
Functional Metabolic Diversity of Bacterioplankton in Maritime Antarctic Lakes

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. FAPROTAX Analisys, Functional Prediction, and Metabolic Indices
2.3. Community Level Physiological Profiling
- ni: average qualitative color value of the three wells,
- c: average qualitative color value of control wells.
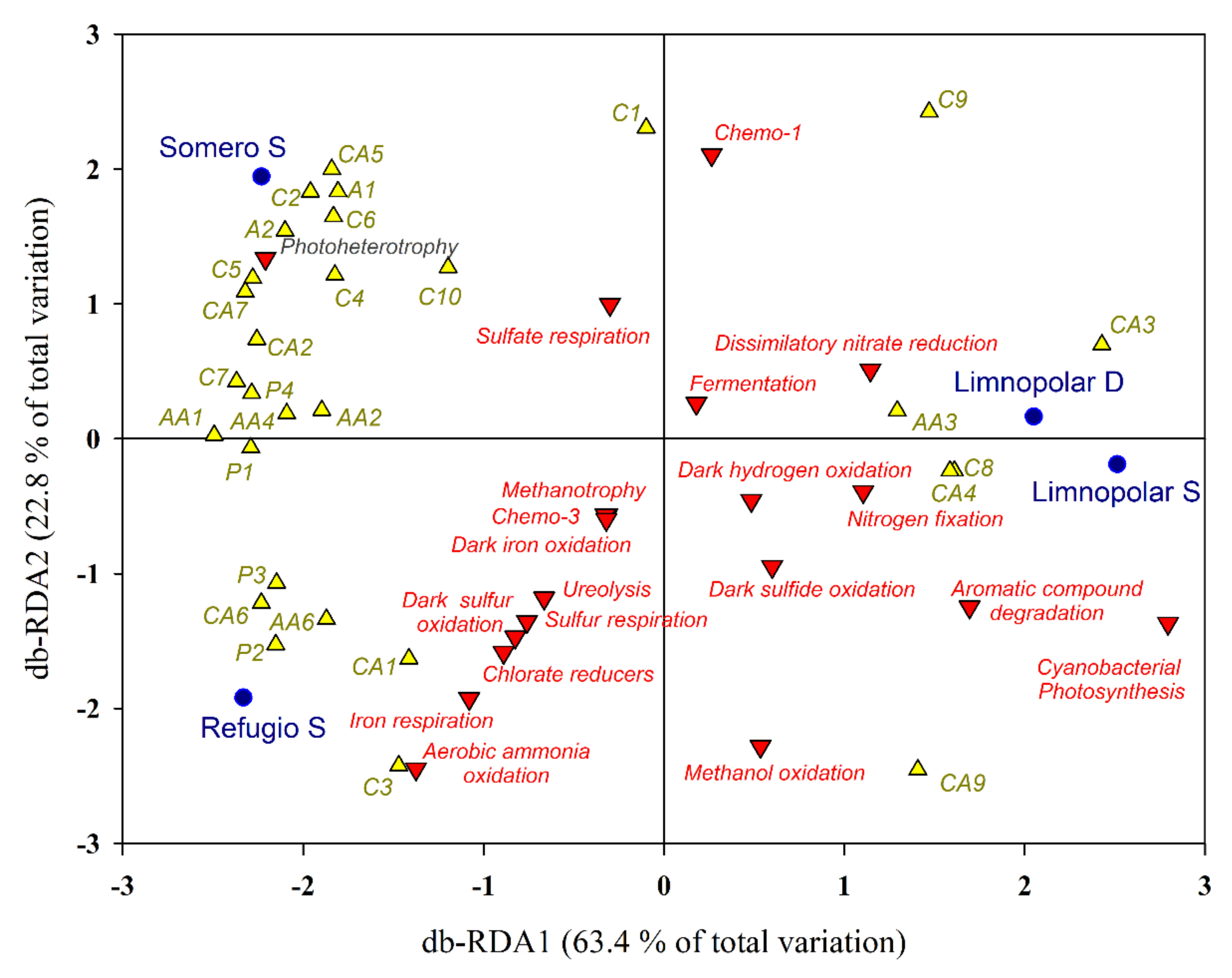
2.4. Clustering and Multivariate Ordination Analyses
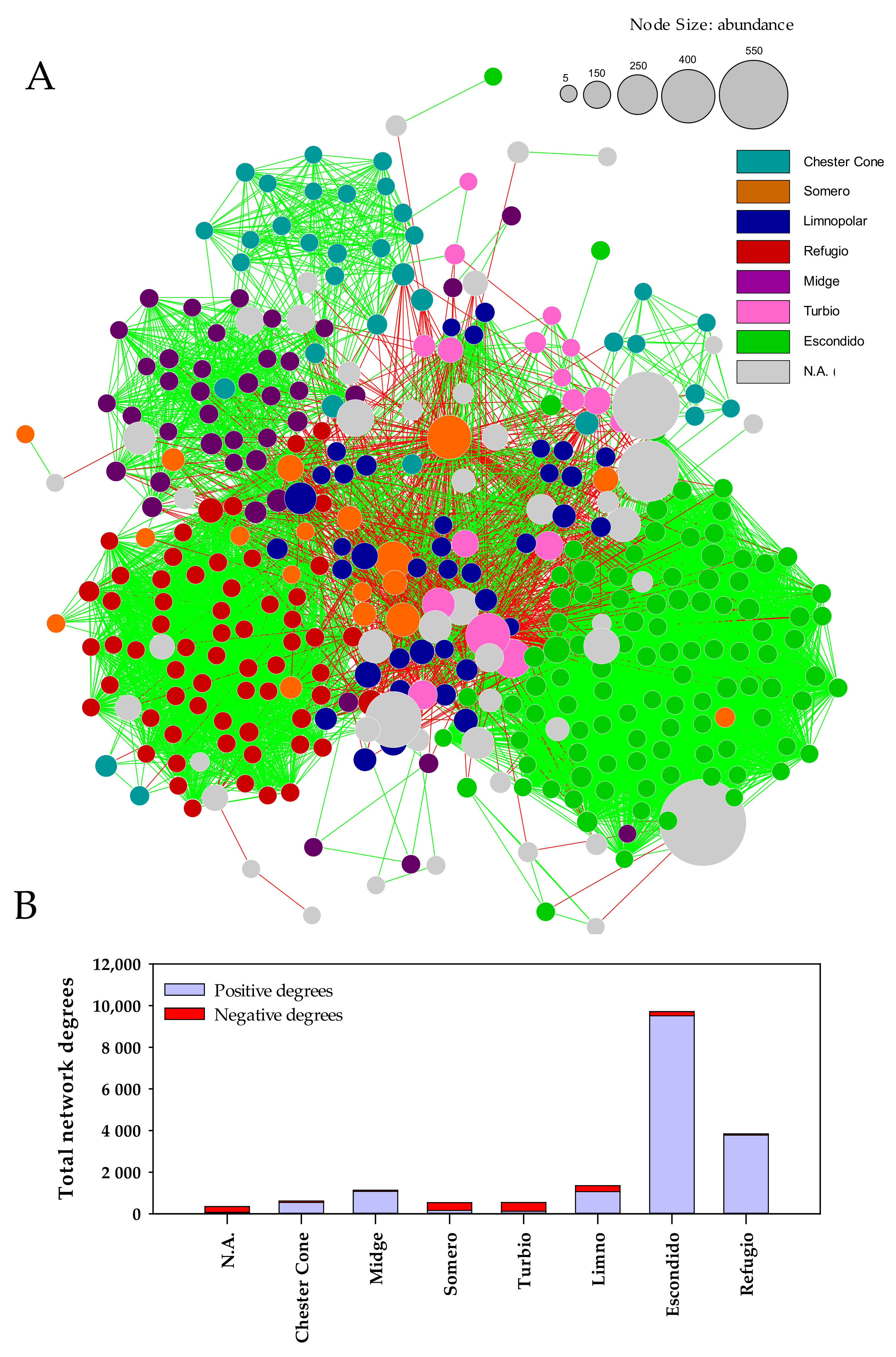
2.5. Co-Occurrence Network Analysis
3. Results
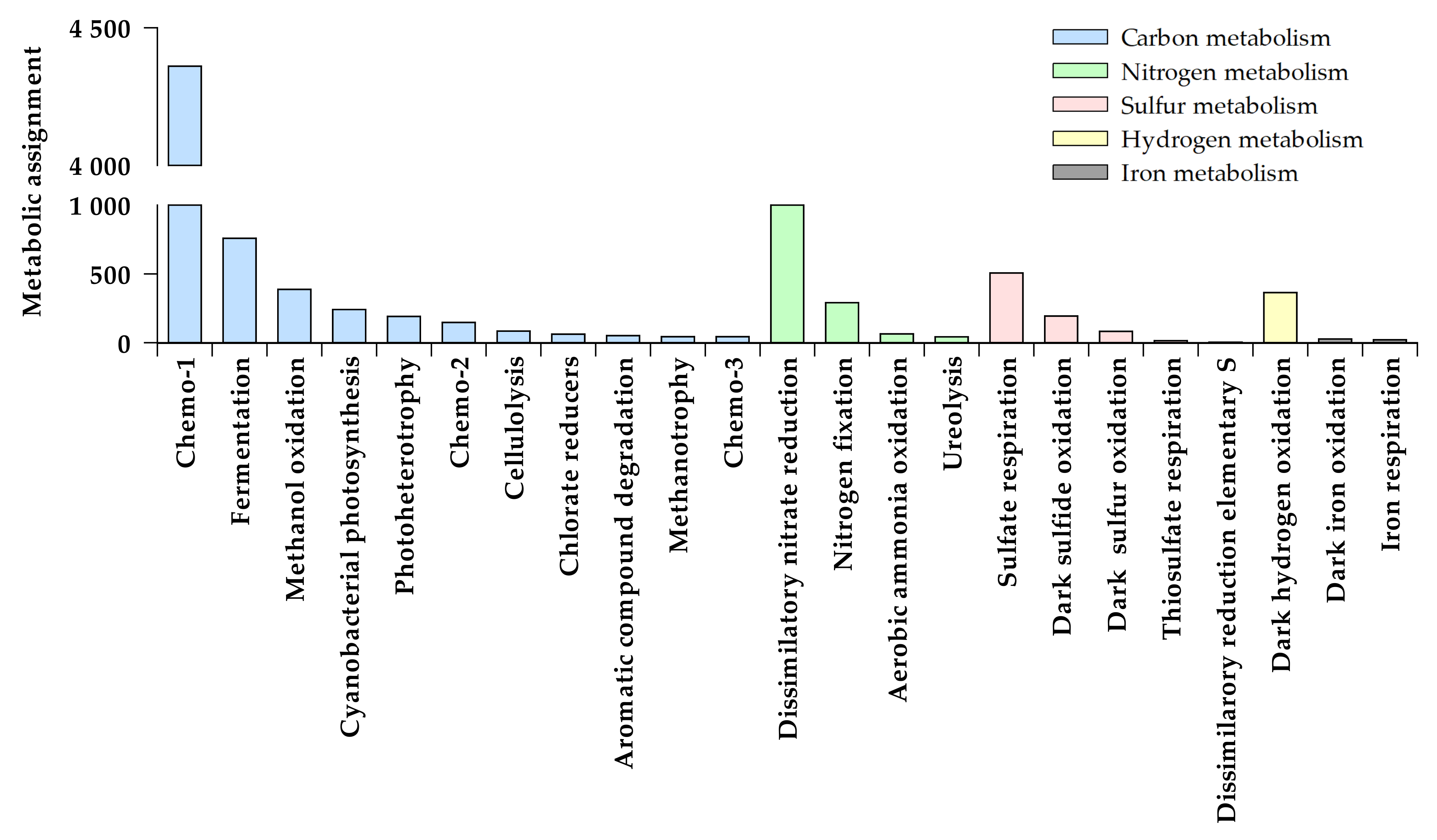
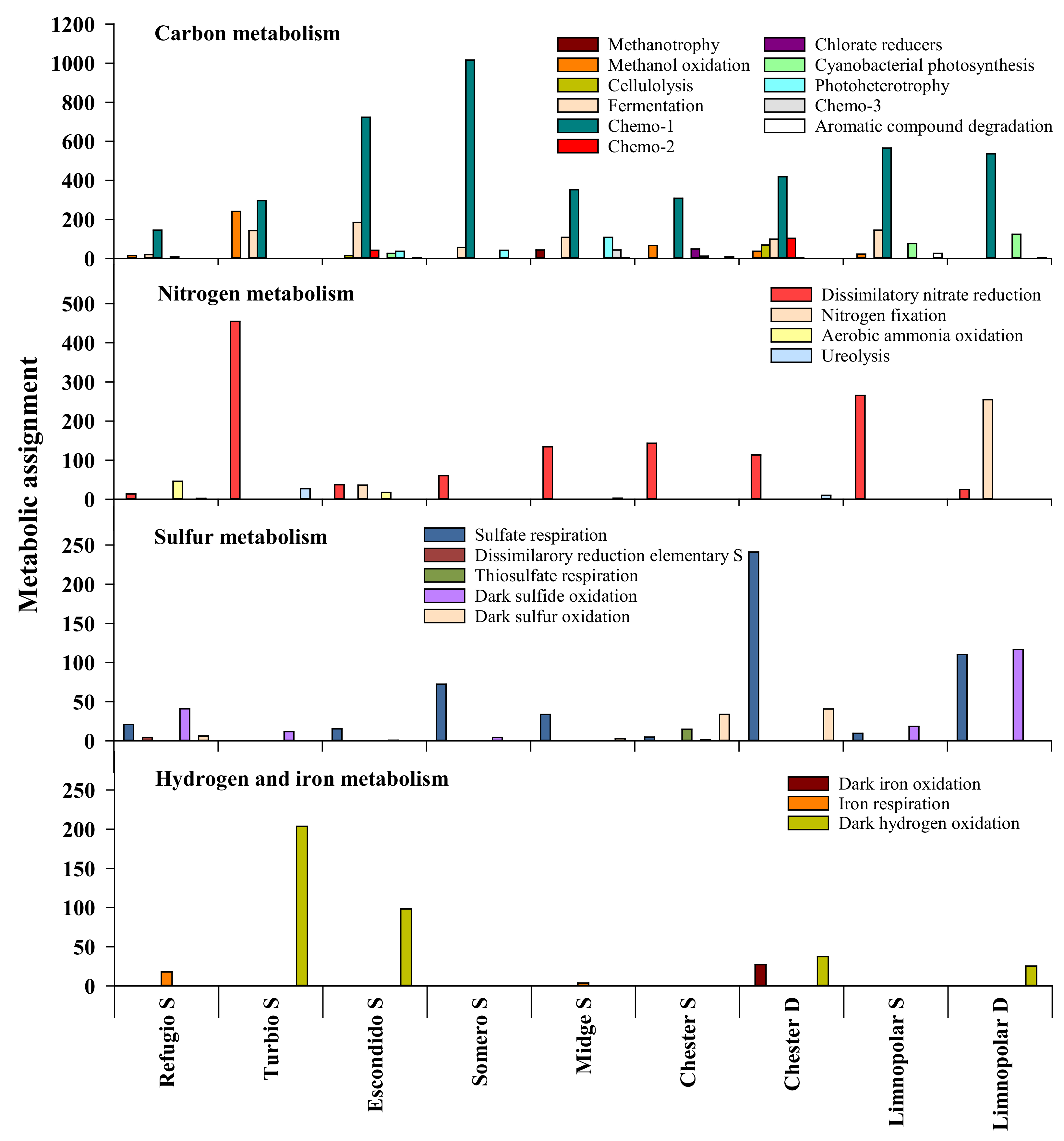
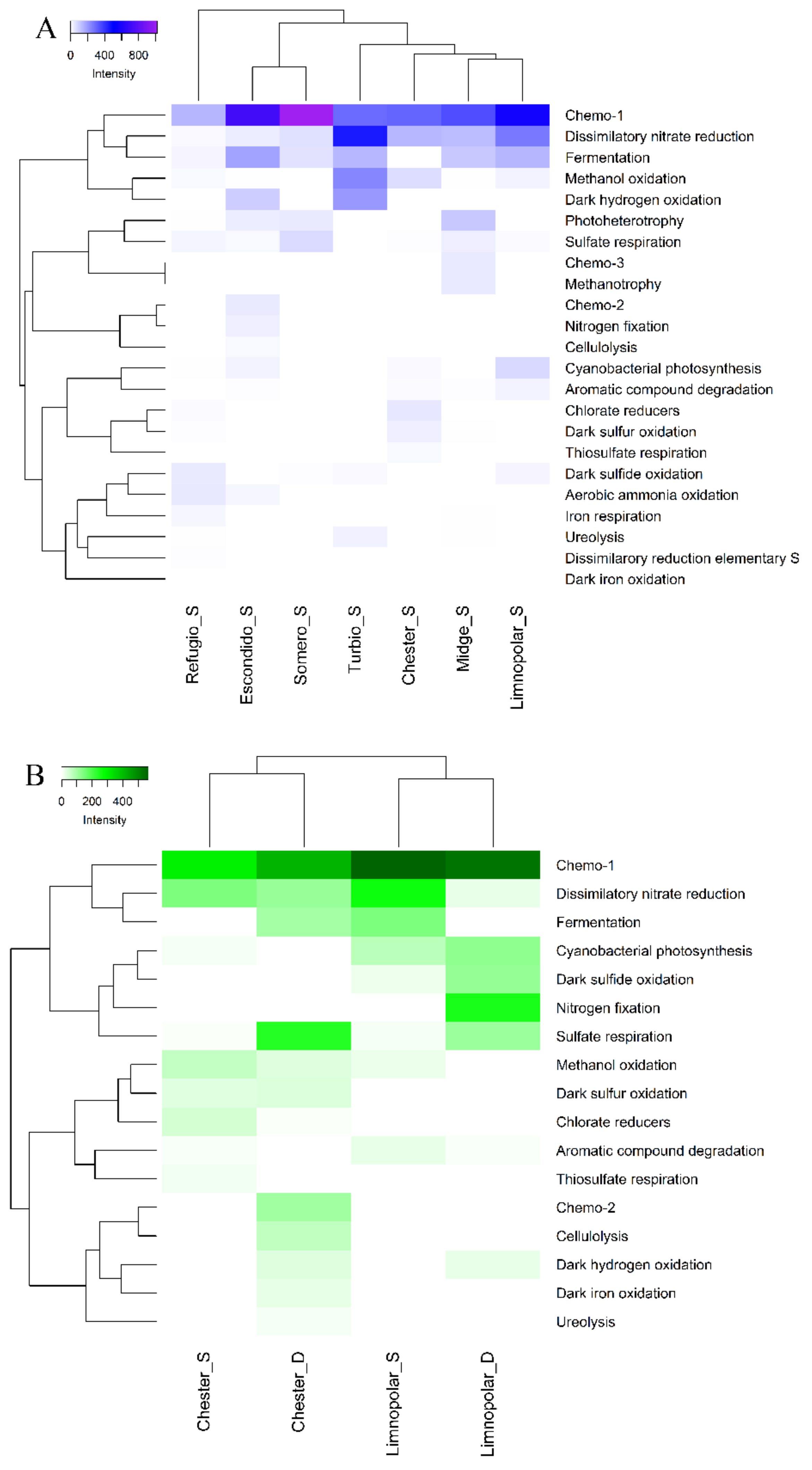
3.1. Functional Structure of Bacterioplankton Communities
3.1.1. Heterotrophic Metabolisms
3.1.2. Autotrophic Metabolisms
3.1.3. Other Respiratory Metabolisms
3.1.4. Assimilation of Nitrogen Compounds
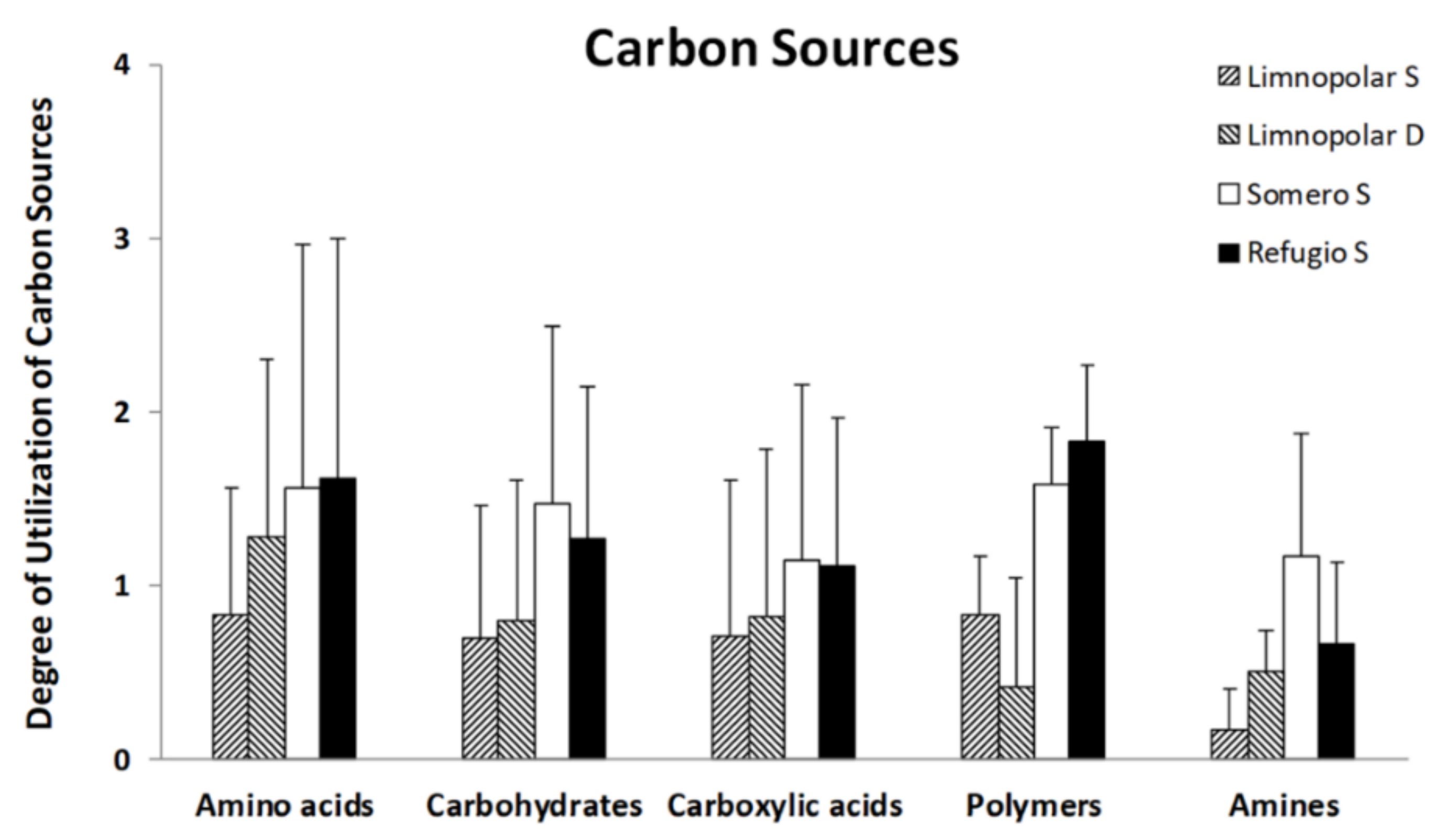
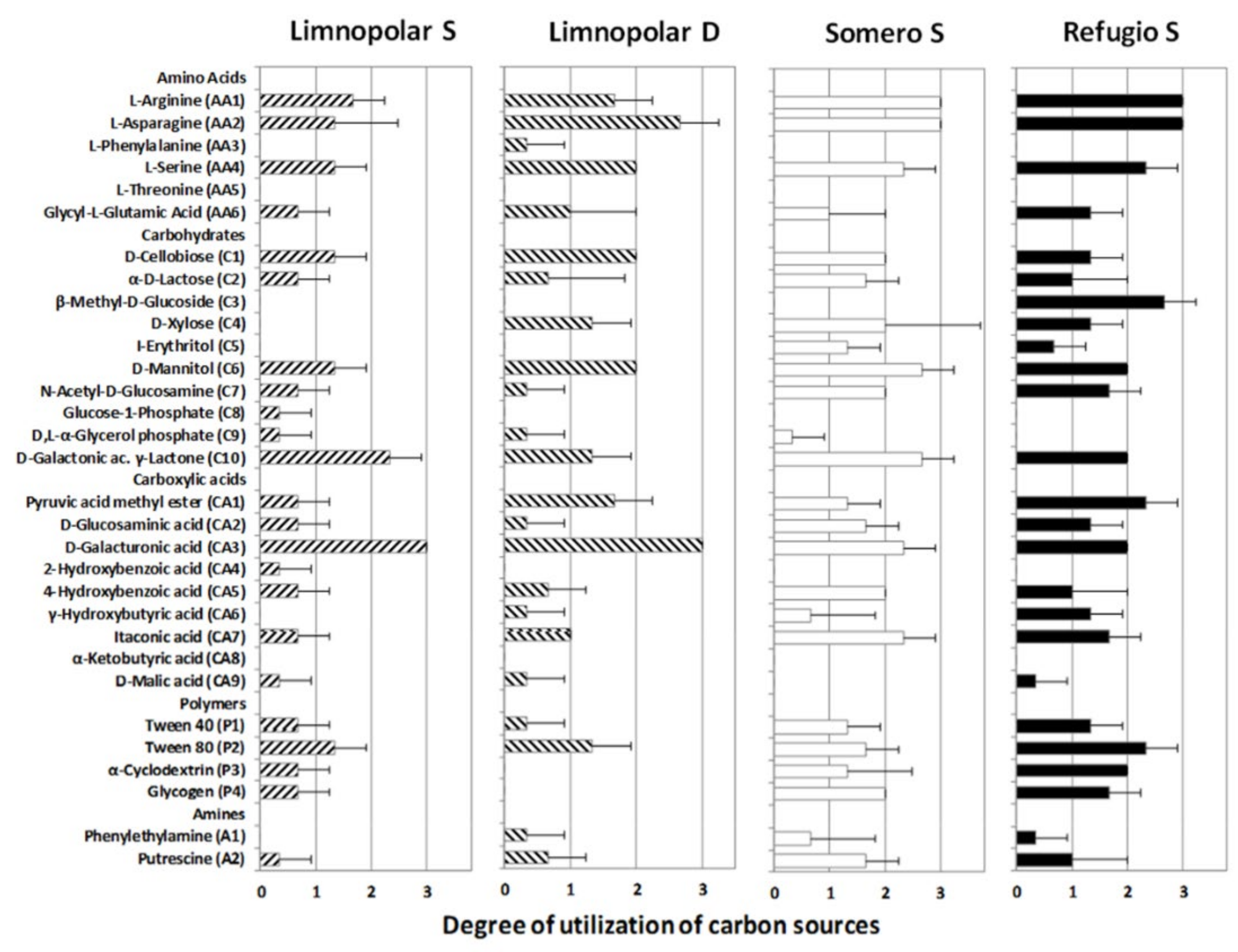
3.2. Carbon-Substrate Utilization Profiles
3.3. Statistical Analyses and Diversity Parameters
3.4. Co-Occurrence Network of Bacterioplankton Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pearce, D.; Van Der Gast, C.; Lawley, B.; Ellis-Evans, J. Bacterioplankton community diversity in a maritime Antarctic lake, determined by culture-dependent and culture-independent techniques. FEMS Microbiol. Ecol. 2003, 45, 59–70. [Google Scholar] [CrossRef]
