
Immunomodulation Potential of Probiotics: A Novel Strategy for Improving Livestock Health, Immunity, and Productivity


 ,
, 
 and
and 
Abstract
1. Introduction
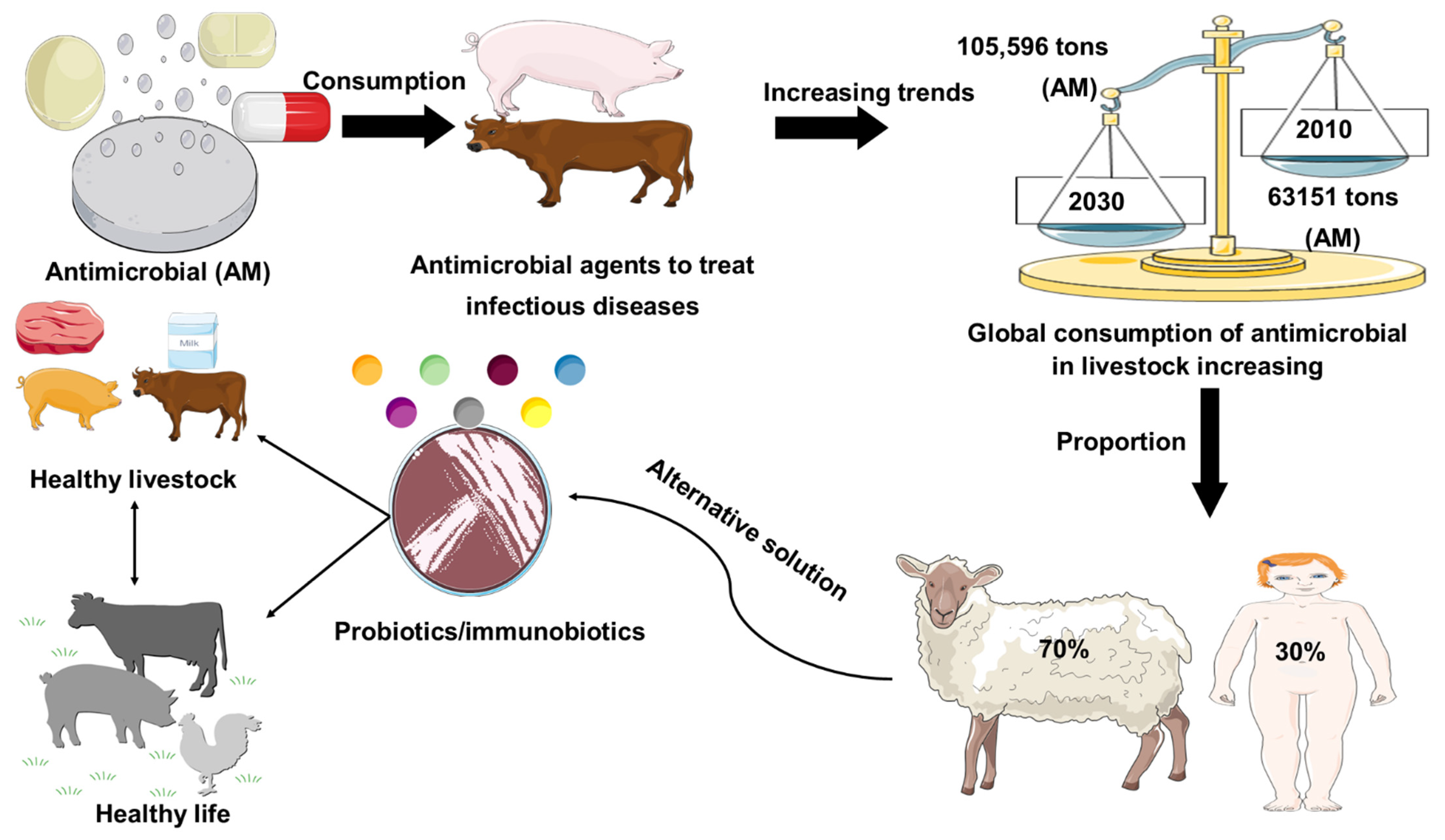
2. Global Trends of Antimicrobial Uses in Livestock
3. Application of Probiotics in In Vivo Studies for Pig Production
4. Application of Probiotics in In Vivo Studies for Ruminant Production
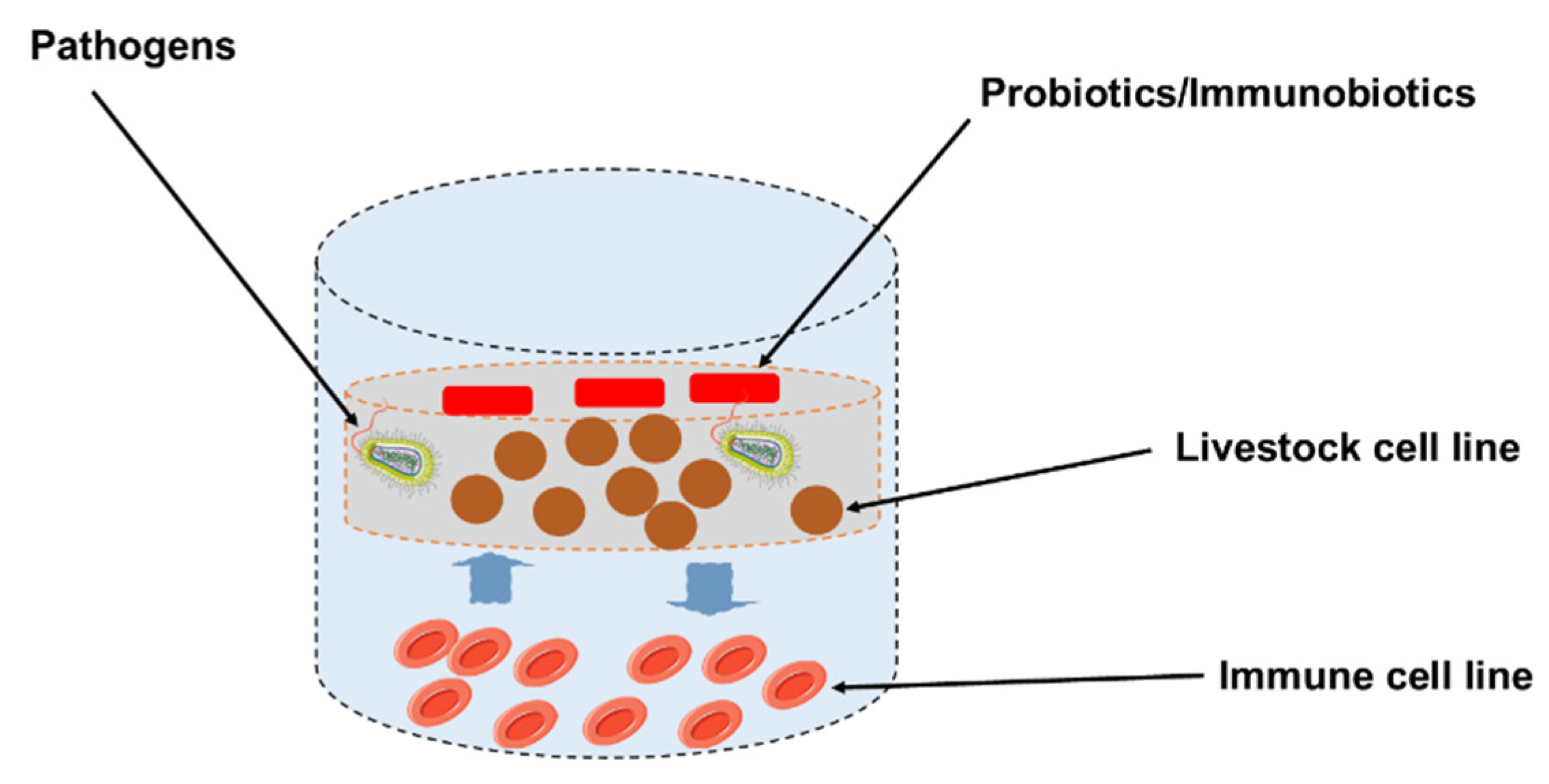
5. Application of Probiotics Using Cell Lines as Livestock Animal Model (In Vitro Study)
Limitation for the Use of Probiotics in In Vitro and In Vivo Research Model
6. Application of Probiotics in Livestock Production
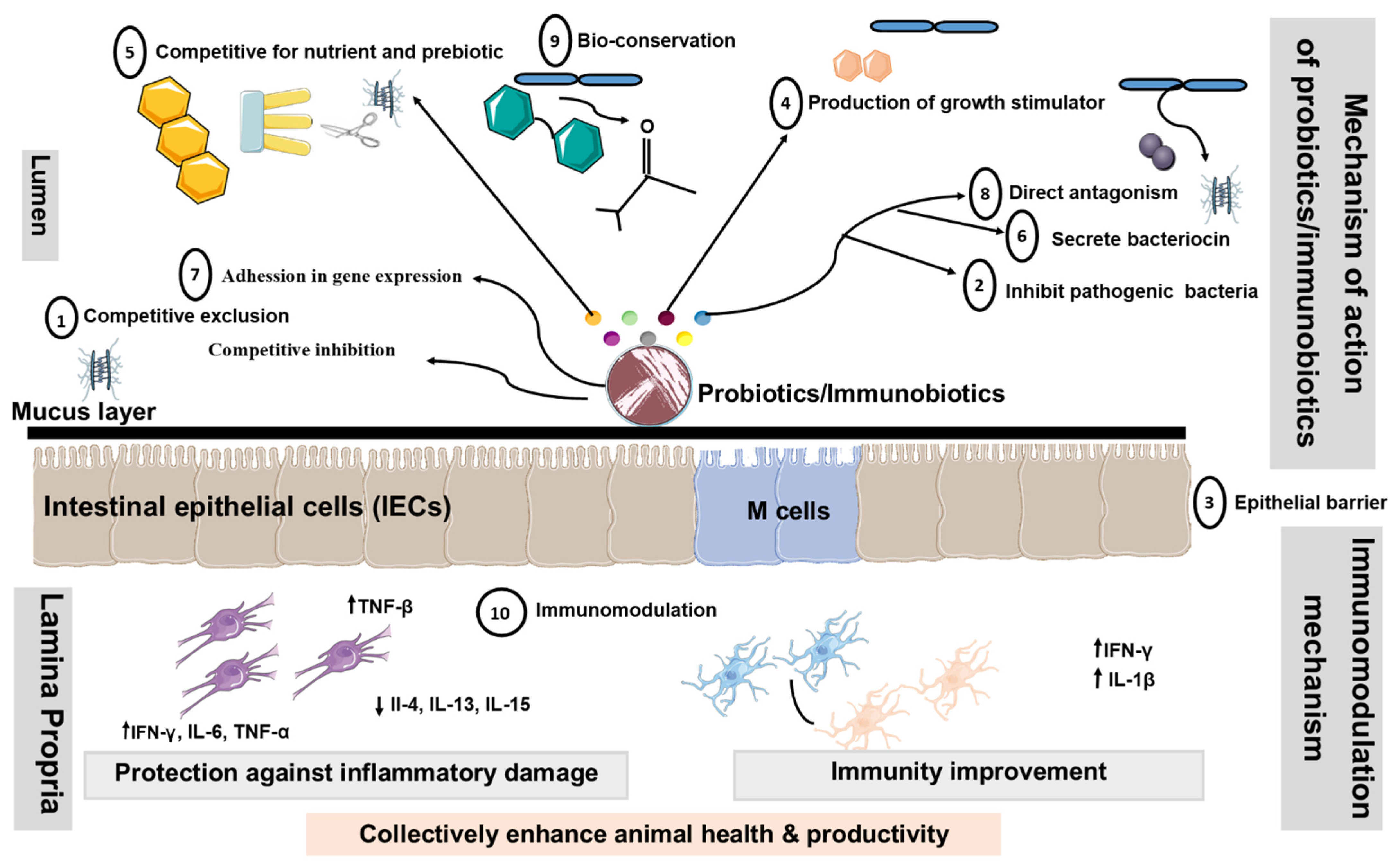
7. Modes of Action of Livestock Probiotics
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Komarek, A.M.; Dunston, S.; Enahoro, D.; Godfray, H.C.J.; Herrero, M.; Mason-D’Croz, D.; Rich, K.M.; Scarborough, P.; Springmann, M.; Sulser, T.B.; et al. Income, consumer preferences, and the future of livestock-derived food demand. Glob. Environ. Change 2021, 70, 102343. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.M.; El Zowalaty, M.E.; Lundkvist, Å.; Järhult, J.D.; Khan Nayem, M.R.; Tanzin, A.Z.; Badsha, M.R.; Khan, S.A.; Ashour, H.M. Residual antimicrobial agents in food originating from animals. Trends Food Sci. Technol. 2021, 111, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Schrijver, R.; Stijntjes, M.; Rodríguez-Baño, J.; Tacconelli, E.; Babu Rajendran, N.; Voss, A. Review of antimicrobial resistance surveillance programmes in livestock and meat in EU with focus on humans. Clin. Microbiol. Infect. 2018, 24, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Tiseo, K.; Huber, L.; Gilbert, M.; Robinson, T.P.; Van Boeckel, T.P. Global Trends in Antimicrobial Use in Food Animals from 2017 to 2030. Antibiotics 2020, 9, 918. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-González, P.F.; Peredo-Lovillo, A.; Castro-López, C.; Vallejo-Cordoba, B.; González-Córdova, A.F.; García, H.S.; Hernández-Mendoza, A. Food-grade lactic acid bacteria and probiotics as a potential protective tool against erythrotoxic dietary xenobiotics. Trends Food Sci. Technol. 2021, 116, 1041–1055. [Google Scholar] [CrossRef]
- Barba-Vidal, E.; Martín-Orúe, S.M.; Castillejos, L. Practical aspects of the use of probiotics in pig production: A review. Livest. Sci. 2019, 223, 84–96. [Google Scholar] [CrossRef]
- Gibson, M.K.; Crofts, T.S.; Dantas, G. Antibiotics and the developing infant gut microbiota and resistome. Curr. Opin. Microbiol. 2015, 27, 51–56. [Google Scholar] [CrossRef]
- Tavoukjian, V. Faecalmicrobiota transplantation for the decolonization of antibiotic-resistant bacteria in the gut: A systematic review and meta-analysis. J. Hosp. Infect. 2019, 102, 174–188. [Google Scholar] [CrossRef]
- Andremont, A.; Cervesi, J.; Bandinelli, P.-A.; Vitry, F.; de Gunzburg, J. Spare and repair the gut microbiota from antibiotic-induced dysbiosis: State-of-the-art. Drug Discov. Today 2021, 26, 2159–2163. [Google Scholar] [CrossRef]
- Tomosada, Y.; Villena, J.; Murata, K.; Chiba, E.; Shimazu, T.; Aso, H.; Iwabuchi, N.; Xiao, J.Z.; Saito, T.; Kitazawa, H. Immunoregulatory effect of bifidobacteria strains in porcine intestinal epithelial cells through modulation of ubiquitin-editing enzyme A20 expression. PLoS ONE 2013, 8, e59259. [Google Scholar] [CrossRef]
- Suda, Y.; Villena, J.; Takahashi, Y.; Hosoya, S.; Tomosada, Y.; Tsukida, K.; Shimazu, T.; Aso, H.; Tohno, M.; Ishida, M.; et al. Immunobiotic Lactobacillus jensenii as immune-health promoting factor to improve growth performance and productivity in post-weaning pigs. BMC Immunol. 2014, 15, 24. [Google Scholar] [CrossRef] [PubMed]
- Fujie, H.; Villena, J.; Tohno, M.; Morie, K.; Shimazu, T.; Aso, H.; Suda, Y.; Shimosato, T.; Iwabuchi, N.; Xiao, J.Z.; et al. Toll-like receptor-2-activating bifidobacteria strains differentially regulate inflammatory cytokines in the porcine intestinal epithelial cell culture system: Finding new anti-inflammatory immunobiotics. FEMS Immunol. Med. Microbiol. 2011, 63, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.; Salva, S.; Núñez, M.; Corzo, J.; Tolaba, R.; Faedda, J.; Font, G.; Alvarez, S. Probiotics for Everyone! The Novel Immunobiotic Lactobacillus rhamnosus CRL1505 and the Beginning of Social Probiotic Programs in Argentina. Int. J. Biotechnol. Wellness Ind. 2012, 189–198. [Google Scholar] [CrossRef]
- Kumagae, N.; Villena, J.; Tomosada, Y.; Kobayashi, H.; Kanmani, P.; Aso, H.; Sasaki, T.; Yoshida, M.; Tanabe, H.; Shibata, I.; et al. Evaluation of the Immunoregulatory Capacities of Feed Microbial Materials in Porcine Intestinal Immune and Epithelial Cells. Open J. Vet. Med. 2014, 4, 14. [Google Scholar] [CrossRef]
- Kobayashi, H.; Kanmani, P.; Ishizuka, T.; Miyazaki, A.; Soma, J.; Albarracin, L.; Suda, Y.; Nochi, T.; Aso, H.; Iwabuchi, N.; et al. Development of an in vitro immunobiotic evaluation system against rotavirus infection in bovine intestinal epitheliocytes. Benef. Microbes 2017, 8, 309–321. [Google Scholar] [CrossRef]
- Dowarah, R.; Verma, A.K.; Agarwal, N.; Patel, B.H.M.; Singh, P. Effect of swine based probiotic on performance, diarrhoea scores, intestinal microbiota and gut health of grower-finisher crossbred pigs. Livest. Sci. 2017, 195, 74–79. [Google Scholar] [CrossRef]
- Kwak, M.-J.; Tan, P.L.; Oh, J.K.; Chae, K.S.; Kim, J.; Kim, S.H.; Eun, J.-S.; Chee, S.W.; Kang, D.-K.; Kim, S.H.; et al. The effects of multispecies probiotic formulations on growth performance, hepatic metabolism, intestinal integrity and fecal microbiota in growing-finishing pigs. Anim. Feed Sci. Technol. 2021, 274, 114833. [Google Scholar] [CrossRef]
