | F-thi-1 endA1 hsdr17(r-, m-) supE44 1lacU169 (Φ80lacZ1M15) recA1 gyrA96 | [41] |
| BL21(DE3) | fhuA2 [lon] ompT gal (λDE3) [dcm] ΔhsdS λ DE3 = λ sBamHIo ΔEcoRI-B int::(lacI::PlacUV5::T7°gene1) i21 Δnin5 | Novagen |
| Corynebacterium glutamicum | ||
| WT | C. glutamicum wild type, ATCC 13032 | ATCC |
| C1* | Genome-reduced strain derived from C. glutamicum WT | [42] |
| ARO9 | Δvdh::PilvC-aroGD146N ΔldhA ΔaroR::PilvC-aroF ΔqsuBCD::Ptuf-qsuC Δppc::Psod-aroB ΔPtkt::Ptuf-tkt ΔiolR::Ptuf-aroE | [36] |
| ARO10 | ΔtrpEG mutant of ARO9 | This work |
| ARO10A | ARO10 carrying pEKEx3-pheAFBR and pVWEx1 | This work |
| ARO10B | ARO10 carrying pEKEx3-pheAFBR-aroK and pVWEx1 | This work |
| ARO11 | ΔilvE mutant of ARO10 | This work |
| ARO11A | ARO11 carrying pEKEx3-pheAFBR and pVWEx1 | This work |
| ARO11B | ARO11 carrying pEKEx3-pheAFBR-aroK and pVWEx1 | This work |
| ARO12 | ΔaroT mutant of ARO11 | This work |
| ARO12A | ARO12 carrying pEKEx3-pheAFBR and pVWEx1 | This work |
| ARO12B | ARO12 carrying pEKEx3-pheAFBR-aroK and pVWEx1 | This work |
| ARO13 | Δpyk mutant of ARO12 | This work |
| ARO13A | ARO13 carrying pEKEx3-pheAFBR and pVWEx1 | This work |
| ARO13B | ARO13 carrying pEKEx3-pheAFBR-aroK and pVWEx1 | This work |
| MePhe1 | ARO10 carrying pEKEx3-pheAFBR and pVWEx1-dpkA-RBSopt | This work |
| MePhe2 | ARO10 carrying pEKEx3-pheAFBR-aroK and pVWEx1-dpkA-RBSopt | This work |
| MePhe3 | ARO11 carrying pEKEx3-pheAFBR and pVWEx1-dpkA-RBSopt | This work |
| MePhe3* | ARO11 carrying pEKEx3-pheAFBRand pVWEx1- dpkAP262M141L | This work |
| MePhe4 | ARO11 carrying pEKEx3-pheAFBR-aroK and pVWEx1-dpkA-RBSopt | This work |
| MePhe4* | ARO11 carrying pEKEx3-pheAFBR-aroK and pVWEx1-dpkAP262M141L | This work |
| MePhe5 | ARO12 carrying pEKEx3-pheAFBR and pVWEx1-dpkA-RBSopt | This work |
| MePhe5* | ARO12 carrying pEKEx3-pheAFBRand pVWEx1-dpkAP262M141L | This work |
| MePhe6 | ARO12 carrying pEKEx3-pheAFBR-aroK and pVWEx1-dpkA-RBSopt | This work |
| MePhe6* | ARO12 carrying pEKEx3-pheAFBR-aroK and pVWEx1-dpkAP262M141L | This work |
| MePhe7 | ARO13 carrying pEKEx3-pheAFBR and pVWEx1-dpkA-RBSopt | This work |
| MePhe7* | ARO13 carrying pEKEx3-pheAFBRand pVWEx1-dpkAP262M141L | This work |
| MePhe8 | ARO13 carrying pEKEx3-pheAFBR-aroK and pVWEx1-DpkA-RBSopt | This work |
| MePhe8* | ARO13 carrying pEKEx3-pheAFBR-aroK and pVWEx1-dpkAP262M141L | This work |
| MePhe9* | MePhe5* carrying pEC-XT99Aprytt-xylAXc-XylBCg | This work |
| Enzyme | 2-Oxoacid Substrate | Km (mM) | Specific Activity (U mg−1) | kcat (s−1) | CatalyticEfficiency (s−1 mM−1) |
|---|---|---|---|---|---|
| DpkA | Pyruvate | 5 ± 1 | 32.7 ± 5 | 19.2 | 3.8 |
| DpkA | Phenylpyruvate | 1 ± 0.3 | 2.5 ± 0.2 | 1.5 | 1.5 |
| DpkAP262A,M141L | Pyruvate | 11.4 ± 1 | 9.5 ± 0.5 | 5.6 | 0.5 |
| DpkAP262A,M141L | Phenylpyruvate | 2 ± 0.3 | 2 ± 0.2 | 1.2 | 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerbs, A.; Mindt, M.; Schwardmann, L.; Wendisch, V.F. Sustainable Production of N-methylphenylalanine by Reductive Methylamination of Phenylpyruvate Using Engineered Corynebacterium glutamicum. Microorganisms 2021, 9, 824. https://doi.org/10.3390/microorganisms9040824
Kerbs A, Mindt M, Schwardmann L, Wendisch VF. Sustainable Production of N-methylphenylalanine by Reductive Methylamination of Phenylpyruvate Using Engineered Corynebacterium glutamicum. Microorganisms. 2021; 9(4):824. https://doi.org/10.3390/microorganisms9040824
Chicago/Turabian StyleKerbs, Anastasia, Melanie Mindt, Lynn Schwardmann, and Volker F. Wendisch. 2021. "Sustainable Production of N-methylphenylalanine by Reductive Methylamination of Phenylpyruvate Using Engineered Corynebacterium glutamicum" Microorganisms 9, no. 4: 824. https://doi.org/10.3390/microorganisms9040824
APA StyleKerbs, A., Mindt, M., Schwardmann, L., & Wendisch, V. F. (2021). Sustainable Production of N-methylphenylalanine by Reductive Methylamination of Phenylpyruvate Using Engineered Corynebacterium glutamicum. Microorganisms, 9(4), 824. https://doi.org/10.3390/microorganisms9040824