
Probiotic Bacillus safensis NPUST1 Administration Improves Growth Performance, Gut Microbiota, and Innate Immunity against Streptococcus iniae in Nile tilapia (Oreochromis niloticus)




Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Husbandry and Bacterial Strains
2.2. Isolation and Characterization of Potential Probiotics
2.3. Diet Preparation
2.4. Feeding Trail
2.5. Growth Performance Parameters
2.6. Assessment of the Digestive Enzyme Activities in Gut
2.7. Assessment of the Innate Immune Parameters
2.8. Challenge Experiment
2.9. Intestinal Microbiota Analysis
2.10. Statistical Analysis
3. Results
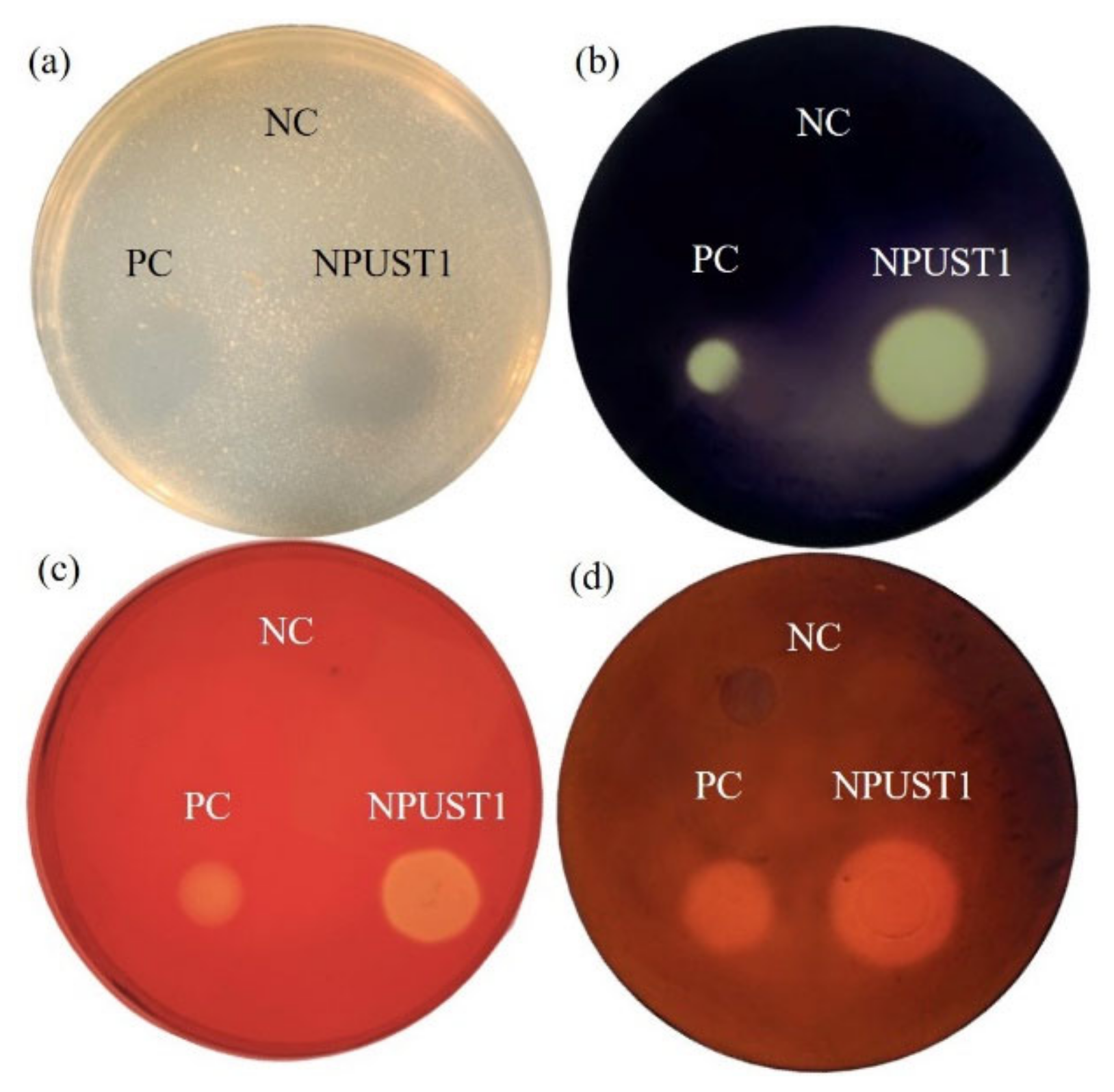
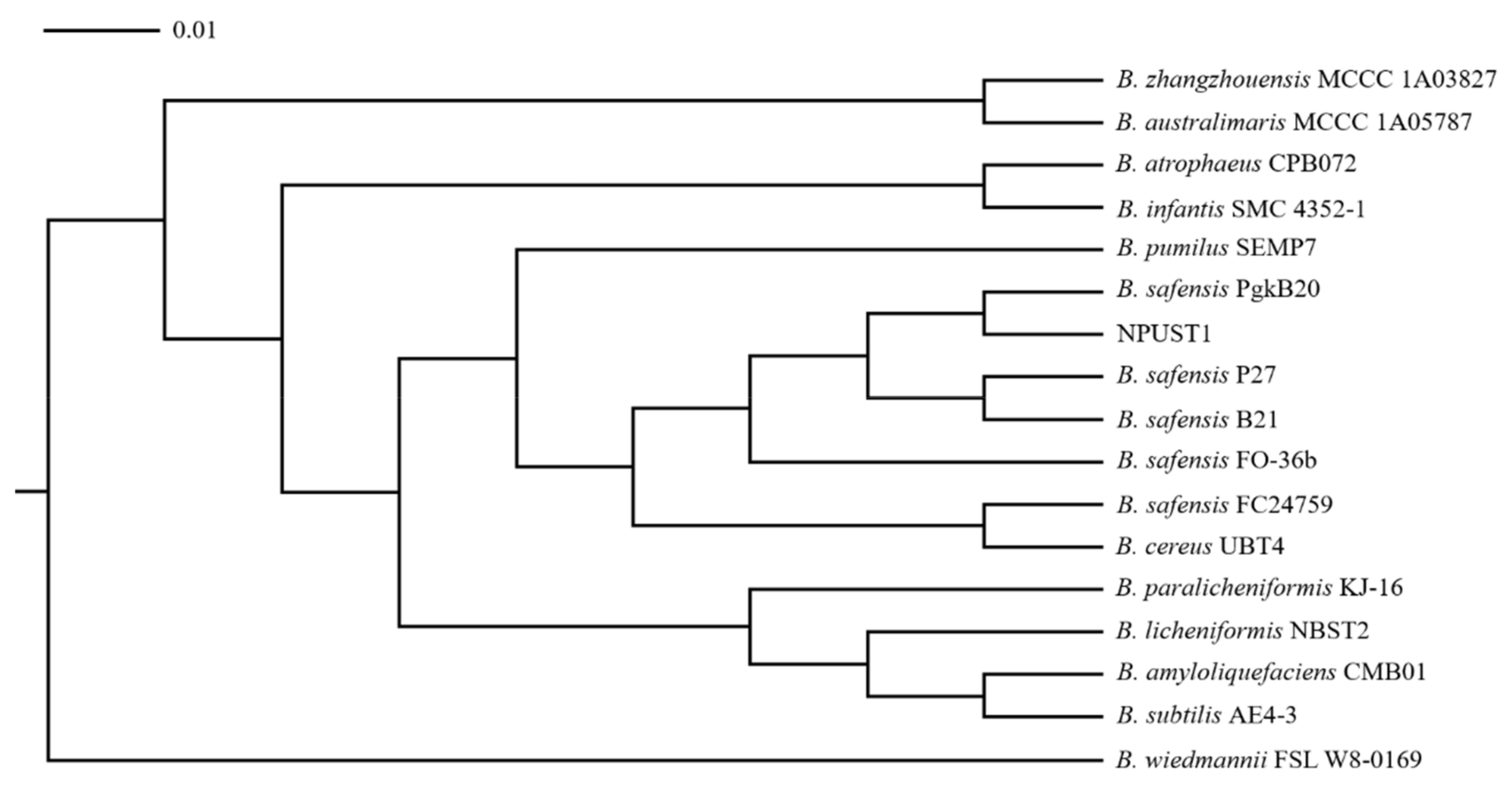
3.1. Isolation of the Potential Probiotics with Hydrolytic Enzyme Activities
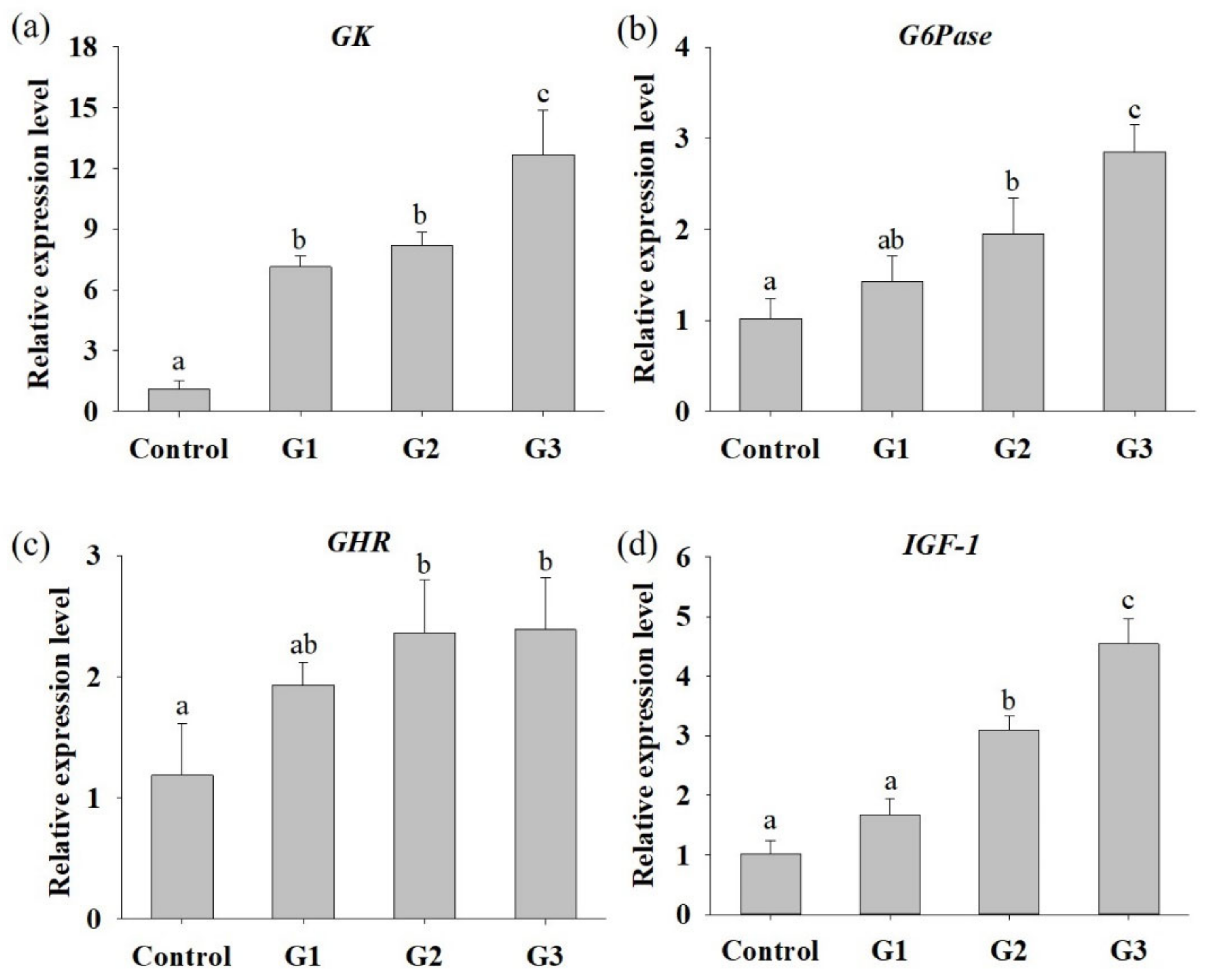
3.2. B. safensis NPUST1 Supplementation Enhances Nutrient Utilization and Growth Performance
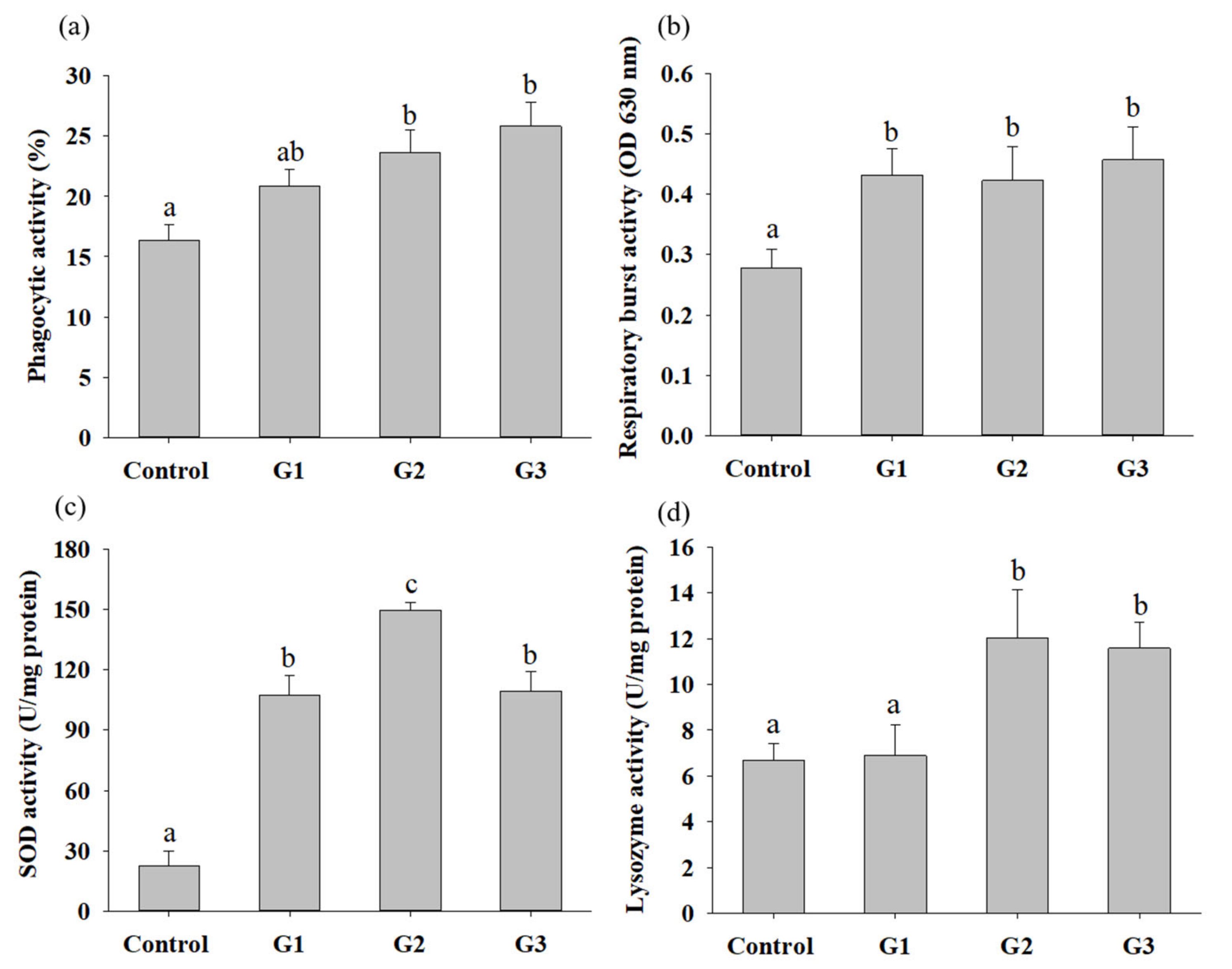
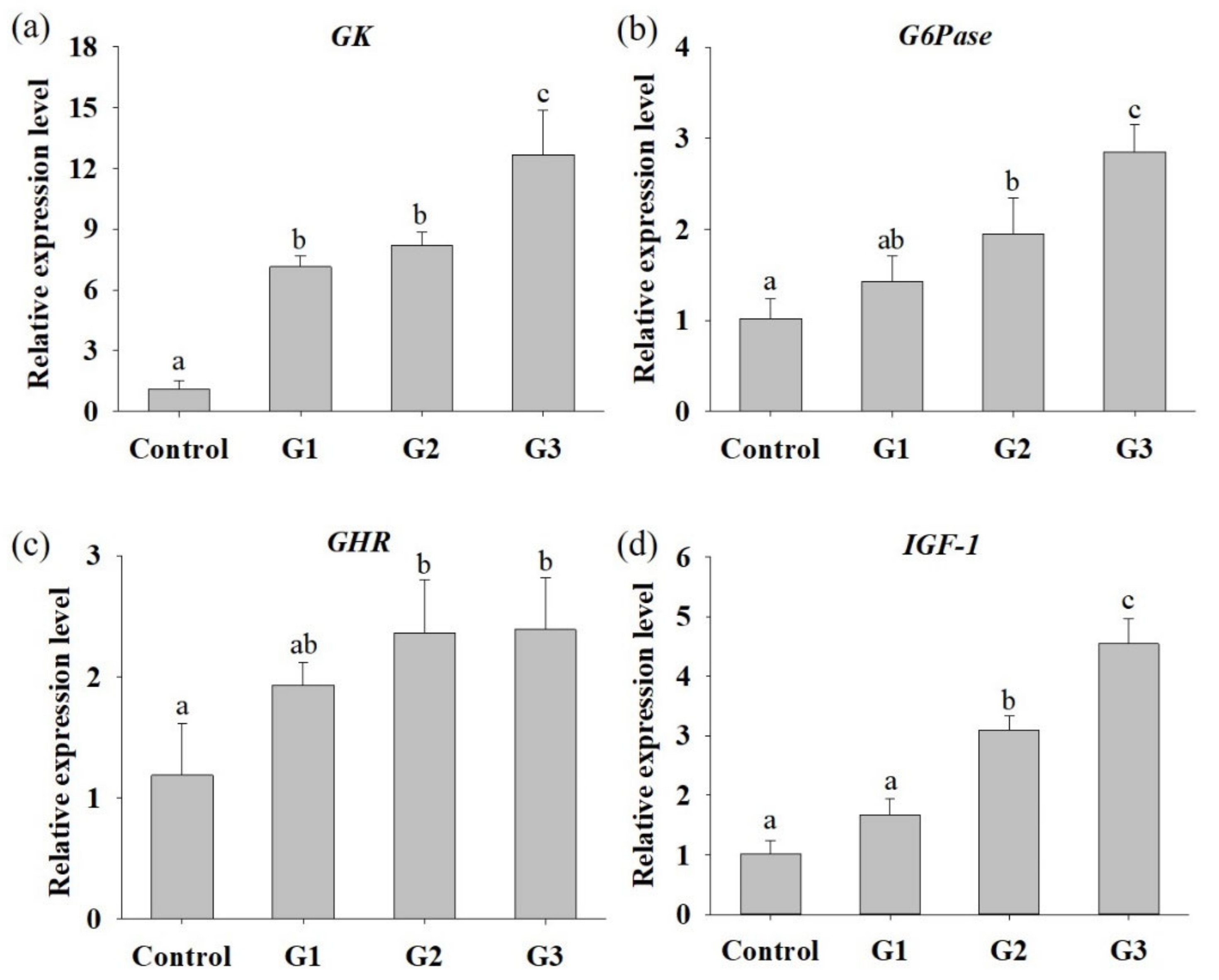
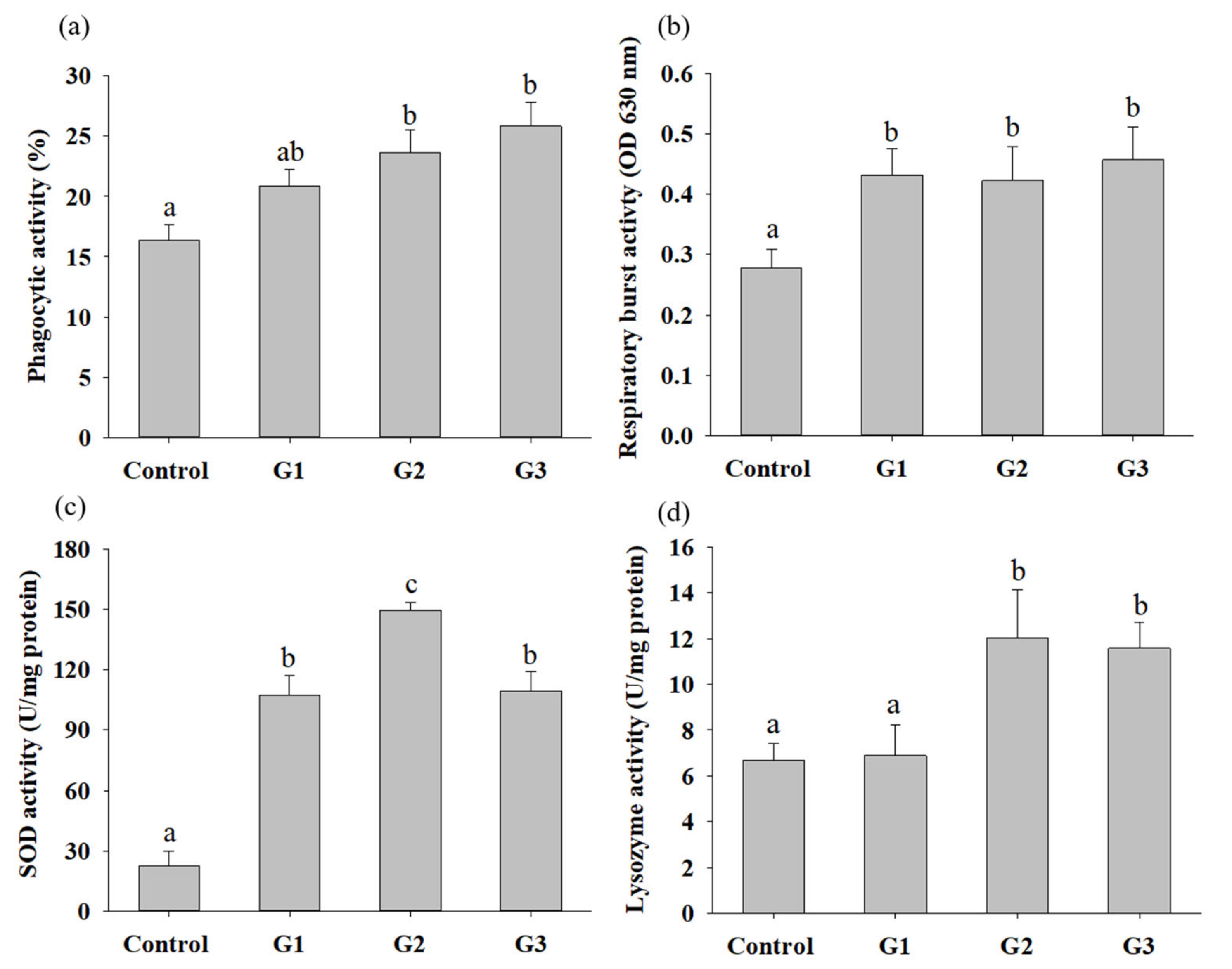
3.3. B. safensis NPUST1 Supplementation Enhances Innate Immunity
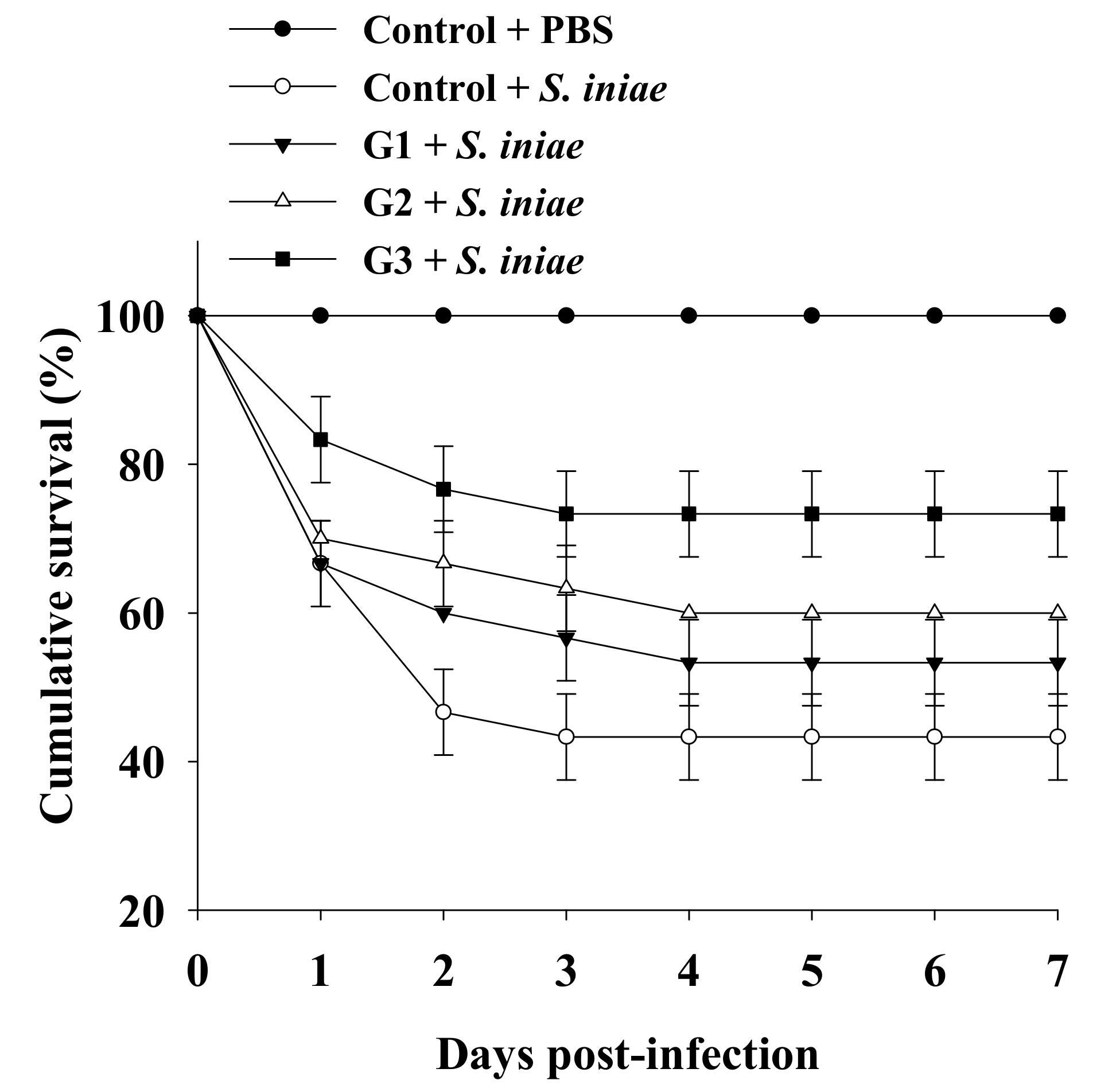
3.4. Dietary B. safensis NPUST1 Enhances Defense against S. iniae Infection in Tilapia
3.5. Dietary B. safensis NPUST1 Improve Intestinal Microbiota
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of Fisheries and Aquaculture. In Sustainability in Action; FAO: Rome, Italy, 2020; Available online: https://www.fao.org/documents/card/en/c/ca9229en (accessed on 30 November 2021).
- Ortega, C.; Garcia, I.; Irgang, R.; Fajardo, R.; Tapia-Cammas, D.; Acosta, J.; Avendano-Herrera, R. First identification and characterization of Streptococcus iniae obtained from tilapia (Oreochromis aureus) farmed in Mexico. J. Fish Dis. 2018, 41, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Rahmatullah, M.; Ariff, M.; Kahieshesfandiari, M.; Daud, H.M.; Zamri-Saad, M.; Sabri, M.Y.; Amal, M.N.A.; Ina-Salwany, M.Y. Isolation and pathogenicity of Streptococcus iniae in cultured red hybrid Tilapia in Malaysia. J. Aquat Anim. Health 2017, 29, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.; Wang, J.; Ji, X.; Yang, H.; Tang, B.; Zhang, H.; Yang, G.; Bao, Z.; Jin, Y. Sub-chronic exposure to antibiotics doxycycline, oxytetracycline or florfenicol impacts gut barrier and induces gut microbiota dysbiosis in adult zebrafish (Daino rerio). Ecotoxicol. Environ. Saf. 2021, 221, 112464. [Google Scholar] [CrossRef]