- Reche, I.; Pulido, E.; Baquero, R.M.; Casamayor, E.O. Does ecosystem size determine aquatic bacterial richness? Ecology 2005, 86, 1715–1722. [Google Scholar] [CrossRef]
- Schiaffino, M.R.; Unrein, F.; Gasol, J.M.; Farias, M.E.; Estevez, C.; Balagué, V.; Izaguirre, I. Comparative analysis of bacterioplankton assemblages from maritime Antarctic freshwater lakes with contrasting trophic status. Polar Biol. 2009, 32, 923–936. [Google Scholar] [CrossRef]
- Picazo, A.; Rochera, C.; Villaescusa, J.A.; Miralles-Lorenzo, J.; Velázquez, D.; Quesada, A.; Camacho, A. Bacterioplankton community composition along environmental gradients in lakes from Byers Peninsula (Maritime Antarctica) as determined by next-generation sequencing. Front. Microbiol. 2019, 10, 908. [Google Scholar] [CrossRef]
- Rochera, C.; Camacho, A. Limnology and aquatic microbial ecology of Byers Peninsula: A Main freshwater biodiversity hotspot in Maritime Antarctica. Diversity 2019, 11, 201. [Google Scholar] [CrossRef]
- Ellis-Evans, J.C. Microbial diversity and function in Antarctic freshwater ecosystems. Biodivers. Conserv. 1996, 5, 1395–1431. [Google Scholar] [CrossRef]
- Laybourn-Parry, J.; Quayle, W.C.; Henshaw, T.; Ruddell, A.; Marchant, H.J. Life on the edge: The plankton and chemistry of Beaver Lake, an ultra-oligotrophic epishelf lake, Antarctica. Freshw. Biol. 2001, 46, 1205–1217. [Google Scholar] [CrossRef]
