
Bacterial Communities Associated with Poa annua Roots in Central European (Poland) and Antarctic Settings (King George Island)

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sites and Sampling
2.2. Bacterial Extraction
2.3. DNA Extraction and Targeted 16S rRNA Gene Amplicon Sequencing
2.4. Phenotype Fingerprinting with Biolog EcoPlate™
2.5. Measurement of Soil Components
2.6. Data Analysis
3. Results
3.1. Soil Components
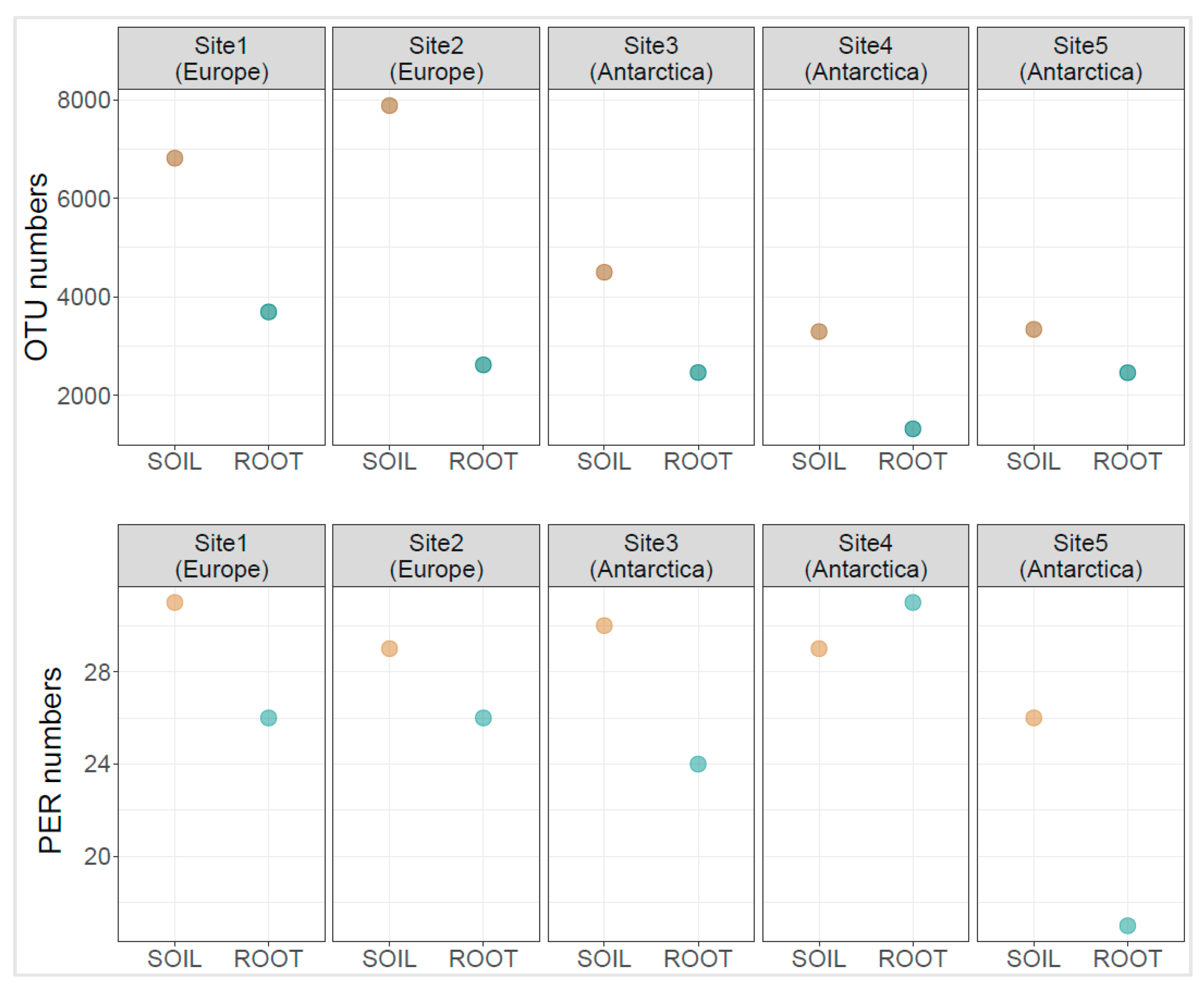
3.2. Diversity Indices
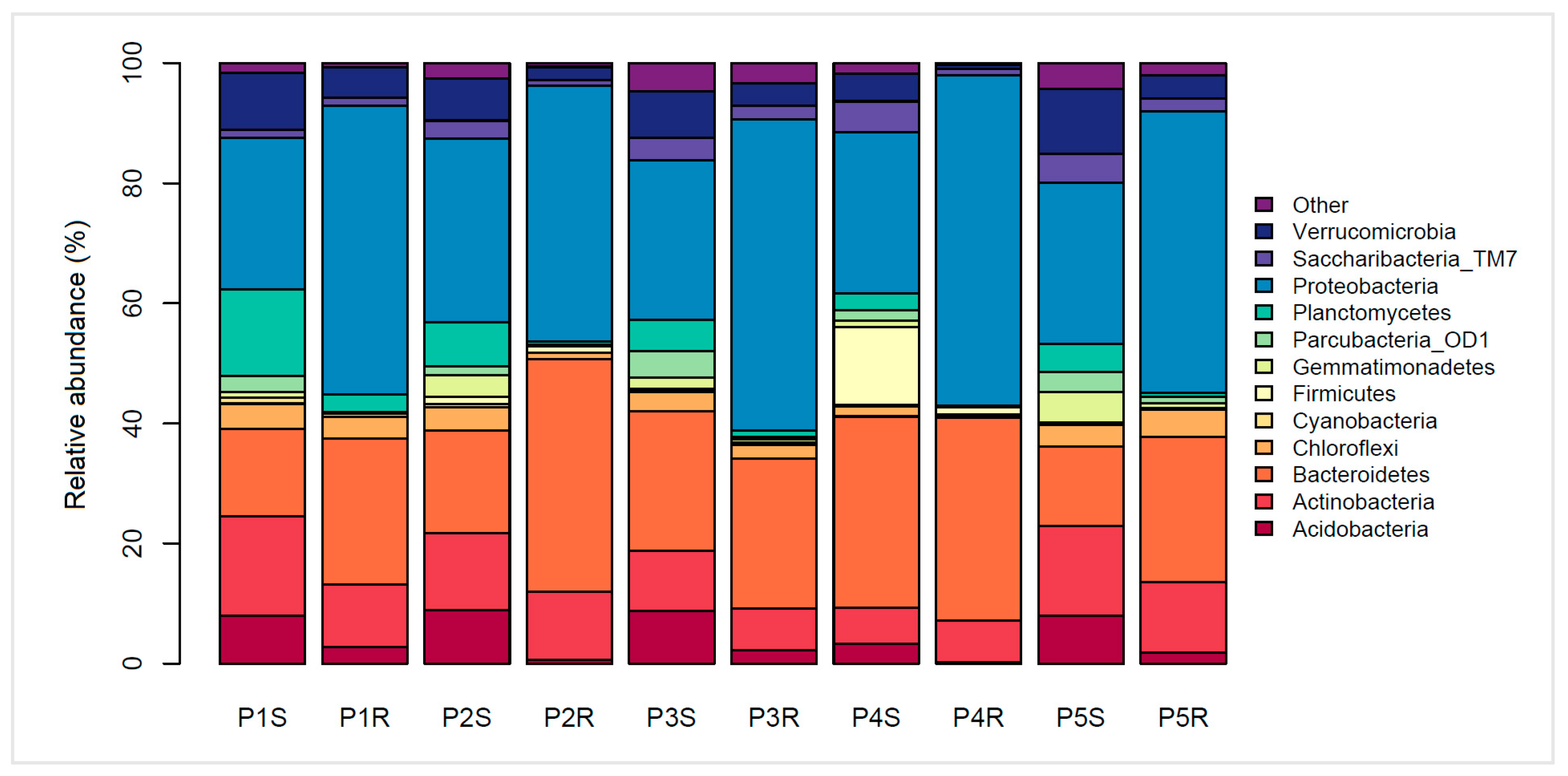
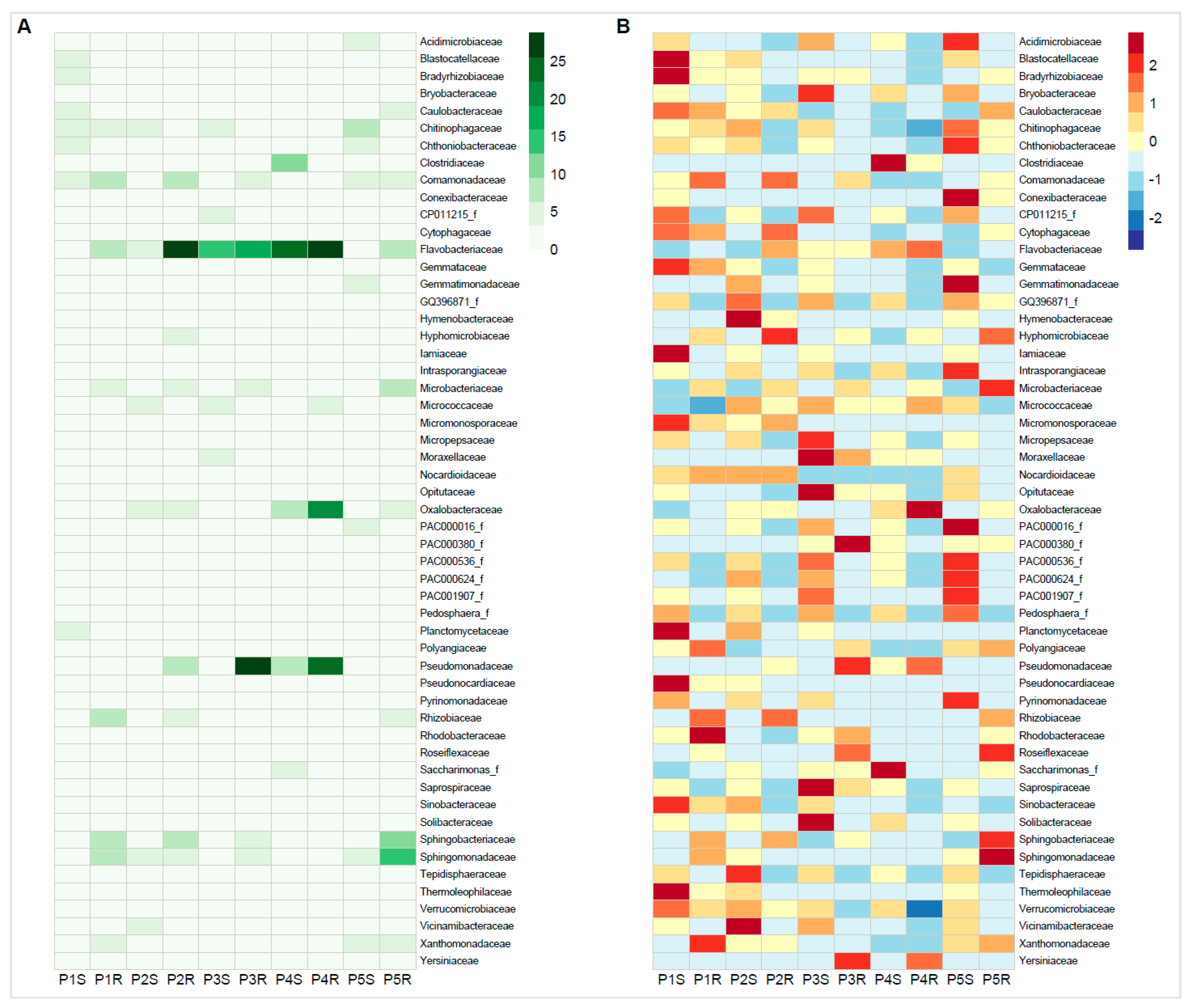
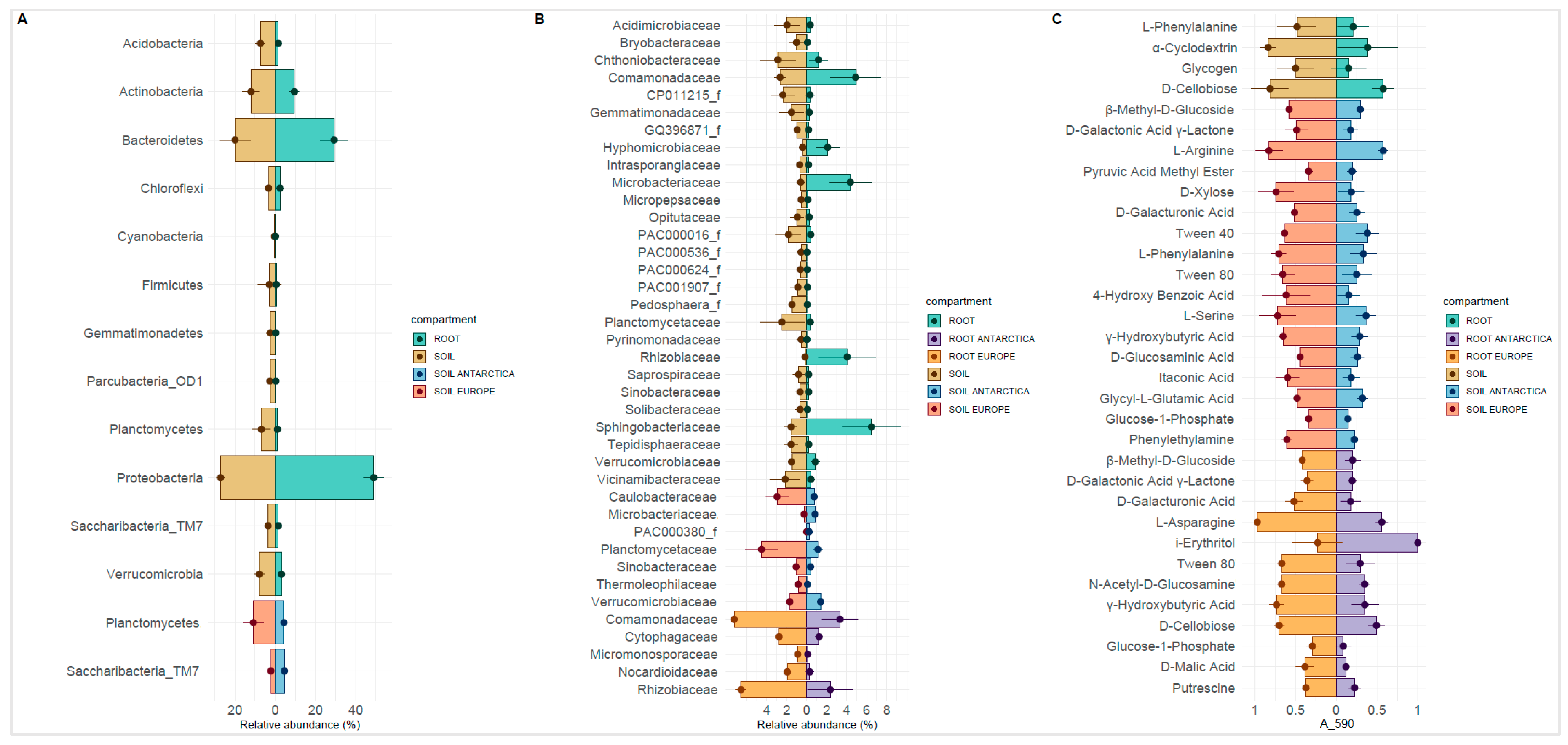
3.3. Bacterial Phylogenetic Diversity
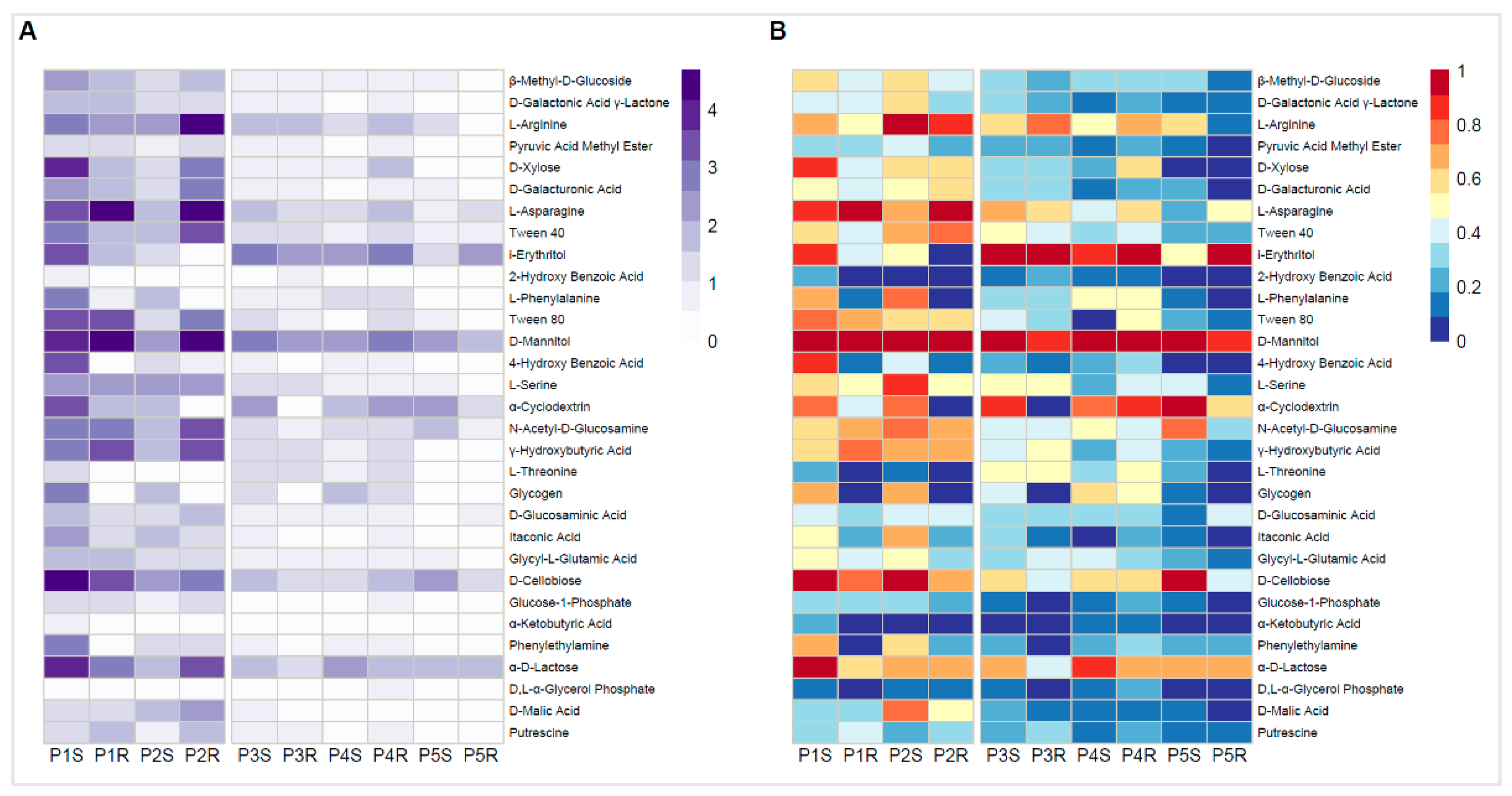
3.4. Community-Level Physiological Profiling by Biolog EcoPlates
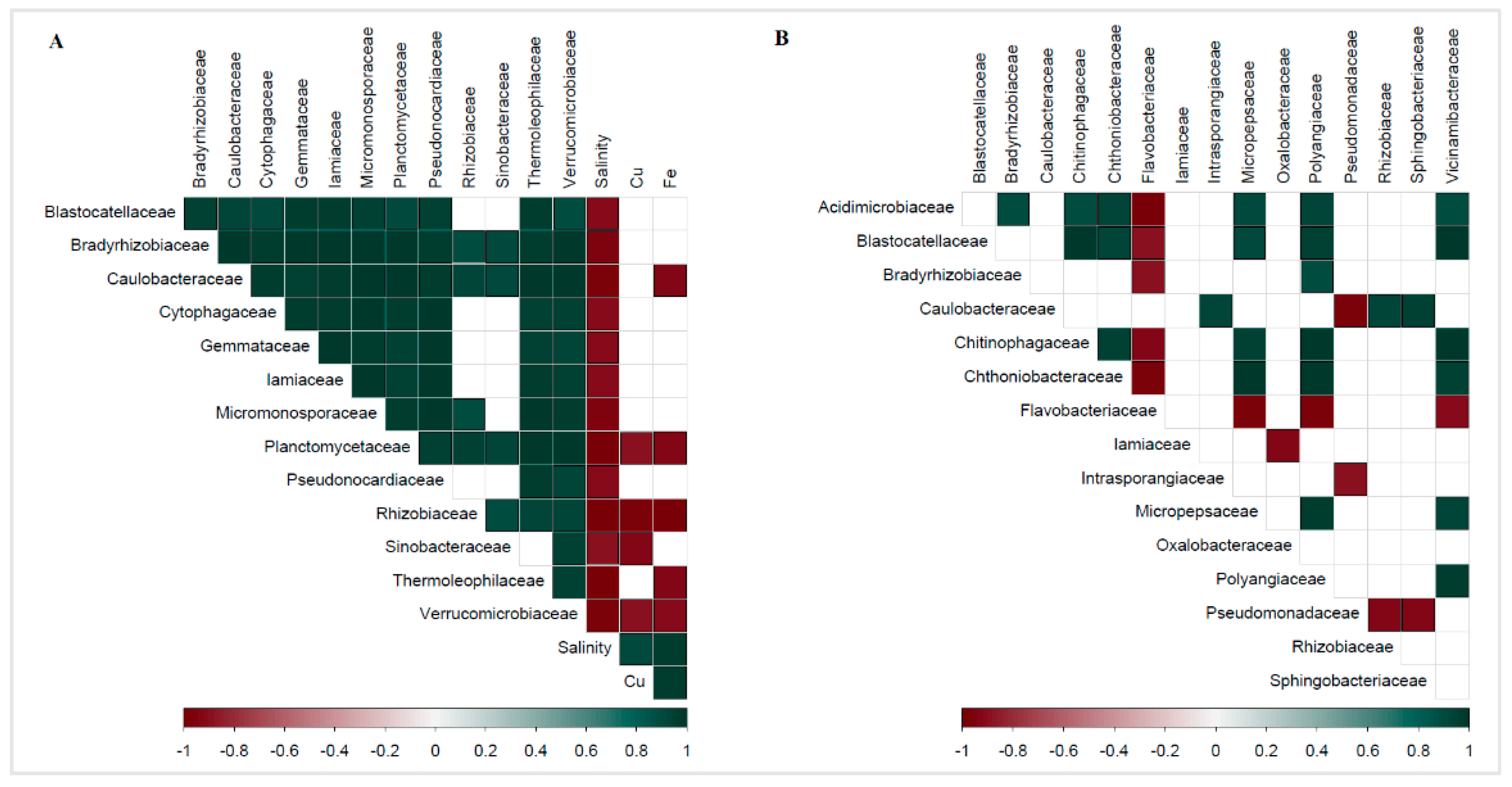
3.5. Correlations between Biological and Geochemical Data
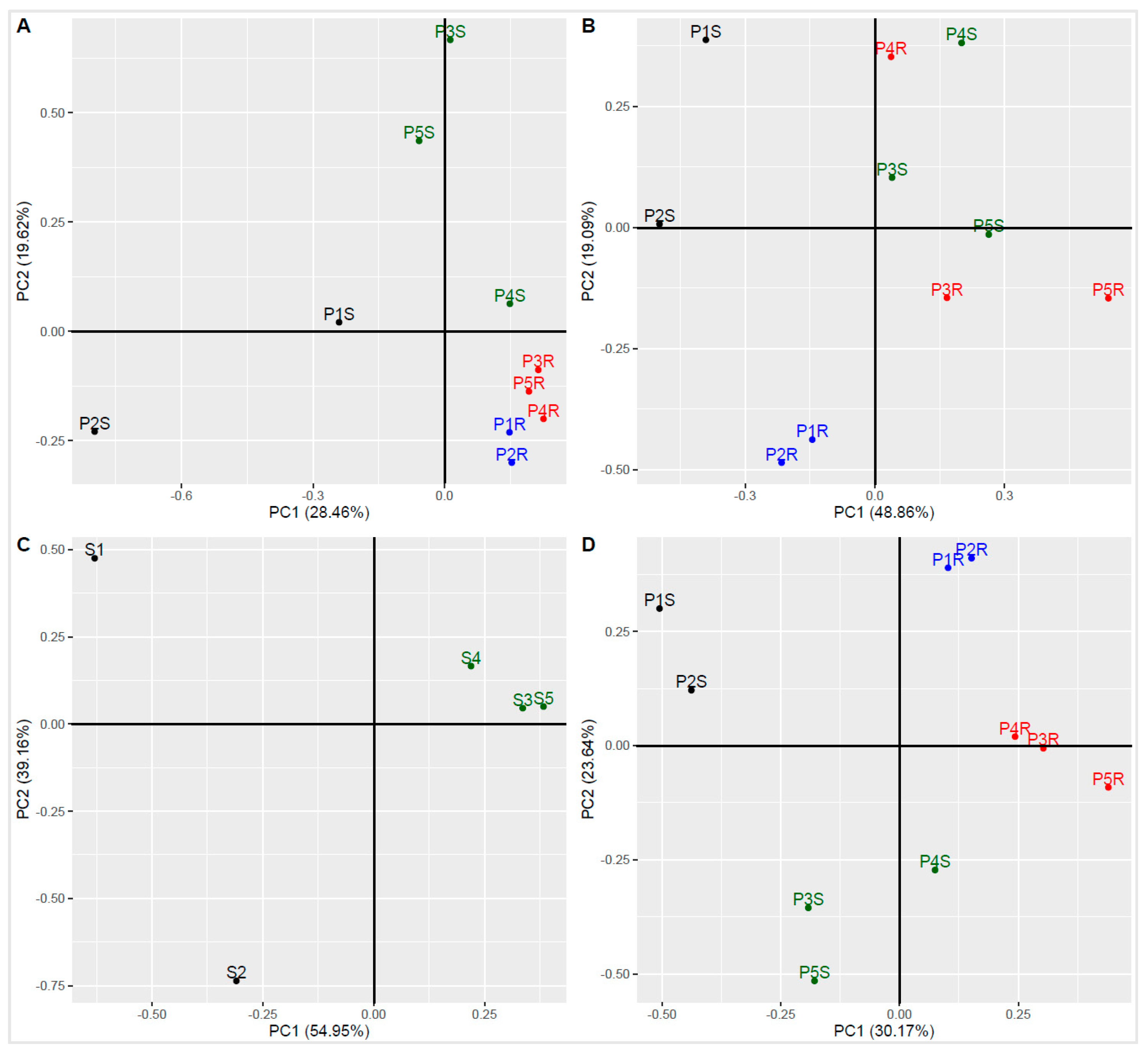
3.6. Principal Component Analysis
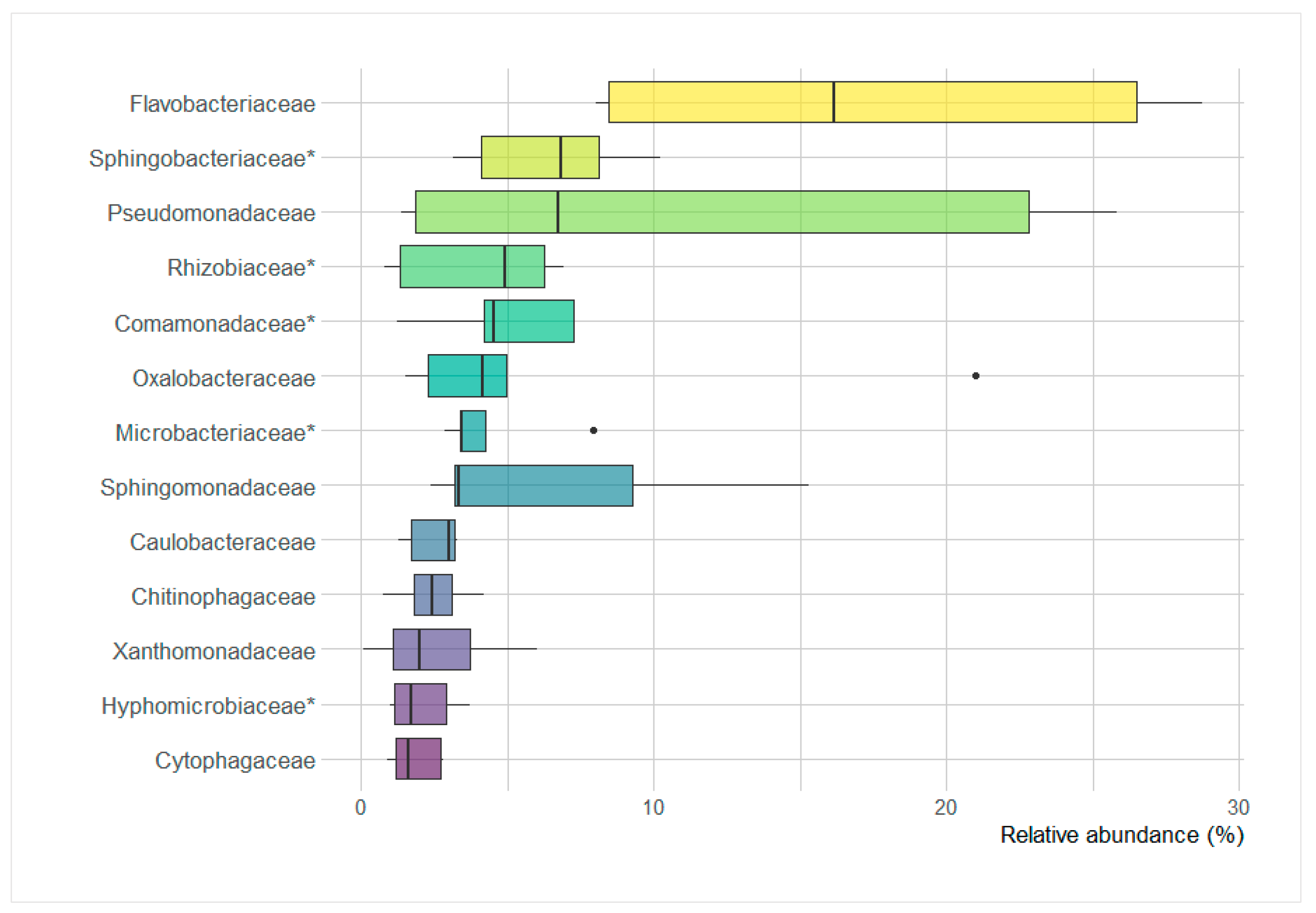
3.7. Significant Differences between Microbial Parameters
3.8. Poa annua Core Microbiome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mao, Q.; Huff, D.R. The evolutionary origin of Poa annua L. Crop. Sci. 2012, 52, 1910–1922. [Google Scholar] [CrossRef]
- Chwedorzewska, K.J. Poa annua L. in Antarctic: Searching for the source of introduction. Polar Biol. 2008, 31, 263–268. [Google Scholar] [CrossRef]
- Heide, O.M. Flowering responses of contrasting ecotypes of Poa annua and their putative ancestors Poa infirma and Poa supina. Ann. Bot. 2001, 87, 795–804. [Google Scholar] [CrossRef]
- Galera, H.; Rudak, A.; Czyż, E.A.; Chwedorzewska, K.J.; Znój, A.; Wódkiewicz, M. The role of the soil seed store in the survival of an invasive population of Poa annua at Point Thomas Oasis, King George Island, maritime Antarctica. Glob. Ecol. Conserv. 2019, 19, e00679. [Google Scholar] [CrossRef]
- Rudak, A.; Galera, H.; Znój, A.; Chwedorzewska, K.J.; Wódkiewicz, M. Seed germination and invasion success of Poa annua L. in Antarctica. Acta Soc. Bot. Pol. 2018, 87. [Google Scholar] [CrossRef]
- Williams, L.K.; Shaw, J.D.; Sindel, B.M.; Wilson, S.C.; Kristiansen, P. Longevity, growth and community ecology of invasive Poa annua across environmental gradients in the subantarctic. Basic Appl. Ecol. 2018, 29, 20–31. [Google Scholar] [CrossRef]
- Gardarin, A.; Dürr, C.; Colbach, N. Which model species for weed seedbank and emergence studies? A review. Weed Res. 2009, 49, 117–130. [Google Scholar] [CrossRef]
- Skottsberg, C. Antarctic vascular plants. Bot. Tidsskr. 1954, 51, 330–338. [Google Scholar]