- Meng, Q.W.; Yan, L.; Ao, X.; Zhou, T.X.; Wang, J.P.; Lee, J.H.; Kim, I.H. Influence of probiotics in different energy and nutrient density diets on growth performance, nutrient digestibility, meat quality, and blood characteristics in growing-finishing pigs. J. Anim. Sci. 2010, 88, 3320–3326. [Google Scholar] [CrossRef]
- Veizaj-Delia, E.; Piu, T.; Lekaj, P.; Tafaj, M. Using combined probiotic to improve growth performance of weaned piglets on extensive farm conditions. Livest. Sci. 2010, 134, 249–251. [Google Scholar] [CrossRef]
- Giang, H.; Viet, T.; Ogle, B.; Lindberg, J. Effects of Supplementation of Probiotics on the Performance, Nutrient Digestibility and FaecalMicroflora in Growing-finishing Pigs. Asian-Australas. J. Anim. Sci. 2011, 24, 655–661. [Google Scholar] [CrossRef]
- Suo, C.; Yin, Y.; Wang, X.; Lou, X.; Song, D.; Wang, X.; Gu, Q. Effects of lactobacillus plantarum ZJ316 on pig growth and pork quality. BMC Vet. Res. 2012, 8, 89. [Google Scholar] [CrossRef] [PubMed]
- Herfel, T.M.; Jacobi, S.K.; Lin, X.; Jouni, Z.E.; Chichlowski, M.; Stahl, C.H.; Odle, J. Dietary supplementation of Bifidobacterium longum strain AH1206 increases its cecal abundance and elevates intestinal interleukin-10 expression in the neonatal piglet. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 60, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, J.; Zhang, S.; Yang, F.; Thacker, P.A.; Zhang, G.; Qiao, S.; Ma, X. Oral administration of Lactobacillus fermentum I5007 favors intestinal development and alters the intestinal microbiota in formula-fed piglets. J. Agric. Food Chem. 2014, 62, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Sonia, T.; Ji, H.; Hong-Seok, M.; Chul-Ju, Y. Evaluation of Lactobacillus and Bacillus-based probiotics as alternatives to antibiotics in enteric microbial challenged weaned piglets. Afr. J. Microbiol. Res. 2014, 8, 96–104. [Google Scholar] [CrossRef]
- Kantas, D.; Papatsiros, V.G.; Tassis, P.D.; Giavasis, I.; Bouki, P.; Tzika, E.D. A feed additive containing Bacillus toyonensis (Toyocerin®) protects against enteric pathogens in postweaning piglets. J. Appl. Microbiol. 2015, 118, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Lan, R.X.; Lee, S.I.; Kim, I.H. Effects of multistrain probiotics on growth performance, nutrient digestibility, blood profiles, faecal microbial shedding, faecal score and noxious gas emission in weaning pigs. J. Anim. Physiol. Anim. Nutr. 2016, 100, 1130–1138. [Google Scholar] [CrossRef]
- Barba-Vidal, E.; Castillejos, L.; López-Colom, P.; RiveroUrgell, M.; Moreno Muñoz, J.A.; Martín-Orúe, S.M. Evaluation of the Probiotic Strain Bifidobacterium longum subsp. Infantis CECT 7210 Capacities to Improve Health Status and Fight Digestive Pathogens in a Piglet Model. Front. Microbiol. 2017, 8, 533. [Google Scholar] [CrossRef]
- Ngo, T.T.; Bang, N.N.; Dart, P.; Callaghan, M.; Klieve, A.; Hayes, B.; McNeill, D. Feed Preference Response of Weaner Bull Calves to Bacillus amyloliquefaciens H57 Probiotic and Associated Volatile Organic Compounds in High Concentrate Feed Pellets. Animals 2021, 11, 51. [Google Scholar] [CrossRef]
- Masumizu, Y.; Zhou, B.; Kober, A.K.M.H.; Islam, M.A.; Iida, H.; Ikeda-Ohtsubo, W.; Suda, Y.; Albarracin, L.; Nochi, T.; Aso, H.; et al. Isolation and Immunocharacterization of Lactobacillus salivarius from the Intestine of Wakame-Fed Pigs to Develop Novel “Immunosynbiotics”. Microorganisms 2019, 7, 167. [Google Scholar] [CrossRef]
- Tan, B.; Lim, T.; Boontiam, W. Effect of dietary supplementation with essential oils and a Bacillus probiotic on growth performance, diarrhoea and blood metabolites in weaned pigs. Anim. Prod. Sci. 2020, 61, 64–71. [Google Scholar] [CrossRef]
- Islam, M.A.; Hashiguchi, K.; Kober, A.K.M.H.; Morie, K.; Zhou, B.; Tomokiyo, M.; Shimazu, T.; Aso, H.; Villena, J.; Suda, Y.; et al. Effect of Dietary Supplementation of ImmunobioticLactiplantibacillusplantarum N14 Fermented Rakkyo (Allium chinense) Pickled Juice on the Immunocompetence and Production Performance of Pigs. Animals 2021, 11, 752. [Google Scholar] [CrossRef] [PubMed]
- Suda, Y.; Sasaki, N.; Kagawa, K.; Elean, M.; Zhou, B.; Tomokiyo, M.; Islam, M.A.; Rajoka, M.S.R.; Kober, A.K.M.H.; Shimazu, T.; et al. Immunobiotic Feed Developed with Lactobacillus delbrueckii subsp. delbrueckii TUA4408L and the Soymilk By-Product Okara Improves Health and Growth Performance in Pigs. Microorganisms 2021, 9, 921. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Cui, Y.; Lu, H.; Wang, G.; Ma, X. Effect of long-term dietary probiotic Lactobacillus reuteri 1 or antibiotics on meat quality, muscular amino acids and fatty acids in pigs. Meat Sci. 2021, 171, 108234. [Google Scholar] [CrossRef]
- Wang, H.; Kim, I.-H. Evaluation of Dietary Probiotic (Lactobacillus plantarum BG0001) Supplementation on the Growth Performance, Nutrient Digestibility, Blood Profile, Fecal Gas Emission, and Fecal Microbiota in Weaning Pigs. Animals 2021, 11, 2232. [Google Scholar] [CrossRef] [PubMed]
- Frizzo, L.S.; Zbrun, M.V.; Soto, L.P.; Signorini, M.L. Effects of probiotics on growth performance in young calves: A meta-analysis of randomized controlled trials. Anim. Feed Sci. Technol. 2011, 169, 147–156. [Google Scholar] [CrossRef]
- Jatkauskas, J.; Vrotniakiene, V. Effects of encapsulated probiotic Enterococcus faecium strain on diarrhoea patterns and performance of early weaned calves. Vet. Zootech. 2014, 67, 47–52. [Google Scholar]