- Laybourn-Parry, J. Survival mechanisms in Antarctic lakes. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 863–869. [Google Scholar] [CrossRef]
- Laybourn-Parry, J. No place too cold. Science 2009, 324, 1521–1522. [Google Scholar] [CrossRef] [PubMed]
- Michaud, L.; Caruso, C.; Mangano, S.; Interdonato, F.; Bruni, V.; Giudice, A.L. Predominance of Flavobacterium, Pseudomonas, and Polaromonas within the prokaryotic community of freshwater shallow lakes in the northern Victoria Land, East Antarctica. FEMS Microbiol. Ecol. 2012, 82, 391–404. [Google Scholar] [CrossRef]
- Camacho, A. Planktonic microbial assemblages and the potential effects of metazooplankton predation on the food web of lakes from the maritime Antarctica and sub-Antarctic islands. Rev. Environ. Sci. Bio/Technol. 2006, 5, 167–185. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Raffaelli, D.; Schmid, B.; Tilman, D.; et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Srivastava, D.S.; Duffy, J.E.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nat. Cell Biol. 2006, 443, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Jacques, S.M.; Pires, A.P.F.; Leal, J.S.; Srivastava, D.S.; Parfrey, L.W.; Farjalla, V.; Doebeli, M. High taxonomic variability despite stable functional structure across microbial communities. Nat. Ecol. Evol. 2017, 1, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PIC-RUSt2: An improved