- Chwedorzewska, K.J.; Giełwanowska, I.; Olech, M.; Molina-Montenegro, M.A.; Wódkiewicz, M.; Galera, H. Poa annua L. in the maritime Antarctic: An overview. Polar Rec. 2015, 51, 637–643. [Google Scholar] [CrossRef]
- Olech, M. Human impact on terrestrial ecosystems in west Antarctica. In Proceedings of the NIPR Symposium on Polar Biology; National Institute of Polar Research: Tokyo, Japan, 1996; pp. 299–306. [Google Scholar]
- Olech, M.; Chwedorzewska, K.J. Short note: The first appearance and establishment of an alien vascular plant in natural habitats on the forefield of a retreating glacier in Antarctica. Antarct. Sci. 2011, 23, 153. [Google Scholar] [CrossRef]
- Chwedorzewska, K.J.; Bednarek, P.T. Genetic and epigenetic variation in a cosmopolitan grass Poa annua from Antarctic and Polish populations. Pol. Polar Res. 2012, 33, 63–80. [Google Scholar] [CrossRef]
- Galera, H.; Chwedorzewska, K.J.; Łapiński, S.; Kowalska, M.E.; Pasik, M.; Rajner, M.; Czyż, E.; Bylina PWódkiewicz, M. First step to eradication of Poa annua L. from Arctowski oasis (King George Island, South Shetlands, Antarctica). Polar Biol. 2017, 40, 939–945. [Google Scholar] [CrossRef]
- De Zelicourt, A.; Al-Yousif, M.; Hirt, H. Rhizosphere microbes as essential partners for plant stress tolerance. Mol. Plant 2013, 6, 242–245. [Google Scholar] [CrossRef]
- Wei, Z.; Yang, T.; Friman, V.P.; Xu, Y.; Shen, Q.; Jousset, A. Trophic network architecture of root-associated bacterial communities determines pathogen invasion and plant health. Nat. Commun. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.R.; Ramakrishnan, K.; Walshaw, J.; Heavens, D.; Alston, M.; Swarbreck, D.; Poole, P.S. Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants. ISME J. 2013, 7, 2248–2258. [Google Scholar] [CrossRef] [PubMed]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef]
- Hartmann, A.; Rothballer, M.; Schmid, M. Lorenz Hiltner, a pioneer in rhizosphere microbial ecology and soil bacteriology research. Plant Soil 2008, 312, 7–14. [Google Scholar] [CrossRef]
- Trognitz, F.; Hackl, E.; Widhalm, S.; Sessitsch, A. The role of plant–microbiome interactions in weed establishment and control. FEMS Microbiol. Ecol. 2016, 92, fiw138. [Google Scholar] [CrossRef]
- Lunau, M.; Lemke, A.; Walther, K.; Martens-Habbena, W.; Simon, M. An improved method for counting bacteria from sediments and turbid environments by epifluorescence microscopy. Environ. Microbiol. 2005, 7, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, S.; Płociniczak, T.; Piotrowska-Seget, Z.; Złoch, M.; Ruppel, S.; Hrynkiewicz, K. Metabolic potential and community structure of endophytic and rhizosphere bacteria associated with the roots of the halophyte Aster tripolium L. Microbiol. Res. 2016, 182, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Weber, K.P.; Legge, R.L. One-dimensional metric for tracking bacterial community divergence using sole carbon source utilization patterns. J. Microbiol. Methods 2009, 79, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Moyer, C.L.; Morita, R.Y. Psychrophiles and psychrotrophs. eLS 2007. [Google Scholar] [CrossRef]