- Kozasa, M. Toyocerin (Bacillus toyoi) as growth promotor for animal feeding. Microbiol. Aliments Nutr. 1986, 4, 121–135. [Google Scholar]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [Green Version]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y.; Hlordzi, V.; Sakyi, M.E.; Afriyie, G.; Wang, Z.; Li, Y.; Xie, C.X. Mechanisms and the role of probiotic Bacillus in mitigating fish pathogens in aquaculture. Fish Physiol. Biochem. 2020, 46, 819–841. [Google Scholar] [CrossRef]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Shellfish Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Benson, A.; Lebeer, S.; Merenstein, D.J.; Klaenhammer, T.R. Shared mechanisms among probiotic taxa: Implications for general probiotic claims. Curr. Opin. Biotechnol. 2018, 49, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Satomi, M.; La Duc, M.T.; Venkateswaran, K. Bacillus safensis sp. nov., isolated from spacecraft and assembly-facility surfaces. Int. J. Syst. Evol. Microbiol. 2006, 56, 1735–1740. [Google Scholar] [CrossRef] [PubMed]
- Lateef, A.; Adelere, I.A.; Gueguim-Kana, E.B. The biology and potential biotechnological applications of Bacillus safensis. Biologia 2015, 70, 411–419. [Google Scholar] [CrossRef]
- Ficarra, F.A.; Santecchia, I.; Lagorio, S.H.; Alarcon, S.; Magni, C.; Espariz, M. Genome mining of lipolytic exoenzymes from Bacillus safensis S9 and Pseudomonas alcaliphila ED1 isolated from a dairy wastewater lagoon. Arch. Microbiol. 2016, 198, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Rekik, H.; Jaouadi, N.Z.; Gargouri, F.; Bejar, W.; Frikha, F.; Jmal, N.; Bejar, S.; Jaouadi, B. Production, purification and biochemical characterization of a novel detergent-stable serine alkaline protease from Bacillus safensis strain RH12. Int. J. Biol. Macromol. 2019, 121, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Thite, V.S.; Nerurkar, A.S.; Baxi, N.N. Optimization of concurrent production of xylanolytic and pectinolytic enzymes by Bacillus safensis M35 and Bacillus altitudinis J208 using agro-industrial biomass through response surface methodology. Sci. Rep. 2020, 10, 3824. [Google Scholar] [CrossRef]