and customizable approach for metagenome inference. bioRxiv 2020, 672295. [Google Scholar] [CrossRef]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Spear, J.R.; Caporaso, G.; Blekhman, R.; Knight, R.; et al. BugBase predicts organism-level microbiome phenotypes. bioRxiv 2017, 133462. [Google Scholar] [CrossRef]
- Barberán, A.; Velazquez, H.C.; Jones, S.; Fierer, N. Hiding in plain sight: Mining bacterial species records for phenotypic trait information. mSphere 2017, 2, e00237-17. [Google Scholar] [CrossRef] [PubMed]
- Galand, P.E.; Pereira, O.; Hochart, C.; Auguet, J.C.; Debroas, D. A strong link between marine microbial community composition and function challenges the idea of functional redundancy. ISME J. 2018, 12, 2470–2478. [Google Scholar] [CrossRef]
- Yan, D.; Xia, P.; Song, X.; Lin, T.; Cao, H. Community structure and functional diversity of epiphytic bacteria and planktonic bacteria on submerged macrophytes in Caohai Lake, southwest of China. Ann. Microbiol. 2019, 69, 933–944. [Google Scholar] [CrossRef]
- Yang, J.; Jiang, H.; Sun, X.; Huang, J.; Han, M.; Wang, B. Distinct co-occurrence patterns of prokaryotic community between the waters and sediments in lakes with different salinity. FEMS Microbiol. Ecol. 2020, 97, fiaa234. [Google Scholar] [CrossRef] [PubMed]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Ge, Z.; Poudel, D.R. Application and optimization of Biolog EcoPlates in functional diversity studies of soil microbial communities. MATEC Web Conf. 2015, 22, 04015. [Google Scholar] [CrossRef]