- Grzesiak, J.; Zdanowski, M.K.; Górniak, D.; Świątecki, A.; Aleksandrzak-Piekarczyk, T.; Szatraj, K.; Nieckarz, M. Microbial community changes along the ecology glacier ablation zone (King George Island, Antarctica). Polar Biol. 2015, 38, 2069–2083. [Google Scholar] [CrossRef]
- Grzesiak, J.; Kaczyńska, A.; Gawor, J.; Żuchniewicz, K.; Aleksandrzak-Piekarczyk, T.; Gromadka, R.; Zdanowski, M.K. A smelly business: Microbiology of Adélie penguin guano (Point Thomas rookery, Antarctica). Sci. Total Environ. 2020, 714, 136714. [Google Scholar] [CrossRef]
- Grzesiak, J.; Woltyńska, A.; Zdanowski, M.K.; Górniak, D.; Świątecki, A.; Olech, M.A.; Aleksandrzak-Piekarczyk, T. Metabolic fingerprinting of the Antarctic cyanolichen Leptogium puberulum–associated bacterial community (Western Shore of Admiralty Bay, King George Island, Maritime Antarctica). Microb. Ecol. 2021, 1–12. [Google Scholar] [CrossRef]
- Jones, J.B. Laboratory Guide for Conducting Soil Tests and Plant Analysis (No. BOOK); CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Tatur, A.; Myrcha, A.; Niegodzisz, J. Formation of abandoned penguin rookery ecosystems in the maritime Antarctic. Polar Biol. 1997, 17, 405–417. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 6 June 2020).
- Wynn-Williams, D.D. Antarctic microbial diversity: The basis of polar ecosystem processes. Biodivers. Conserv. 1996, 5, 1271–1293. [Google Scholar] [CrossRef]
- Convey, P.; Chown, S.L.; Clarke, A.; Barnes, D.K.; Bokhorst, S.; Cummings, V.; Griffiths, H.J. The spatial structure of Antarctic biodiversity. Ecol. Monogr. 2014, 84, 203–244. [Google Scholar] [CrossRef]
- Znój, A.; Chwedorzewska, K.J.; Androsiuk, P.; Cuba-Diaz, M.; Giełwanowska, I.; Koc, J.; Korczak-Abshire, M.; Grzesiak, J.; Zmarz, A. Rapid environmental changes in the Western Antarctic Peninsula region due to climate change and human activity. Appl. Ecol. Environ. Res. 2017, 15, 525–539. [Google Scholar] [CrossRef]
- Magalhães, C.; Stevens, M.I.; Cary, S.C.; Ball, B.A.; Storey, B.C.; Wall, D.H.; Ruprecht, U. At limits of life: Multidisciplinary insights reveal environmental constraints on biotic diversity in continental Antarctica. PLoS ONE 2012, 7, e44578. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.K.; Hodgson, D.A.; Convey, P.; Allen, C.S.; Clarke, A. Incursion and excursion of Antarctic biota: Past, present and future. Glob. Ecol. Biogeogr. 2006, 5, 121–142. [Google Scholar] [CrossRef]