- Roos, T.B.; de Moraes, C.M.; Sturbelle, R.T.; Dummer, L.A.; Fischer, G.; Leite, F.P.L. Probiotics Bacillus toyonensis and Saccharomyces boulardii improve the vaccine immune response to Bovine herpesvirus type 5 in sheep. Res. Vet. Sci. 2018, 117, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.K.; Karim, S.A. Effect of individual and mixed live yeast culture feeding on growth performance, nutrient utilization and microbial crude protein synthesis in lambs. Anim. Feed Sci. Technol. 2010, 155, 163–171. [Google Scholar] [CrossRef]
- Bayatkouhsar, J.; Tahmasebi, A.M.; Naserian, A.A.; Mokarram, R.R.; Valizadeh, R. Effects of supplementation of lactic acid bacteria on growth performance, blood metabolites and fecal coliform and lactobacilli of young dairy calves. Anim. Feed Sci. Technol. 2013, 186, 1–11. [Google Scholar] [CrossRef]
- Soren, N.M.; Tripathi, M.K.; Bhatt, R.S.; Karim, S.A. Effect of yeast supplementation on the growth performance of Malpura lambs. Trop. Anim. Health Prod. 2013, 45, 547–554. [Google Scholar] [CrossRef]
- Agazzi, A.; Tirloni, E.; Stella, S.; Maroccolo, S.; Ripamonti, B.; Bersani, C.; Caputo, J.; Dell’Orto, V.; Rota, N.; Savoini, G. Effects of species-specific probiotic addition to milk replacer on calf health and performance during the first month of life. Ann. Anim. Sci. 2014, 14, 101–115. [Google Scholar] [CrossRef][Green Version]
- Foditsch, C.; Pereira, R.V.; Ganda, E.K.; Gomez, M.S.; Marques, E.C.; Santin, T.; Bicalho, R.C. Oral Administration of Faecalibacteriumprausnitzii Decreased the Incidence of Severe Diarrhea and Related Mortality Rate and Increased Weight Gain in Preweaned Dairy Heifers. PLoS ONE 2015, 10, e0145485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhou, M.; Tu, Y.; Zhang, N.F.; Deng, K.D.; Ma, T.; Diao, Q.Y. Effect of oral administration of probiotics on growth performance, apparent nutrient digestibility and stress-related indicators in Holstein calves. J. Anim. Physiol. Anim. Nutr. 2016, 100, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.; Zanouny, A.; Singer, A. Growth Performance, Nutrients Digestibility, and Blood Metabolites of Lambs Fed Diets Supplemented with Probiotics during Pre- and Post-Weaning Period. Asian-Australas. J. Anim. Sci. 2016, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Adjei-Fremah, S.; Ekwemalor, K.; Asiamah, E.K.; Ismail, H.; Ibrahim, S.; Worku, M. Effect of probiotic supplementation on growth and global gene expression in dairy cows. J. Appl. Anim. Res. 2018, 46, 257–263. [Google Scholar] [CrossRef]
- Sharma, A.N.; Kumar, S.; Tyagi, A.K. Effects of mannan-oligosaccharides and Lactobacillus acidophilus supplementation on growth performance, nutrient utilization and faecal characteristics in Murrah buffalo calves. J. Anim. Physiol. Anim. Nutr. 2018, 102, 679–689. [Google Scholar] [CrossRef]
- Maldonado, N.C.; Chiaraviglio, J.; Bru, E.; De Chazal, L.; Santos, V.; Nader-Macías, M.E.F. Effect of Milk Fermented with Lactic Acid Bacteria on Diarrheal Incidence, Growth Performance and Microbiological and Blood Profiles of Newborn Dairy Calves. Probiotics Antimicrob. Proteins 2018, 10, 668–676. [Google Scholar] [CrossRef]
- Hassan, A.; Gado, H.; Anele, U.Y.; Berasain, M.A.M.; Salem, A.Z.M. Influence of dietary probiotic inclusion on growth performance, nutrient utilization, ruminal fermentation activities and methane production in growing lambs. Anim. Biotechnol. 2020, 31, 365–372. [Google Scholar] [CrossRef]
- Zábranský, L. Effect of prebiotic and probiotic supplements to increase live weight of calves in the diet. Acta Fytotech. Zootech. 2021, 24, 164–167. [Google Scholar] [CrossRef]
- Liu, F.; Li, G.; Wen, K.; Bui, T.; Cao, D.; Zhang, Y.; Yuan, L. Porcine small intestinal epithelial cell line (IPEC-J2) of rotavirus infection as a new model for the study of innate immune responses to rotaviruses and probiotics. Viral Immunol. 2010, 23, 135–149. [Google Scholar] [CrossRef]
- Kanmani, P.; Kim, H. Immunobiotic strains modulate toll-like receptor 3 agonist induced innate antiviral immune response in human intestinal epithelial cells by modulating IFN regulatory factor 3 and NF-κB signaling. Front. Immunol. 2019, 10, 1536. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, N.; Tomosada, Y.; Villena, J.; Murata, K.; Takahashi, T.; Chiba, E.; Tohno, M.; Shimazu, T.; Aso, H.; Suda, Y.; et al. Advanced application of bovine intestinal epithelial cell line for evaluating regulatory effect of lactobacilli against heat-killed enterotoxigenic Escherichia coli-mediated inflammation. BMC Microbiol. 2013, 13, 54. [Google Scholar] [CrossRef] [PubMed]
- Abedi, D.; Feizizadeh, S.; Akbari, V.; Jafarian-Dehkordi, A. In vitro anti-bacterial and anti-adherence effects of Lactobacillus delbrueckii subsp bulgaricus on Escherichia coli. Res. Pharm. Sci. 2013, 8, 260–268. [Google Scholar] [PubMed]
- Shimazu, T.; Villena, J.; Tohno, M.; Fujie, H.; Hosoya, S.; Shimosato, T.; Aso, H.; Suda, Y.; Kawai, Y.; Saito, T.; et al. Immunobiotic Lactobacillus jensenii elicits anti-inflammatory activity in porcine intestinal epithelial cells by modulating negative regulators of the Toll-like receptor signaling pathway. Infect. Immun. 2012, 80, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.J.; Shin, H.S. Antimicrobial and immunomodulatory effects of bifidobacterium strains: A review. J. Microbiol. Biotechnol. 2021, 30, 1793–1800. [Google Scholar] [CrossRef]