- Chróst, R.J.; Münster, U.; Rai, H.; Albrecht, D.; Witzel, P.K.; Overbeck, J. Photosynthetic production and exoenzymatic degradation of organic matter in the euphotic zone of a eutrophic lake. J. Plankton Res. 1989, 11, 223–242. [Google Scholar] [CrossRef]
- Jãrgensen, N.O.; Jensen, R.E.; Jørgensen, N.O. Microbial fluxes of free monosaccharides and total carbohydrates in freshwater determined by PAD-HPLC. FEMS Microbiol. Ecol. 1994, 14, 79–93. [Google Scholar] [CrossRef]
- Choi, K.-H.; Dobbs, F.C. Comparison of two kinds of Biolog microplates (GN and ECO) in their ability to distinguish among aquatic microbial communities. J. Microbiol. Methods 1999, 36, 203–213. [Google Scholar] [CrossRef]
- Grzesiak, J.; Zdanowski, M.K.; Górniak, D.; Świątecki, A.; Aleksandrzak-Piekarczyk, T.; Szatraj, K.; Sasin-Kurowska, J.; Nieckarz, M. Microbial community changes along the Ecology Glacier ablation zone (King George Island, Antarctica). Polar Biol. 2015, 38, 2069–2083. [Google Scholar] [CrossRef]
- López-Martínez, J.; Serrano, E.; Schmid, T.; Mink, S.; Linés, C. Periglacial processes and landforms in the South Shetland Islands (northern Antarctic Peninsula region). Geomorphology 2012, 155, 62–79. [Google Scholar] [CrossRef]