- Smith, D.J.; Griffin, D.W.; McPeters, R.D.; Ward, P.D.; Schuerger, A.C. Microbial survival in the stratosphere and implications for global dispersal. Aerobiologia 2011, 27, 319–332. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Alberdi, M.; Bravo, L.A.; Gutiérrez, A.; Gidekel, M.; Corcuera, L.J. Ecophysiology of Antarctic vascular plants. Physiol. Plant 2002, 115, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.W.; Pearce, D.A.; Convey, P.; Yew, W.C.; Tan, I.K.P. Patterns in the distribution of soil bacterial 16S rRNA gene sequences from different regions of Antarctica. Geoderma 2012, 181, 45–55. [Google Scholar] [CrossRef]
- Navas, A.; López-Martínez, J.; Casas, J.; Machín, J.; Durán, J.J.; Serrano, E.; Mink, S. Soil characteristics on varying lithological substrates in the South Shetland Islands, maritime Antarctica. Geoderma 2008, 144, 123–139. [Google Scholar] [CrossRef]
- Hua SS, T.; Tsai, V.Y.; Lichens, G.M.; Noma, A.T. Accumulation of amino acids in Rhizobium sp. strain WR1001 in response to sodium chloride salinity. Appl. Environ. Microbiol. 1982, 44, 135–140. [Google Scholar] [PubMed]
- Elsheikh, E.A. Effects of salt on rhizobia and bradyrhizobia: A review. Ann. Appl. Biol. 1998, 132, 507–524. [Google Scholar] [CrossRef]
- Broos, K.; Beyens, H.; Smolders, E. Survival of rhizobia in soil is sensitive to elevated zinc in the absence of the host plant. Soil Biol. Biochem. 2005, 37, 573–579. [Google Scholar] [CrossRef]
- Buckley, D.H.; Huangyutitham, V.; Nelson, T.A.; Rumberger, A.; Thies, J.E. Diversity of planctomycetes in soil in relation to soil history and environmental heterogeneity. Appl. Environ. Microbiol. 2006, 72, 4522–4531. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.M.C.; de Souza, J.A.M.; de Mello Varani, A.; de Macedo Lemos, E.G. The Family Rhizobiaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 419–437. [Google Scholar]
- Liu, Y.; Ludewig, U. Nitrogen dependent bacterial community shifts in root, rhizome and rhizosphere of nutrient efficient Miscanthus x giganteus from long term field trials. GCB Bioenergy 2019, 11, 1334–1347. [Google Scholar] [CrossRef]
- Schlatter, D.C.; Yin, C.; Hulbert, S.; Paulitz, T.C. Core rhizosphere microbiomes of dryland wheat are influenced by location and land use history. Appl. Environ. Microbiol. 2020, 86, e02135-19. [Google Scholar] [CrossRef]