- Villena, J.; Chiba, E.; Vizoso-Pinto, M.G.; Tomosada, Y.; Takahashi, T.; Ishizuka, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H. Immunobiotic Lactobacillus rhamnosus strains differentially modulate antiviral immune response in porcine intestinal epithelial and antigen presenting cells. BMC Microbiol. 2014, 14, 126. [Google Scholar] [CrossRef]
- Kang, J.Y.; Lee, D.K.; Ha, N.J.; Shin, H.S. Antiviral effects of Lactobacillus ruminis SPM0211 and Bifidobacterium longum SPM1205 and SPM1206 on rotavirus-infected Caco-2 cells and a neonatal mouse model. J. Microbiol. 2015, 53, 796–803. [Google Scholar] [CrossRef]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory Effects Triggered by Lactic Acid Bacteria Exopolysaccharides: New Insights into Molecular Interactions with Host Cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef]
- Kanmani, P.; Albarracin, L.; Kobayashi, H.; Iida, H.; Komatsu, R.; Humayun Kober, A.K.M.; Ikeda-Ohtsubo, W.; Suda, Y.; Aso, H.; Makino, S.; et al. Exopolysaccharides from Lactobacillus delbrueckii OLL1073R-1 modulate innate antiviral immune response in porcine intestinal epithelial cells. Mol. Immunol. 2018, 93, 253–265. [Google Scholar] [CrossRef]
- Iida, H.; Tohno, M.; Islam, M.A.; Sato, N.; Kobayashi, H.; Albarracin, L.; Kober, A.H.; Ikeda-Ohtsubo, W.; Suda, Y.; Aso, H.; et al. Paraimmunobiotic Bifidobacteria Modulate the Expression Patterns of Peptidoglycan Recognition Proteins in Porcine Intestinal Epitheliocytes and Antigen Presenting Cells. Cells 2019, 8, 891. [Google Scholar] [CrossRef]
- Mizuno, H.; Arce, L.; Tomotsune, K.; Albarracin, L.; Funabashi, R.; Vera, D.; Islam, M.A.; Vizoso-Pinto, M.G.; Takahashi, H.; Sasaki, Y.; et al. Lipoteichoic Acid Is Involved in the Ability of the Immunobiotic Strain Lactobacillus plantarum CRL1506 to Modulate the Intestinal Antiviral Innate Immunity Triggered by TLR3 Activation. Front. Immunol 2020, 11, 571. [Google Scholar] [CrossRef] [PubMed]
- Śliżewska, K.; Chlebicz-Wójcik, A.; Nowak, A. Probiotic Properties of New Lactobacillus Strains Intended to Be Used as Feed Additives for Monogastric Animals. Probiotics Antimicrob. Proteins 2021, 13, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- De Rycker, M.; Baragaña, B.; Duce, S.L.; Gilbert, I.H. Challenges and recent progress in drug discovery for tropical diseases. Nature 2018, 559, 498–506. [Google Scholar] [CrossRef]
- Álvarez-Ordóñez, A.; Carvajal, A.; Arguello, H.; Martínez-Lobo, F.J.; Naharro, G.; Rubio, P. Antibacterial activity and mode of action of a commercial citrus fruit extract. J. Appl. Microbiol. 2013, 115, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ha, B.; Kim, I.H. Effects of probiotics complex supplementation in low nutrient density diet on growth performance, nutrient digestibility, faecal microbial, and faecal noxious gas emission in growing pigs. Ital. J. Anim. Sci. 2021, 20, 163–170. [Google Scholar] [CrossRef]
- Méndez-Palacios, N.; Méndez-Mendoza, M.; Vázquez-Flores, F.; Castro-Colombres, J.G.; Ramírez-Bribiesca, J.E. Productive and economic parameters of pigs supplemented from weaning to finishing with prebiotic and probiotic feed additives. Anim. Sci. J. 2018, 89, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Zommiti, M.; Chikindas, M.L.; Ferchichi, M. Probiotics—Live Biotherapeutics: A Story of Success, Limitations, and Future Prospects—Not Only for Humans. Probiotics Antimicrob. Proteins 2020, 12, 1266–1289. [Google Scholar] [CrossRef]
- Tang, Q.; Yi, H.; Hong, W.; Wu, Q.; Yang, X.; Hu, S.; Xiong, Y.; Wang, L.; Jiang, Z. Comparative Effects of L. plantarum CGMCC 1258 and L. reuteri LR1 on Growth Performance, Antioxidant Function, and Intestinal Immunity in Weaned Pigs. Front. Vet. Sci. 2021, 8, 728849. [Google Scholar] [CrossRef]
- Raabis, S.; Li, W.; Cersosimo, L. Effects and immune responses of probiotic treatment in ruminants. Vet. Immunol. Immunopathol. 2019, 208, 58–66. [Google Scholar] [CrossRef]
- Flint, H.J. The rumen microbial ecosystem—some recent developments. Trends Microbiol. 1997, 5, 483–488. [Google Scholar] [CrossRef]
- Andersen, T.O.; Kunath, B.J.; Hagen, L.H.; Arntzen, M.Ø.; Pope, P.B. Rumen metaproteomics: Closer to linking rumen microbial function to animal productivity traits. Methods 2021, 186, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Cagle, C.M.; Fonseca, M.A.; Callaway, T.R.; Runyan, C.A.; Cravey, M.D.; Tedeschi, L.O. Evaluation of the effects of live yeast on rumen parameters and in situ digestibility of dry matter and neutral detergent fiber in beef cattle fed growing and finishing diets. Appl. Anim. Sci. 2020, 36, 36–47. [Google Scholar] [CrossRef]
- Sousa, D.O.; Oliveira, C.A.; Velasquez, A.V.; Souza, J.M.; Chevaux, E.; Mari, L.J.; Silva, L.F.P. Live yeast supplementation improves rumen fibre degradation in cattle grazing tropical pastures throughout the year. Anim. Feed Sci. Technol. 2018, 236, 149–158. [Google Scholar] [CrossRef]
- Rossow, H.; Riordan, T.; Riordan, A. Effects of addition of a live yeast product on dairy cattle performance. J. Appl. Anim. Res. 2017, 46, 1–5. [Google Scholar] [CrossRef]
- Maamouri, O.; Ben Salem, M. Effect of yeast culture feed supply on growth, ruminal pH, and digestibility of fattening calves. Food Sci. Nutr. 2021, 9, 2762–2767. [Google Scholar] [CrossRef]
- Frizzo, L.; Bertozzi, E.; Soto, L.P.; Zbrun, M.; Sequeira, G.; Santina, R.; Armesto, R.; Rosmini, M. The Effect of Supplementation with Three Lactic Acid Bacteria from Bovine Origin on Growth Performance and Health Status of Young Calves. J. Anim. Vet. Adv. 2008, 7, 400–408. [Google Scholar]