- Walsh, G.A.; Power, R.F.; Headon, D.R. Enzymes in the animal-feed industry. Trends Biotechnol. 1993, 11, 424–430. [Google Scholar] [CrossRef]
- Tan, H.Y.; Chen, S.W.; Hu, S.Y. Improvements in the growth performance, immunity, disease resistance, and gut microbiota by the probiotic Rummeliibacillus stabekisii in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019, 92, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Saputra, F.; Shiu, Y.L.; Chen, C.Y.; Puspitasari, A.W.; Danata, R.H.; Liu, C.H.; Hu, S.Y. Dietary supplementation with xylanase-expressing B. amyloliquefaciens R8 improves growth performance and enhances immunity against Aeromonas hydrophila in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2016, 58, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, S.; Cai, Y.; Guo, X.; Cao, Z.; Zhang, Y.; Liu, S.; Yuan, W.; Zhu, W.; Zheng, Y.; et al. Dietary administration of Bacillus subtilis HAINUP40 enhances growth, digestive enzyme activities, innate immune responses and disease resistance of tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2017, 60, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Bairagi, A.; Ghosh, K.S.; Sen, S.K.; Ray, A.K. Evaluation of the nutritive value of Leucaena leucocephala leaf meal, inoculated with fish intestinal bacteria Bacillus subtilis and Bacillus circulans in formulated diets for rohu, Labeo rohita (Hamilton) fingerlings. Aquac. Res. 2004, 35, 436–446. [Google Scholar] [CrossRef]
- Zhang, M.; Pan, L.; Fan, D.; He, J.; Su, C.; Gao, S.; Zhang, M. Study of fermented feed by mixed strains and their effects on the survival, growth, digestive enzyme activity and intestinal flora of Penaeus vannamei. Aquaculture 2021, 530, 735703. [Google Scholar] [CrossRef]
- Chen, S.W.; Liu, C.H.; Hu, S.Y. Dietary administration of probiotic Paenibacillus ehimensis NPUST1 with bacteriocin-like activity improves growth performance and immunity against Aeromonas hydrophila and Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019, 84, 695–703. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analyses, 16th ed.; Association of official Analytical Chemists: Washington, DC, USA, 1997. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.T.; Peng, X.; Deng, G.H.; Sheng, H.F.; Wang, Y.; Zhou, H.W.; Tam, N.F. Illumina sequencing of 16S rRNA tag revealed spatial variations of bacterial communities in a mangrove wetland. Microb. Ecol. 2013, 66, 96–104. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Olmos, J.; Acosta, M.; Mendoza, G.; Pitones, V. Bacillus subtilis, an ideal probiotic bacterium to shrimp and fish aquaculture that increase feed digestibility, prevent microbial diseases, and avoid water pollution. Arch. Microbiol. 2020, 202, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Docando, F.; Nunez-Ortiz, N.; Tafalla, C.; Diaz-Rosales, P. Mechanisms used by probiotics to confer pathogen resistance to teleost fish. Front. Immunol. 2021, 12, 653025. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K.; Kumar, V.; Makkar, H.P.S.; Boeck, G.D.; Becker, K. Non-starch polysaccharides and their role in fish nutrition – A review. Food Chem. 2011, 127, 1409–1426. [Google Scholar] [CrossRef]
- Maas, R.M.; Verdegem, M.C.J.; Schrama, J.W. Effect of non-starch polysaccharide composition and enzyme supplementation on growth performance and nutrient digestibility in Nile tilapia (Oreochromis niloticus). Aquac. Nutr. 2019, 25, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Luo, Z.; Chen, F.; Huang, C.; Zhu, X.M.; Liu, X. Effects of dietary cellulase addition on growth performance, nutrient digestibility and digestive enzyme activities of juvenile crucian carp Carassius auratus. Aquac. Nutr. 2017, 23, 618–628. [Google Scholar] [CrossRef]
- Zuo, Z.H.; Shang, B.J.; Shao, Y.C.; Li, W.Y.; Sun, J.S. Screening of intestinal probiotics and the effects of feeding probiotics on the growth, immune, digestive enzyme activity and intestinal flora of Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 86, 160–168. [Google Scholar] [CrossRef]
- Yi, C.C.; Liu, C.H.; Chuang, K.P.; Chang, Y.T.; Hu, S.Y. A potential probiotic Chromobacterium aquaticum with bacteriocin-like activity enhances the expression of indicator genes associated with nutrient metabolism, growth performance and innate immunity against pathogen infections in zebrafish (Danio rerio). Fish Shellfish Immunol. 