- Da Rocha, U.N.; van Elsas, J.D.; van Overbeek, L.S. Real-time PCR detection of Holophagae (Acidobacteria) and Verrucomicrobia subdivision 1 groups in bulk and leek (Allium porrum) rhizosphere soils. J. Microbiol. Methods 2010, 83, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Gonzalez, P.A.; Bornemann, T.L.; Adam, P.S.; Plewka, J.; Révész, F.; von Hagen, C.A.; Probst, A.J. Saccharibacteria as organic carbon sinks in hydrocarbon-fueled communities. Front. Microbiol. 2020, 11, 3343. [Google Scholar] [CrossRef] [PubMed]
- Nuccio, E.E.; Anderson Furgeson, J.; Estera, K.Y.; Pett Ridge, J.; de Valpine, P.; Brodie, E.L.; Firestone, M.K. Climate and edaphic controllers influence rhizosphere community assembly for a wild annual grass. Ecology 2016, 97, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Wiegel, J.; Tanner, R.; Rainey, F.A. An introduction to the family Clostridiaceae. Prokaryotes 2006, 4, 654–678. [Google Scholar]
- Pudełko, R.; Angiel, P.J.; Potocki, M.; Jędrejek, A.; Kozak, M. Fluctuation of glacial retreat rates in the eastern part of Warszawa Icefield, King George Island, Antarctica, 1979–2018. Remote Sens. 2018, 10, 892. [Google Scholar] [CrossRef]
- Zdanowski, M.K.; Bogdanowicz, A.; Gawor, J.; Gromadka, R.; Wolicka, D.; Grzesiak, J. Enrichment of cryoconite hole anaerobes: Implications for the subglacial microbiome. Microb. Ecol. 2017, 73, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Barret, M.; Morrissey, J.P.; O’gara, F. Functional genomics analysis of plant growth-promoting rhizobacterial traits involved in rhizosphere competence. Biol. Fertil. Soils 2011, 47, 729. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo-and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef]
- Fournier, J.; Teillet, A.; Chabaud, M.; Ivanov, S.; Genre, A.; Limpens, E.; Barker, D.G. Remodeling of the infection chamber before infection thread formation reveals a two-step mechanism for rhizobial entry into the host legume root hair. Plant Physiol. 2015, 167, 1233–1242. [Google Scholar] [CrossRef]
- Tian, X.Y.; Zhang, C.S. Illumina-based analysis of endophytic and rhizosphere bacterial diversity of the coastal halophyte Messerschmidia sibirica. Front. Microbiol. 2017, 8, 2288. [Google Scholar] [CrossRef] [PubMed]
- Moronta-Barrios, F.; Gionechetti, F.; Pallavicini, A.; Marys, E.; Venturi, V. Bacterial microbiota of rice roots: 16S-based taxonomic profiling of endophytic and rhizospheric diversity, endophytes isolation and simplified endophytic community. Microorganisms 2018, 6, 14. [Google Scholar] [CrossRef]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef] [PubMed]
- Cocking, E.C. Endophytic colonization of plant roots by nitrogen-fixing bacteria. Plant Soil 2003, 252, 169–175. [Google Scholar] [CrossRef]