- Arowolo, M.A.; He, J. Use of probiotics and botanical extracts to improve ruminant production in the tropics: A review. Anim. Nutr. 2018, 4, 241–249. [Google Scholar] [CrossRef]
- Uyeno, Y.; Shigemori, S.; Shimosato, T. Effect of probiotics/prebiotics on cattle health and productivity. Microbes Environ. 2015, 30, 126–132. [Google Scholar] [CrossRef]
- Colombo, E.A.; Cooke, R.F.; Brandão, A.P.; Wiegand, J.B.; Schubach, K.M.; Sowers, C.A.; Duff, G.C.; Block, E.; Gouvêa, V.N. Performance, health, and physiological responses of newly received feedlot cattle supplemented with pre- and probiotic ingredients. Animal 2021, 15, 100214. [Google Scholar] [CrossRef]
- Batista, L.H.C.; Cidrini, I.A.; Prados, L.F.; Cruz, A.A.C.; Torrecilhas, J.A.; Siqueira, G.R.; Resende, F.D. A meta-analysis of yeast products for beef cattle under stress conditions: Performance, health and physiological parameters. Anim. Feed Sci. Technol. 2022, 283, 115182. [Google Scholar] [CrossRef]
- Wang, H.; Yu, Z.; Gao, Z.; Li, Q.; Qiu, X.; Wu, F.; Guan, T.; Cao, B.; Su, H. Effects of compound probiotics on growth performance, rumen fermentation, blood parameters, and health status of neonatal Holstein calves. J. Dairy Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Górka, P.; Budzińska, K.; Budziński, W.; Jankowiak, T.; Kehoe, S.; Kański, J. Effect of probiotic and nucleotide supplementation in milk replacer on growth performance and fecal bacteria in calves. Livest. Sci. 2021, 250, 104556. [Google Scholar] [CrossRef]
- Alawneh, J.I.; Barreto, M.O.; Moore, R.J.; Soust, M.; Al-harbi, H.; James, A.S.; Krishnan, D.; Olchowy, T.W.J. Systematic review of an intervention: The use of probiotics to improve health and productivity of calves. Prev. Vet. Med. 2020, 183, 105147. [Google Scholar] [CrossRef]
- Izadi, b.; MohebbiFani, M.; Hosseinzadeh, S.; Shekarforoush, S.S.; Rasooli, A.; Nazifi, S. Effect of Bacillus coagulans probiotic on milk production and important economic and health indicators of raw milk of Holstein cows. Iran. Vet. J. 2020, 16, 5–14. [Google Scholar] [CrossRef]
- Endo, K.; Mine, Y.; Shuto, T.; Taji, T.; Murayama, T.; Nikawa, H. Comprehensive analysis of transcriptional profiles in oral epithelial-like cells stimulated with oral probiotic Lactobacillus spp. Arch. Oral Biol. 2020, 118, 104832. [Google Scholar] [CrossRef]
- Vale, G.C.; Mayer, M.P.A. Effect of probiotic Lactobacillus rhamnosus by-products on gingival epithelial cells challenged with Porphyromonasgingivalis. Arch. Oral Biol. 2021, 128, 105174. [Google Scholar] [CrossRef]
- Moue, M.; Tohno, M.; Shimazu, T.; Kido, T.; Aso, H.; Saito, T.; Kitazawa, H. Toll-like receptor 4 and cytokine expression involved in functional immune response in an originally established porcine intestinal epitheliocyte cell line. Biochim. Biophys. Acta 2008, 1780, 134–144. [Google Scholar] [CrossRef]
- Miyazawa, K.; Hondo, T.; Kanaya, T.; Tanaka, S.; Takakura, I.; Itani, W.; Rose, M.T.; Kitazawa, H.; Yamaguchi, T.; Aso, H. Characterization of newly established bovine intestinal epithelial cell line. Histochem. Cell Biol. 2010, 133, 125–134. [Google Scholar] [CrossRef]
- Hosoya, S.; Villena, J.; Shimazu, T.; Tohno, M.; Fujie, H.; Chiba, E.; Shimosato, T.; Aso, H.; Suda, Y.; Kawai, Y.; et al. Immunobiotic lactic acid bacteria beneficially regulate immune response triggered by poly(I:C) in porcine intestinal epithelial cells. Vet. Res. 2011, 42, 111. [Google Scholar] [CrossRef]
- Wachi, S.; Kanmani, P.; Tomosada, Y.; Kobayashi, H.; Yuri, T.; Egusa, S.; Shimazu, T.; Suda, Y.; Aso, H.; Sugawara, M.; et al. Lactobacillus delbrueckii TUA4408L and its extracellular polysaccharides attenuate enterotoxigenic Escherichia coli-induced inflammatory response in porcine intestinal epitheliocytes via Toll-like receptor-2 and 4. Mol. Nutr. Food Res. 2014, 58, 2080–2093. [Google Scholar] [CrossRef] [PubMed]
- Albarracin, L.; Kobayashi, H.; Iida, H.; Sato, N.; Nochi, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Transcriptomic Analysis of the Innate Antiviral Immune Response in Porcine Intestinal Epithelial Cells: Influence of Immunobiotic Lactobacilli. Front. Immunol. 2017, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Duarte, M.E.; Kim, S.W. Dietary inclusion of multispecies probiotics to reduce the severity of post-weaning diarrhea caused by Escherichia coli F18(+) in pigs. Anim. Nutr./Zhongguoxu Mu Shouyixuehui 2021, 7, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Sobrino, O.J.; Alba, C.; Arroyo, R.; Pérez, I.; Sariego, L.; Delgado, S.; Fernández, L.; de María, J.; Fumanal, P.; Fumanal, A.; et al. Replacement of Metaphylactic Antimicrobial Therapy by Oral Administration of Ligilactobacillus salivarius MP100 in a Pig Farm. Front. Vet. Sci. 2021, 8, 666887. [Google Scholar] [CrossRef] [PubMed]
- Dou, S.; Gadonna-Widehem, P.; Rome, V.; Hamoudi, D.; Rhazi, L.; Lakhal, L.; Larcher, T.; Bahi-Jaber, N.; Pinon-Quintana, A.; Guyonvarch, A.; et al. Characterisation of Early-Life Fecal Microbiota in Susceptible and Healthy Pigs to Post-Weaning Diarrhoea. PLoS ONE 2017, 12, e0169851. [Google Scholar] [CrossRef]
- Yang, Q.; Huang, X.; Zhao, S.; Sun, W.; Yan, Z.; Wang, P.; Li, S.; Huang, W.; Zhang, S.; Liu, L.; et al. Structure and Function of the Fecal Microbiota in Diarrheic Neonatal Piglets. Front. Microbiol. 2017, 8, 502. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, W.; Rył, A.; Mizerski, A.; Walczakiewicz, K.; Sipak, O.; Laszczyńska, M. Immunomodulatory potential of gut microbiome-derived short-chain fatty acids (SCFAs). Acta Biochim. Pol. 2019, 66, 1–12. [Google Scholar] [CrossRef]