- Rochera, C.; Justel, A.; Fernández-Valiente, E.; Bañón, M.; Rico, E.; Toro, M.; Camacho, A.; Quesada, A. Interannual meteorological variability and its effects on a lake from maritime Antarctica. Polar Biol. 2010, 33, 1615–1628. [Google Scholar] [CrossRef]
- Bañón, M.; Justel, A.; Velázquez, D.; Quesada, A. Regional weather survey on Byers Peninsula, Livingston Island, South Shetland Islands, Antarctica. Antarct. Sci. 2013, 25, 146–156. [Google Scholar] [CrossRef]
- Van Lipzig, N.P.M.; King, J.C.; Lachlan-Cope, T.A.; Broeke, M.R. Precipitation, sublimation, and snow drift in the Antarctic Peninsula region from a regional atmospheric model. J. Geophys. Res. Space Phys. 2004, 109, 1–16. [Google Scholar] [CrossRef]
- Toro, M.; Camacho, A.; Rochera, C.; Rico, E.; Bañón, M.; Fernández-Valiente, E.; Marco, E.; Justel, A.; Avendaño, M.C.; Ariosa, Y.; et al. Limnological characteristics of the freshwater ecosystems of Byers Peninsula, Livingston Island, in maritime Antarctica. Polar Biol. 2007, 30, 635–649. [Google Scholar] [CrossRef]
- Oliva, M.; Hrbáček, F.; Ruiz-Fernández, J.; de Pablo, M.Á.; Vieira, G.; Ramos, M.; Antoniades, D. Active layer dynamics in three topographically distinct lake catchments in Byers Peninsula (Livingston Island, Antarctica). Catena 2017, 149, 548–559. [Google Scholar] [CrossRef]