2019, 93, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Saputra, F.; Chen, Y.C.; Hu, S.Y. Dietary administration of Bacillus amyloliquefaciens R8 reduces hepatic oxidative stress and enhances nutrient metabolism and immunity against Aeromonas hydrophila and Streptococcus agalactiae in zebrafish (Danio rerio). Fish Shellfish Immunol. 2019, 86, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Meidong, R.; Khotchanalekha, K.; Doolgindachbaporn, S.; Nagasawa, T.; Nakao, M.; Sakai, K.; Tongpim, S. Evaluation of probiotic Bacillus aerius B81e isolated from healthy hybrid catfish on growth, disease resistance and innate immunity of Pla-mong Pangasius bocourti. Fish Shellfish Immunol. 2018, 73, 1–10. [Google Scholar] [CrossRef]
- Biller, J.D.; Takahashi, L.S. Oxidative stress and fish immune system: Phagocytosis and leukocyte respiratory burst activity. An. Acad. Bras. Cienc. 2018, 90, 3403–3414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, e1006512. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Austin, B. Cytokine expression in leucocytes and gut cells of rainbow trout, Oncorhynchus mykiss Walbaum, induced by probiotics. Vet. Immunol. Immunopathol. 2006, 114, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Adeshina, I.; Abubakar, M.I.; Ajala, B.E. Dietary supplementation with Lactobacillus acidophilus enhanced the growth, gut morphometry, antioxidant capacity, and the immune response in juveniles of the common carp, Cyprinus carpio. Fish Physiol. Biochem. 2020, 46, 1375–1385. [Google Scholar] [CrossRef]
- Beck, B.R.; Kim, D.; Jeon, J.; Lee, S.M.; Kim, H.K.; Kim, O.J.; Lee, J.I.; Suh, B.S.; Do, H.K.; Lee, K.H.; et al. The effects of combined dietary probiotics Lactococcus lactis BFE920 and Lactobacillus plantarum FGL0001 on innate immunity and disease resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2015, 42, 177–183. [Google Scholar] [CrossRef]
- Butt, R.L.; Volkoff, H. Gut microbiota and energy homeostasis in fish. Front. Endocrinol. 2019, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Opiyo, M.A.; Jumbe, J.; Ngugi, C.C.; Charo-Karisa, H. Dietary administration of probiotics modulates non-specific immunity and gut microbiota of Nile tilapia (Oreochromis niloticus) cultured in low input ponds. Int. J. Vet. Sci. Med. 2019, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kuebutornye, F.K.A.; Wang, Z.; Lu, Y.; Abarike, E.D.; Sakyi, M.E.; Li, Y.; Xie, C.X.; Hlordzi, V. Effects of three host-associated Bacillus species on mucosal immunity and gut health of Nile tilapia, Oreochromis niloticus and its resistance against Aeromonas hydrophila infection. Fish Shellfish Immunol. 