- Sun, J.; Du, L.; Li, X.; Zhong, H.; Ding, Y.; Liu, Z.; Ge, L. Identification of the core bacteria in rectums of diarrheic and non-diarrheic piglets. Sci. Rep. 2019, 9, 18675. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, Z.; Yu, L.; Wu, S.; Sun, L.; Wu, S.; Xu, Q.; Cai, S.; Qin, N.; Bao, W. Examination of the temporal and spatial dynamics of the gut microbiome in newborn piglets reveals distinct microbial communities in six intestinal segments. Sci. Rep. 2019, 9, 3453. [Google Scholar] [CrossRef]
- Gruninger, R.J.; Puniya, A.K.; Callaghan, T.M.; Edwards, J.E.; Youssef, N.; Dagar, S.S.; Fliegerova, K.; Griffith, G.W.; Forster, R.; Tsang, A.; et al. Anaerobic fungi (phylum Neocallimastigomycota): Advances in understanding their taxonomy, life cycle, ecology, role and biotechnological potential. FEMS Microbiol. Ecol. 2014, 90, 1–17. [Google Scholar] [CrossRef]
- Hess, M.; Paul, S.S.; Puniya, A.K.; van der Giezen, M.; Shaw, C.; Edwards, J.E.; Fliegerová, K. Anaerobic Fungi: Past, Present, and Future. Front. Microbiol. 2020, 11, 584893. [Google Scholar] [CrossRef] [PubMed]
- Hagen, L.H.; Brooke, C.G.; Shaw, C.A.; Norbeck, A.D.; Piao, H.; Arntzen, M.Ø.; Olson, H.M.; Copeland, A.; Isern, N.; Shukla, A.; et al. Proteome specialization of anaerobic fungi during ruminal degradation of recalcitrant plant fiber. ISME J. 2021, 15, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Sella, S.R.B.; Bueno, T.; de Oliveira, A.A.B.; Karp, S.G.; Soccol, C.R. Bacillus subtilis natto as a potential probiotic in animal nutrition. Crit. Rev. Biotechnol. 2021, 41, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Yu, J.; Hartanto, R.; Qi, D. Dietary Supplementation with Saccharomyces cerevisiae, Clostridium butyricum and Their Combination Ameliorate Rumen Fermentation and Growth Performance of Heat-Stressed Goats. Animals 2021, 11, 2116. [Google Scholar] [CrossRef]
- Angulo, M.; Reyes-Becerril, M.; Medina-Córdova, N.; Tovar-Ramírez, D.; Angulo, C. Probiotic and nutritional effects of Debaryomyces hansenii on animals. Appl. Microbiol. Biotechnol. 2020, 104, 7689–7699. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Thirumdas, R.; Mehwish, H.M.; Umair, M.; Khurshid, M.; Hayat, H.F.; Phimolsiripol, Y.; Pallarés, N.; Martí-Quijal, F.J.; Barba, F.J. Role of Food Antioxidants in Modulating Gut Microbial Communities: Novel Understandings in Intestinal Oxidative Stress Damage and Their Impact on Host Health. Antioxidants 2021, 10, 1563. [Google Scholar] [CrossRef]
- Saleem, M.; Malik, S.; Mehwish, H.M.; Ali, M.W.; Hussain, N.; Khurshid, M.; Rajoka, M.S.R.; Chen, Y. Isolation and functional characterization of exopolysaccharide produced by Lactobacillus plantarum S123 isolated from traditional Chinese cheese. Arch. Microbiol. 2021, 203, 3061–3070. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Mehwish, H.M.; Xiong, Y.; Song, X.; Hussain, N.; Zhu, Q.; He, Z. Gut microbiota targeted nanomedicine for cancer therapy: Challenges and future considerations. Trends Food Sci. Technol. 2021, 107, 240–251. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Wu, Y.; Mehwish, H.M.; Bansal, M.; Zhao, L. Lactobacillus exopolysaccharides: New perspectives on engineering strategies, physiochemical functions, and immunomodulatory effects on host health. Trends Food Sci. Technol. 2020, 103, 36–48. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Mehwish, H.M.; Zhang, H.; Ashraf, M.; Fang, H.; Zeng, X.; Wu, Y.; Khurshid, M.; Zhao, L.; He, Z. Antibacterial and antioxidant activity of exopolysaccharide mediated silver nanoparticle synthesized by Lactobacillus brevis isolated from Chinese koumiss. Colloids Surf. B Biointerfaces 2020, 186, 110734. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Mehwish, H.M.; Fang, H.; Padhiar, A.A.; Zeng, X.; Khurshid, M.; He, Z.; Zhao, L. Characterization and anti-tumor activity of exopolysaccharide produced by Lactobacillus kefiri isolated from Chinese kefir grains. J. Funct. Foods 2019, 63, 103588. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Zhao, H.; Mehwish, H.M.; Li, N.; Lu, Y.; Lian, Z.; Shao, D.; Jin, M.; Li, Q.; Zhao, L.; et al. Anti-tumor potential of cell free culture supernatant of Lactobacillus rhamnosus strains isolated from human breast milk. Food Res. Int. 2019, 123, 286–297. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Zhao, H.; Lu, Y.; Lian, Z.; Li, N.; Hussain, N.; Shao, D.; Jin, M.; Li, Q.; Shi, J. Anticancer potential against cervix cancer (HeLa) cell line of probiotic Lactobacillus casei and Lactobacillus paracasei strains isolated from human breast milk. Food Funct. 2018, 9, 2705–2715. [Google Scholar] [CrossRef] [PubMed]
- Rajoka, M.S.R.; Mehwish, H.M.; Hayat, H.F.; Hussain, N.; Sarwar, S.; Aslam, H.; Nadeem, A.; Shi, J. Characterization, the Antioxidant and Antimicrobial Activity of Exopolysaccharide Isolated from Poultry Origin Lactobacilli. Probiotics Antimicrob. Proteins 2019, 11, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Mountzouris, K.C.; Balaskas, C.; Xanthakos, I.; Tzivinikou, A.; Fegeros, K. Effects of a multi-species probiotic on biomarkers of competitive exclusion efficacy in broilers challenged with Salmonella enteritidis. Br. Poult. Sci. 2009, 50, 467–478. [Google Scholar] [CrossRef]
- Zhao, P.Y.; Kim, I.H. Effect of direct-fed microbial on growth performance, nutrient digestibility, fecal noxious gas emission, fecal microbial flora and diarrhea score in weanling pigs. Anim. Feed Sci. Technol. 2015, 200, 86–92. [Google Scholar] [CrossRef]