- Rochera, C.; Toro, M.; Rico, E.; Fernández-Valiente, E.; Villaescusa, J.A.; Picazo, A.; Quesada, A.; Camacho, A. Structure of planktonic microbial communities along a trophic gradient in lakes of Byers Peninsula, South Shetland Islands. Antarct. Sci. 2013, 25, 277–287. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Casamayor, E.O.; Rochera, C.; Velázquez, D.; Chicote, A.; Quesada, A.; Camacho, A. A close link between bacterial community composition and environmental heterogeneity in maritime Antarctic lakes. Int. Microbiol. 2010, 13, 67–77. [Google Scholar] [CrossRef]
- Gil-Delgado, J.A.; Villaescusa, J.A.; Diazmacip, M.E.; Velazquez, D.; Rico, E.; Toro, M.; Quesada, A.; Camacho, A. Minimum population size estimates demonstrate an increase in southern elephant seals (Mirounga leonina) on Livingston Island, maritime Antarctica. Polar Biol. 2012, 36, 607–610. [Google Scholar] [CrossRef]
- Moura, P.A.; Francelino, M.R.; Schaefer, C.E.G.; Simas, F.N.; de Mendonça, B.A. Distribution and characterization of soils and landform relationships in Byers Peninsula, Livingston Island, Maritime Antarctica. Geomorphology 2012, 155, 45–54. [Google Scholar] [CrossRef]
- Santamans, A.C.; Boluda, R.; Picazo, A.; Gil, C.; Ramos-Miras, J.J.; Tejedo, P.; Pertierra, L.R.; Benayas, J.; Camacho, A. Soil features in rookeries of Antarctic penguins reveal sea to land biotransport of chemical pollutants. PLoS ONE 2017, 12, e0181901. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org/ (accessed on 1 May 2021).