2020, 97, 83–95. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, M.; Gao, F.; Lu, M.; Chen, G. Effects of dietary probiotic supplementation on the growth, gut health and disease resistance of juvenile Nile tilapia (Oreochromis niloticus). Anim. Nutr. 2020, 6, 69–79. [Google Scholar] [CrossRef]
- Jung, A.; Jung-Schroers, V. Detection of Deefgea chitinilytica in freshwater ornamental fish. Lett. Appl. Microbiol. 2011, 52, 497–500. [Google Scholar] [CrossRef]
- Yu, L.; Qiao, N.; Li, T.; Yu, R.; Zhai, Q.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Dietary supplementation with probiotics regulates gut microbiota structure and function in Nile tilapia exposed to aluminum. PeerJ 2019, 7, e6963. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Lu, M.; Chen, G.; Cao, J.; Gao, F.; Wang, M.; Liu, Z.; Zhang, D.; Zhu, H.; Yi, M. Effects of dietary Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp. lactis JCM5805 on the growth, intestinal microbiota, morphology, immune response and disease resistance of juvenile Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2018, 76, 368–379. [Google Scholar] [CrossRef]
- Dong, H.T.; Techatanakitarnan, C.; Jindakittikul, P.; Thaiprayoon, A.; Taengphu, S.; Charoensapsri, W.; Khunrae, P.; Rattanarojpong, T.; Senapin, S. Aeromonas jandaei and Aeromonas veronii caused disease and mortality in Nile tilapia, Oreochromis niloticus (L.). J. Fish Dis. 2017, 40, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Austin, D.; Sutherland, R.; Thompson, F.; Swings, J. Pathogenicity of vibrios to rainbow trout (Oncorhynchus mykiss, Walbaum) and Artemia nauplii. Environ. Microbiol. 2005, 7, 1488–1495. [Google Scholar] [CrossRef]
- Wu, F.S.; Tang, K.H.; Yuan, M.; Shi, X.C.; Shakeela, Q.; Zhang, X.H. Studies on bacterial pathogens isolated from diseased torafugu (Takifugu rubripes) cultured in marine industrial recirculation aquaculture system in Shandong Province, China. Aquac. Res. 2015, 46, 736–744. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Treatment (g kg−1) | |||
|---|---|---|---|---|