- Bahulikar, R.A.; Chaluvadi, S.R.; Torres-Jerez, I.; Mosali, J.; Bennetzen, J.L.; Udvardi, M. Nitrogen fertilization reduces nitrogen fixation activity of diverse diazotrophs in switchgrass roots. Phytobiomes J. 2020. [Google Scholar] [CrossRef]
- Jiang, Z.K.; Tuo, L.; Huang, D.L.; Osterman, I.A.; Tyurin, A.P.; Liu, S.W.; Li, F.N. Diversity, novelty, and antimicrobial activity of endophytic actinobacteria from mangrove plants in Beilun Estuary National Nature Reserve of Guangxi, China. Front. Microbiol. 2018, 9, 868. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Takahashi, Y. Endophytic actinomycetes: Promising source of novel bioactive compounds. J. Antibiot. 2017, 70, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Acuña, J.J.; Inostroza, N.G.; Duran, P.; Mora, M.L.; Sadowsky, M.J.; Jorquera, M.A. Niche differentiation in the composition, predicted function, and co-occurrence networks in bacterial communities associated with Antarctic vascular plants. Front. Microbiol. 2020, 11, 1036. [Google Scholar] [CrossRef] [PubMed]
- Ofek, M.; Hadar, Y.; Minz, D. Ecology of root colonizing Massilia (Oxalobacteraceae). PLoS ONE 2012, 7, e40117. [Google Scholar] [CrossRef]
- Zdanowski, M.K.; Żmuda-Baranowska, M.J.; Borsuk, P.; Świątecki, A.; Górniak, D.; Wolicka, D.; Grzesiak, J. Culturable bacteria community development in postglacial soils of Ecology Glacier, King George Island, Antarctica. Polar Biol. 2013, 36, 511–527. [Google Scholar] [CrossRef]
- Gustavs, L.; Görs, M.; Karsten, U. Polyol patterns in biofilm forming aeroterrestrial green algae (Trebouxiophyceae, Chlorophyta) 1. J. Phycol. 2011, 47, 533–537. [Google Scholar] [CrossRef]
- Kolton, M.; Erlacher, A.; Berg, G.; Cytryn, E. The Flavobacterium genus in the plant holobiont: Ecological, physiological, and applicative insights. In Microbial Models: From Environmental to Industrial Sustainability; Springer: Singapore, 2016; pp. 189–207. [Google Scholar]
- Molina-Montenegro, M.A.; Ballesteros, G.I.; Castro-Nallar, E.; Meneses, C.; Gallardo-Cerda, J.; Torres-Díaz, C. A first insight into the structure and function of rhizosphere microbiota in Antarctic plants using shotgun metagenomic. Polar Biol. 2019, 42, 1825–1835. [Google Scholar] [CrossRef]
- Maimaiti, J.; Zhang, Y.; Yang, J.; Cen, Y.P.; Layzell, D.B.; Peoples, M.; Dong, Z. Isolation and characterization of hydrogen oxidizing bacteria induced following exposure of soil to hydrogen gas and their impact on plant growth. Environ. Microbiol. 2007, 9, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Coats, V.C.; Rumpho, M.E. The rhizosphere microbiota of plant invaders: An overview of recent advances in the microbiomics of invasive plants. Front. Microbiol. 2014, 5, 368. [Google Scholar] [CrossRef] [PubMed]