- Walker, W.A. Role of nutrients and bacterial colonization in the development of intestinal host defense. J. Pediatr. Gastroenterol. Nutr. 2000, 30 (Suppl. 2), S2–S7. [Google Scholar] [CrossRef]
- Dowarah, R.; Verma, A.K.; Agarwal, N. The use of Lactobacillus as an alternative of antibiotic growth promoters in pigs: A review. Anim. Nutr./Zhongguoxu Mu Shouyixuehui 2017, 3, 1–6. [Google Scholar] [CrossRef]
- Johnson-Henry, K.C.; Hagen, K.E.; Gordonpour, M.; Tompkins, T.A.; Sherman, P.M. Surface-layer protein extracts from Lactobacillus helveticus inhibit enterohaemorrhagic Escherichia coli O157:H7 adhesion to epithelial cells. Cell. Microbiol. 2007, 9, 356–367. [Google Scholar] [CrossRef]
- Lessard, M.; Dupuis, M.; Gagnon, N.; Nadeau, E.; Matte, J.; Goulet, J.; Fairbrother, J. Administration of Pediococcus Acidilactici or Saccharomyces CerevisiaeBoulardii modulates development of porcine mucosal immunity and reduces intestinal bacterial translocation after Escherichia Coli challenge. J. Anim. Sci. 2008, 87, 922–934. [Google Scholar] [CrossRef]
- Sato, N.; Garcia-Castillo, V.; Yuzawa, M.; Islam, M.A.; Albarracin, L.; Tomokiyo, M.; Ikeda-Ohtsubo, W.; Garcia-Cancino, A.; Takahashi, H.; Villena, J.; et al. Immunobiotic Lactobacillus jensenii TL2937 Alleviates Dextran Sodium Sulfate-Induced Colitis by Differentially Modulating the Transcriptomic Response of Intestinal Epithelial Cells. Front. Immunol. 2020, 11, 2174. [Google Scholar] [CrossRef] [PubMed]
- Gangadharan, D.; Sivaramakrishnan, S.; Nampoothiri, K.M.; Sukumaran, R.K.; Pandey, A. Response surface methodology for the optimization of alpha amylase production by Bacillus amyloliquefaciens. Bioresour. Technol. 2008, 99, 4597–4602. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Kim, B.K.; Lee, B.H.; Jo, K.I.; Lee, N.K.; Chung, C.H.; Lee, Y.C.; Lee, J.W. Purification and characterization of cellulase produced by Bacillus amyoliquefaciens DL-3 utilizing rice hull. Bioresour. Technol. 2008, 99, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Fayol-Messaoudi, D.; Berger, C.N.; Coconnier-Polter, M.H.; Liévin-Le Moal, V.; Servin, A.L. pH-, Lactic acid-, and non-lactic acid-dependent activities of probiotic Lactobacilli against Salmonella enterica Serovar Typhimurium. Appl. Environ. Microbiol. 2005, 71, 6008–6013. [Google Scholar] [CrossRef] [PubMed]
- Daşkıran, M.; Önol, A.G.; Cengiz, Ö.; Ünsal, H.; Türkyılmaz, S.; Tatlı, O.; Sevim, Ö. Influence of dietary probiotic inclusion on growth performance, blood parameters, and intestinal microflora of male broiler chickens exposed to posthatch holding time. J. Appl. Poult. Res. 2012, 21, 612–622. [Google Scholar] [CrossRef]
- Higgins, D.A.; Pomianek, M.E.; Kraml, C.M.; Taylor, R.K.; Semmelhack, M.F.; Bassler, B.L. The major Vibrio cholerae autoinducer and its role in virulence factor production. Nature 2007, 450, 883–886. [Google Scholar] [CrossRef] [PubMed]
- Pringsulaka, O.; Rueangyotchanthana, K.; Suwannasai, N.; Watanapokasin, R.; Amnueysit, P.; Sunthornthummas, S.; Sukkhum, S.; Sarawaneeyaruk, S.; Rangsiruji, A. In vitro screening of lactic acid bacteria for multi-strain probiotics. Livest. Sci. 2015, 174, 66–73. [Google Scholar] [CrossRef]
- Oh, B.-T.; Jeong, S.-Y.; Velmurugan, P.; Park, J.-H.; Jeong, D.-Y. Probiotic-mediated blueberry (Vaccinium corymbosum L.) fruit fermentation to yield functionalized products for augmented antibacterial and antioxidant activity. J. Biosci. Bioeng. 2017, 124, 542–550. [Google Scholar] [CrossRef]
- Jana, U.K.; Suryawanshi, R.K.; Prajapati, B.P.; Kango, N. Prebiotic mannooligosaccharides: Synthesis, characterization and bioactive properties. Food Chem. 2021, 342, 128328. [Google Scholar] [CrossRef]
- García, C.; Rendueles, M.; Díaz, M. Liquid-phase food fermentations with microbial consortia involving lactic acid bacteria: A review. Food Res. Int. 2019, 119, 207–220. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genus | Species/Strains | Age Group | Probiotic Effects in Respect to | Ref. | ||||
|---|---|---|---|---|---|---|---|---|
| Weight Gain/Feed Intake | Feed Efficiency | Health | Immunity | Others | ||||
| Bacillus/Clostridium | B. subtilis and C. butyricum | Growing-finishing pigs (GFP) | ↑ | ↑ | ↑ | - | ↑ Meat qua | [18] |
| Lactobacillus/Enterococcus | L. plantarum ATCC 4336, L. fermentum DSM 20016 and E. faecium ATCC 19434 | Weaned piglets (WP) | ↑ | ↑ | - | - | - | [19] |
| Bacillus/Saccharomyces | B. subtilis and S. boulardi | GFP | ↑ | - | ↓ E. coli | [20] | ||
| Lactobacillus | L.plantarum ZJ316 | GFP | ↑ | - | ↑ | ↑ | ↑ Meat qua | [21] |
| Bifidobacterium | B. longum (AH1206) | Neonatal piglets | ↔ | ↔ | ↑ | ↑ | ↑ Mucosal immune | [22] |
| Lactobacillus | L. fermentum | Suckling/nursery piglets | ↑ | ↑ | ↑ | ↑ | ↓ Diarrhea | [23] |
| Lactobacillus | L. reuteri, B. subtillis and B. licheniformis | WP | ↑ | - | ↑ | ↑ | ↓ Salmonella and E. coli | [24] |
| Lactobacillus | L.jensenii TL2937 (LjTL2937) | GFP | ↑ | ↑ | ↑ | ↑ | ↑ Meat qua | [11] |
| Bacillus | B. toyonensis | WP | ↑ | ↑ | ↑ | - | ↓ Enteric pathogens | [25] |
| Bacillus/Clostridium | B. coagulans, B.licheniformis, B. subtilis and C. butyricum | WP | ↑ | ↓ | ↑ | ↑ | ↓ Fecal noxious gas emission | [26] |
| Bifidobacterium | B. longum | WP | ↑ | ↑ | ↑ | ↑ | ↓ Intestinal colonization by pathogens | [27] |
| Lactobacillus/Bifidobacterium | L. spp., B. spp. | GFP | ↑ | ↑ | ↑ | - | ↓ Post-weaning mortality | [28] |