| Control Group | G1 Group | G2 Group | G3 Group | |
| B. safensis NPUST1 | 0 | 108 CFU | 109 CFU | 1010 CFU |
| Fish meal | 50 | 50 | 50 | 50 |
| Soybean meal | 300 | 300 | 300 | 300 |
| Wheat middling | 161 | 161 | 161 | 161 |
| Rice bran | 300 | 300 | 300 | 300 |
| α-starch | 60 | 60 | 60 | 60 |
| Cellulose | 70 | 70 | 70 | 70 |
| Skim milk | 10 | 10 | 10 | 10 |
| Soybean oil | 30 | 30 | 30 | 30 |
| Mineral mixture a | 16 | 16 | 16 | 16 |
| Vitamin mixture a | 3 | 3 | 3 | 3 |
| Proximate composition | ||||
| Crude protein | 236.1 | 234.0 | 236.1 | 230.1 |
| Crude lipid | 103.5 | 104.8 | 102.1 | 101.6 |
| Moisture | 96.8 | 96.2 | 93.8 | 96.5 |
| Ash | 65.9 | 64.9 | 66.1 | 64.2 |
| Gene Name | Primer Sequences (5′→3′) | Amplicon Size | Accession No. |
|---|---|---|---|
| Glucose kinase (GK) | F: GCAGCGAGGAAGCCATGAAGA R: GAGGTCCCTGACGACTTTGTGG | 101 bp | XM_003451020 |
| Glucose-6-phosphatease (G6 Pase) | F: AGCGCGAGCCTGAAGAAGTACT R: ATGGTCCACAGCAGGTCCACAT | 107 bp | XM_003448671 |
| Growth hormone receptor-1 (GHR-1) | F: GAATACAAGTCCTTCCGGGCTAA R: CTCATACTCCACACGCATCCA | 100 bp | AY973232 |
| Insulin-like growth factor-1 (IGF-1) | F: TGTCTGCCAGTAAGGATGTTCTTG R: GGCTTTCCACGCCACTTAAC | 100 bp | EU272149 |
| Tumor necrosis factor (TNF-α) | F: CCAGAAGCACTAAAGGCGAAGA R: CCTTGGCTTTGCTGCTGATC | 82 bp | AY428948 |
| Interleukin-1β (ΙL-1β) | F: TGTCGCTCTGGGCATCAA R: GGCTTGTCGTCATCCTTGTGA | 63 bp | KJ574402 |
| Interleukin-8 (IL-8) | F: CCTCGAGAAGGTGGATGTGAA R: CATGAGACCCAGGGCATCA | 100 bp | GQ355864 |
| Lysozyme (Lyz) | F: GCCTGACCGAACATGAGTCA R: CACCAGCGGCTATTTATCTGAA | 100 bp | LC012581 |
| β-actin | F: CCACACAGTGCCCATCTACGA R: CCACGCTCTGTCAGGATCTTCA | 111 bp | EU887951 |
| Parameters | Administrations | |||
|---|---|---|---|---|
| Control | 105 CFU/g (G1) | 106 CFU/g (G2) | 107 CFU/g (G3) | |
| Initial weight (g) | 0.56 ± 0.021 a | 0.54 ± 0.016 a | 0.54 ± 0.020 a | 0.56 ± 0.017 a |
| Finial weight (g) | 1.31 ± 0.004 a | 1.25 ± 0.025 a | 1.41 ± 0.095 b | 1.58 ± 0.125 b |
| Weight gain (g) | 0.75 ± 0.020 a | 0.71 ± 0.013 a | 0.87 ± 0.114 b | 1.02 ± 0.111 b |
| Feed efficiency | 0.33 ± 0.010 a | 0.33 ± 0.015 a | 0.39 ± 0.044 b | 0.41 ± 0.034 b |
| Specific growth rate | 2.41 ± 0.153 a | 2.34 ± 0.061 a | 2.81 ± 0.471 b | 3.26 ± 0.290 b |
| Survival rate (%) | 97.8 ± 1.92 a | 95.6 ± 3.85a | 96.7 ± 0.00 a | 97.8 ± 1.92 a |