- Galera, H.; Chwedorzewska, K.J.; Wódkiewicz, M. Response of Poa annua to extreme conditions: Comparison of morphological traits between populations from cold and temperate climate conditions. Polar Biol. 2015, 38, 1657–1666. [Google Scholar] [CrossRef]
- Vega-Celedón, P.; Bravo, G.; Velásquez, A.; Cid, F.P.; Valenzuela, M.; Ramírez, I.; Seeger, M. Microbial diversity of psychrotolerant bacteria isolated from wild flora of andes mountains and patagonia of chile towards the selection of plant growth-promoting bacterial consortia to alleviate cold stress in plants. Microorganisms 2021, 9, 538. [Google Scholar] [CrossRef] [PubMed]
- Tiryaki, D.; Aydın, İ.; Atıcı, Ö. Psychrotolerant bacteria isolated from the leaf apoplast of cold-adapted wild plants improve the cold resistance of bean (Phaseolus vulgaris L.) under low temperature. Cryobiology 2019, 86, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Araya, M.A.; Valenzuela, T.; Inostroza, N.G.; Maruyama, F.; Jorquera, M.A.; Acuña, J.J. Isolation and characterization of cold-tolerant hyper-ACC-degrading bacteria from the rhizosphere, endosphere, and phyllosphere of antarctic vascular plants. Microorganisms 2020, 8, 1788. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Gao, L. Bacterial diversity in the rhizosphere of cucumbers grown in soils covering a wide range of cucumber cropping histories and environmental conditions. Microb. Ecol. 2014, 68, 794–806. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Geographical Coordinates | Distance to the Sea | Altitude | Structure of Vegetation | Landform and Habitat |
|---|---|---|---|---|---|
| P1 | 52°06′22″ N 21°05′56″ E Powsin, Poland | 285 km | 90 m.a.s.l. | Poa annua (90%) and Poa pratensis (10%). | Rubble path, sand, and sandstone. Dry site, sheltered away from sources of nutrients. |
| P2 | 52°06′29″ N 21°05′33″ E Powsin, Poland | 285 km | 95 m.a.s.l. | Poa annua (30%), Malus domestica (10%), Trifolium arvense (30%), Sonchus oleraceus L. (10%), Sonchus arvensis (20%). | Soil, mechanically altered by human activities. Wet site, fertile, extra fertilization once a year with an animal post-food mass. |
| P3 | 62°09′35″ S 58°28′26″ W Arctowski Station, Antarctica | 120 m | 0.5 m.a.s.l., | Colobanthus quitensis (15%), Deschampsia antarctica (25%), Poa annua (10%), mosses (25%) and fruticose, foliose and crustose lichens (25%). | Base station, soil mechanically altered by human activities; Skeletic Eutric Fluvisol (Turbic) The site is strongly influenced by marine aerosols, moist, with large human influence. |