| Lactobacillus | L. salivarius FFIG71 L. salivarius FFIG131 | GFP | - | - | ↑ | ↑ | - | [29] |
| Bacillus | B. subtilis PB6 | WP | ↑ | ↔ | ↑ | ↑ | ↓ Fecal ammonia and diarrhea | [30] |
| Lactiplantibacillus | L. plantarum N14 | GFP | ↑ | ↑ | - | ↑ | ↑ Meat qua | [31] |
| Lactobacillus | delbrueckii subsp. Delbrueckii TUA4408L | WP | ↑ | ↑ | ↑ | ↑ | ↑ Meat qua | [32] |
| Lactobacillus | L. reuteri 1 (LR1) | GFP | ↑ | ↑ | - | - | ↑ Meat qua | [33] |
| Lactobacillus | L. plantarum BG0001 | WP | ↑ | ↑ | - | - | ↓ Fecal E. coli | [34] |
| Genus | Species/Strains | Source | Probiotic Effects in Respect to | Ref. | ||||
|---|---|---|---|---|---|---|---|---|
| Weight Gain/Feed Intake | Feed Efficiency | Health | Immunity | Others | ||||
| - | Multi-species probiotic | Cattle | ↑ | ↔ | ↑ | - | [35] | |
| Enterococcus | E. faecium M74 | Calf | ↑ | - | ↑ | ↑ | - | [36] |
| Bacillus/Saccharomyces | B. cereus S. boulardii | Sheep | ↑ | [37] | ||||
| Kluyveromyces/Saccharomyces | K. marximanus NRRL3234, S. cerevisiae NCDC42, S. uvarum ATCC9080 | Sheep | ↑ | ↑ | - | - | - | [38] |
| - | Multi-species probiotic | Calf | ↔ | ↔ | ↔ | - | - | [35] |
| - | Multi-species probiotic | Cattle | ↑ | ↔ | - | - | - | [39] |
| Saccharomyces | S. cerevisiae or combination of S. cerevisiae and L. sporogenes | Sheep | ↔ | - | - | - | - | [40] |
| - | Multi-species probiotic LAB | Calves | ↑ | ↑ | - | ↓ Weaning time | [39] | |
| Lactobacillus | L.animalis SB310, L.paracasei subsp. Paracasei SB137 and B.coagulans SB117 | Calves | ↑ | ↑ | ↑ | - | Improved gut microbiota | [41] |
| Faecalibacterium | F. prausnitzii | Calves | ↑ | ↑ | - | ↑ | - | [42] |
| Lactobacillus | L. plantarum | Calves | ↑ | ↑ | ↑ | ↑ | ↓ Weaning stress | [43] |
| Pediococcus | P. pentosaceus | Sheep | ↑ | ↑ | ↑ | - | ↑ Digestibility | [44] |
| Lactobacillus | L. acidophilus, S. cerevisiae, E. faecium, A. oryza, and B. subtilis | Cattle | - | - | ↑ | ↑ | - | [45] |
| Lactobacillus | L. acidophilus | Buffalo calves | ↑ | ↑ | - | - | ↓ Fecal coliform | [46] |
| Lactobacillus | L. murinus CRL1695, L. mucosae CRL1696, L. johnsonii CRL1693, L. salivarius CRL1702 | Calves | ↑ | ↑ | - | - | ↓ Diarrhea and calf mortality | [47] |
| Ruminococcus | R. flavefaciens | Sheep | ↑ | ↑ | - | - | - | [48] |
| Lactobacillus | L. sporogenes | Calves | ↑ | ↑ | - | - | - | [49] |
| Genus | Species/Strains | Source | Expt. Mode | Time | Probiotic Effects in Respect to | Ref. | |||
|---|---|---|---|---|---|---|---|---|---|
| Anti-Inflammation | Pro-Inflammation | Immune-Health | Others | ||||||
| Lactobacillus | L. acidophilus (LA) or L. rhamnosus GG (LGG) | Dairy products | IPEC-J2 | 24 h | ↓ | ↑ | ↑ | ↓ RVs titers | [50] |
| Lactobacillus | L. casei MEP221106 | Korean food | PIE | 48 h | ↓ | ↑ | ↑ | ↑ AV immune | [51] |
| Bifidobacterium | B. breve MCC-117 | Human gut | PIE | 48 h | ↓ | ↑ | ↑ | ↓ Intestinal inflammation | [12] |
| Lactobacillus | L. casei OLL2768 | Dairy products | BIE | 48 h | ↓ | ↑ | ↑ | ↑ APP | [52] |
| Lactobacillus | L. delbrueckii | Yogurt | Caco-2 | Overnight | - | - | ↑ | ↓ E. coli infection | [53] |
| Bifidobacterium | B. longum BB536 and B. breve M-16V | Infant | PIE | 48 h | ↑ | ↓ | ↑ | ↑ APP | [10] |
| Lactobacillus | L. jensenii TL2937 | Human feces | PIE | 48 h | ↑ | ↓ | ↑ | ↑ APP | [11,54] |
| Bifidobacterium | B. thermophilum and B. infantis/B. brevis | Feed | PIE | 48 h | ↓ | ↑ | ↑ | - | [14,55] |
| Lactobacillus | L. rhamnosus CRL1506 | Goat milk | PIE | 48 h | ↓ | ↑ | ↑ | ↑ AV | [56] |
| Lactobacillus/Bifidobacterium | L. ruminis SPM0211, B. longum SPM1205 and SPM1206 | Human | Caco-2 | ↓ | ↑ | ↑ | ↑ AV immune | [57] | |
| Lactobacillus | L. salivarius FFIG53 | Pig intestine | PIE | 48 h | ↑ | ↓ | ↑ | ↑ APP | [29] |
| Lactobacillus | L. plantarum N14 and L. delbrueckii TUA4408L | Fermented pickle | PIE | 48 h | ↑ | ↓ | ↑ | - | [58] |
| Bifidobacterium | B.infantis MCC12 or B. breve MCC1274 | - | BIE | 48 h | ↓ | ↑ | ↑ | ↓ RVs titers | [15] |
| Lactobacillus | L. delbrueckii OLL1073R-1 | Yogurt | PIE | ↑ | ↓ | ↑ | ↑ AV | [59] | |
| Bifidobacterium | B. longum BB536 and B. breve M-16V | Human feces | PIE | 120 h | ↑ | ↓ | ↑ | - | [60] |
| Lactobacillus | L. plantarum CRL1506 | Goat milk | PIE | 72 h | ↑ | ↓ | ↑ | - | [61] |
| Lactobacillus | Lactobacillus spp. | Manure | Caco-2 cells | 7–10 D | - | - | - | ↓ Infections | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kober, A.K.M.H.; Riaz Rajoka, M.S.; Mehwish, H.M.; Villena, J.; Kitazawa, H. Immunomodulation Potential of Probiotics: A Novel Strategy for Improving Livestock Health, Immunity, and Productivity. Microorganisms 2022, 10, 388. https://doi.org/10.3390/microorganisms10020388
Kober AKMH, Riaz Rajoka MS, Mehwish HM, Villena J, Kitazawa H. Immunomodulation Potential of Probiotics: A Novel Strategy for Improving Livestock Health, Immunity, and Productivity. Microorganisms. 2022; 10(2):388. https://doi.org/10.3390/microorganisms10020388
Chicago/Turabian StyleKober, A. K. M. Humayun, Muhammad Shahid Riaz Rajoka, Hafiza Mahreen Mehwish, Julio Villena, and Haruki Kitazawa. 2022. "Immunomodulation Potential of Probiotics: A Novel Strategy for Improving Livestock Health, Immunity, and Productivity" Microorganisms 10, no. 2: 388. https://doi.org/10.3390/microorganisms10020388
APA StyleKober, A. K. M. H., Riaz Rajoka, M. S., Mehwish, H. M., Villena, J., & Kitazawa, H. (2022). Immunomodulation Potential of Probiotics: A Novel Strategy for Improving Livestock Health, Immunity, and Productivity. Microorganisms, 10(2), 388. https://doi.org/10.3390/microorganisms10020388