| Activity (U/mg) | Control | G1 | G2 | G3 |
|---|---|---|---|---|
| Protease | 0.50 ± 0.007 a | 0.51 ± 0.009 a | 0.64 ± 0.013 b | 0.68 ± 0.005 b |
| Amylase | 0.15 ± 0.003 a | 0.15 ± 0.08 a b | 0.18 ± 0.004 b | 0.18 ± 0.009 b |
| Cellulase | 0.14 ± 0.010 a | 0.15 ± 0.005 a b | 0.16 ± 0.004 a b | 0.18 ± 0.025 b |
| Xylanase | 0.10 ± 0.001 a | 0.11 ± 0.013 a | 0.11 ± 0.007 a | 0.10 ± 0.001 a |
| Lipase | 0.03 ± 0.003 a | 0.05 ± 0.004 b | 0.05 ± 0.003 b | 0.06 ± 0.003 c |
| Species | Control | G1 | G2 | G3 |
|---|---|---|---|---|
| Potential probiotics | ||||
| Bacillus safensis | 0.001% | 0.082% | 0.074% | 0.091% |
| Bacillus licheniformis | 0.002% | 0.23% | 0.19% | 0.28% |
| Lactococcus lactis | 0.004% | 0.11% | 0.09% | 0.14% |
| Potential pathogens | ||||
| Aeromonas veronii | 1.23% | 0.12% | 0.06% | 0.07% |
| Aeromonas jandaei | 0.95% | 0.081% | 0.017% | 0.048% |
| Enterovibrio nigricans | 0.31% | 0.002% | 0.005% | 0.009% |
| Enterovibrio coralii | 2.04% | 0.01% | 0.09% | 0.007% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, P.-S.; Liu, C.-H.; Hu, S.-Y. Probiotic Bacillus safensis NPUST1 Administration Improves Growth Performance, Gut Microbiota, and Innate Immunity against Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Microorganisms 2021, 9, 2494. https://doi.org/10.3390/microorganisms9122494
Wu P-S, Liu C-H, Hu S-Y. Probiotic Bacillus safensis NPUST1 Administration Improves Growth Performance, Gut Microbiota, and Innate Immunity against Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Microorganisms. 2021; 9(12):2494. https://doi.org/10.3390/microorganisms9122494
Chicago/Turabian StyleWu, Pei-Shan, Chun-Hong Liu, and Shao-Yang Hu. 2021. "Probiotic Bacillus safensis NPUST1 Administration Improves Growth Performance, Gut Microbiota, and Innate Immunity against Streptococcus iniae in Nile tilapia (Oreochromis niloticus)" Microorganisms 9, no. 12: 2494. https://doi.org/10.3390/microorganisms9122494
APA StyleWu, P.-S., Liu, C.-H., & Hu, S.-Y. (2021). Probiotic Bacillus safensis NPUST1 Administration Improves Growth Performance, Gut Microbiota, and Innate Immunity against Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Microorganisms, 9(12), 2494. https://doi.org/10.3390/microorganisms9122494