| P4 | 62°10′05″ S 58°27′46″ W Ecology Glacier foreland, Antarctica | 20 m | 0.5 m.a.s.l., | Mosses (40%), fruticose and foliose lichens (40%), Colobanthus quitensis (10%) and Deschampsia antarctica (10%). | Fluted moraine; Eutric Skeletic Protic Regosol (Turbic). Dry site, sheltered away from sources of nutrients with a little influence of marine aerosols. |
| P5 | 62°09′33″ S 58°28′25″ W Arctowski Station, Antarctica | 100 m | 0.5 m.a.s.l., | Colobanthus quitensis (15%), Deschampsia antarctica (25%), Poa annua (10%), mosses (25%) and fruticose, foliose and crustose lichens (25%). | Base station, soil, mechanically altered by human activities; Skeletic Eutric Fluvisol (Turbic) The site is medium influenced by marine aerosols, moist, with large human influence, especially by using big vehicles. |
| Soil Components | S1 | S2 | S3 | S4 | S5 |
|---|---|---|---|---|---|
| N (mgNO3/100 g) | 17.4 | 27.5 | 8.8 | 16.3 | 7.7 |
| P (mgP2O5/100 g) | 6.0 | 219.2 | 5.6 | 15.1 | 4.9 |
| K (mg K2O/100 g) | 36.1 | 252.8 | 31.1 | 55.8 | 31.9 |
| Mg (mg Mg/100 g) | 38.4 | 185.4 | 244.7 | 205 | 250.7 |
| Ca (mg Ca/100 g) | >5000 | 2024.6 | 498.0 | 1020.1 | 533.3 |
| Na (mg Na/100 g) | 4.9 | 4.0 | >150 | >150 | >150 |
| Salinity (g NaCl/L) | 0.2 | 0.3 | 0.4 | 0.4 | 0.4 |
| pH | 7.8 | 7.0 | 7.4 | 7.5 | 7.4 |
| Mn (mg Mn/kg) | 52.8 | 218.1 | 147.5 | 132.1 | 155.5 |
| Zn (mg Zn/kg) | 5.9 | >50 | >50 | 9.2 | >50 |
| Cu (mg Cu/kg) | 2.7 | 5.7 | 21.8 | >30 | 29.8 |
| Fe (mg Fe/kg) | 389.4 | 1502.5 | 4412.6 | 4670.2 | 4269.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Znój, A.; Grzesiak, J.; Gawor, J.; Gromadka, R.; Chwedorzewska, K.J. Bacterial Communities Associated with Poa annua Roots in Central European (Poland) and Antarctic Settings (King George Island). Microorganisms 2021, 9, 811. https://doi.org/10.3390/microorganisms9040811
Znój A, Grzesiak J, Gawor J, Gromadka R, Chwedorzewska KJ. Bacterial Communities Associated with Poa annua Roots in Central European (Poland) and Antarctic Settings (King George Island). Microorganisms. 2021; 9(4):811. https://doi.org/10.3390/microorganisms9040811
Chicago/Turabian StyleZnój, Anna, Jakub Grzesiak, Jan Gawor, Robert Gromadka, and Katarzyna J. Chwedorzewska. 2021. "Bacterial Communities Associated with Poa annua Roots in Central European (Poland) and Antarctic Settings (King George Island)" Microorganisms 9, no. 4: 811. https://doi.org/10.3390/microorganisms9040811
APA StyleZnój, A., Grzesiak, J., Gawor, J., Gromadka, R., & Chwedorzewska, K. J. (2021). Bacterial Communities Associated with Poa annua Roots in Central European (Poland) and Antarctic Settings (King George Island). Microorganisms, 9(4), 811. https://doi.org/10.3390/microorganisms9040811