- Shannon, C.E.; Weaver, W. The Measurement Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Whittaker, R.H. Evolution of species diversity in land communities. Evol. Biol. 1977, 10, 1–67. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Christian, B.W.; Lind, O.T. Multiple carbon substrate utilization by bacteria at the sediment–water interface: Seasonal patterns in a stratified eutrophic reservoir. Hydrobiologia 2007, 586, 43–56. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Faust, K.; Raes, J. Conet app: Inference of biological association networks using cytoscape. F1000Research 2016, 5, 1519. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.T.; Franz, M.; Kazi, F.; Donaldson, S.L.; Morris, Q.; Bader, G.D. Cytoscape Web: An interactive web-based network browser. Bioinformatics 2010, 26, 2347–2348. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef]
- Mackey, T.J.; Sumner, D.Y.; Hawes, I.; Jungblut, A.D.; Andersen, D. Growth of modern branched columnar stromatolites in Lake Joyce, Antarctica. Geobiology 2015, 13, 373–390. [Google Scholar] [CrossRef]
- Koo, H.; Mojib, N.; Hakim, J.A.; Hawes, I.; Tanabe, Y.; Andersen, D.T.; Bej, A.K. Microbial communities and their predicted metabolic functions in growth laminae of a unique large conical mat from Lake Untersee, East Antarctica. Front. Microbiol. 2017, 8, 1347. [Google Scholar] [CrossRef]
- Song, Y.; Wang, Y.; Mao, G.; Gao, G.; Wang, Y. Impact of planktonic low nucleic acid-content bacteria to bacterial community structure and associated ecological functions in a shallow lake. Sci. Total. Environ. 2019, 658, 868–878. [Google Scholar] [CrossRef]
- Camacho, A. Sulfur bacteria. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Elsevier Academic Press: Oxford, NY, USA, 2009; pp. 261–278. [Google Scholar] [CrossRef]
- Fernández-Valiente, E.; Camacho, A.; Rochera, C.; Rico, E.; Vincent, W.; Quesada, A. Community structure and physiological characterization of microbial mats in Byers Peninsula, Livingston Island (South Shetland Islands, Antarctica). FEMS Microbiol. Ecol. 2007, 59, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, D.; Jungblut, A.D.; Rochera, C.; Rico, E.; Camacho, A.; Quesada, A. Trophic interactions in microbial mats on Byers Peninsula, maritime Antarctica. Polar Biol. 2016, 40, 1115–1126. [Google Scholar] [CrossRef]
- Barbaro, E.; Zangrando, R.; Vecchiato, M.; Turetta, C.; Barbante, C.; Gambaro, A. D- and l-amino acids in Antarctic lakes: Assessment of a very sensitive HPLC-MS method. Anal. Bioanal. Chem. 2014, 406, 5259–5270. [Google Scholar] [CrossRef] [PubMed]
- Villaescusa, J.A.; O’ Casamayor, E.; Rochera, C.; Quesada, A.; Michaud, L.; Camacho, A. Heterogeneous vertical structure of the bacterioplankton community in a non-stratified Antarctic Lake. Antarct. Sci. 2013, 25, 229–238. [Google Scholar] [CrossRef]
- Schellenberger, S.; Drake, H.L.; Kolb, S. Functionally Redundant cellobiose-degrading soil bacteria respond differentially to oxygen. Appl. Environ. Microbiol. 2011, 77, 6043–6048. [Google Scholar] [CrossRef][Green Version]
- Nichols, D.; Bowman, J.; Sanderson, K.; Nichols, C.M.; Lewis, T.; McMeekin, T.; Nichols, P.D. Developments with Antarctic microorganisms: Culture collections, bioactivity screening, taxonomy, PUFA production and cold-adapted enzymes. Curr. Opin. Biotechnol. 1999, 10, 240–246. [Google Scholar] [CrossRef]
- Budnick, W.R.; Mruzek, J.L.; Larson, C.A.; Passy, S.I. The impacts of nutrient supply and imbalance on subcontinental co-occurrence networks and metacommunity composition of stream algae. Ecography 2021, 44, 1109–1120. [Google Scholar] [CrossRef]
- Zhao, D.; Shen, F.; Zeng, J.; Huang, R.; Yu, Z.; Wu, Q.L. Network analysis reveals seasonal variation of co-occurrence correlations between Cyanobacteria and other bacterioplankton. Sci. Total Environ. 2016, 573, 817–825. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | X-UTM | Y-UTM | Catchment Size (km2) | Lake Surface (km2) | Maximum Depth (m) | Analyses Conducted | Trophic Status |
|---|---|---|---|---|---|---|---|
| Refugio | 602200 | 3050550 | 0.12 | 0.016 | 0.5 | F, B | Eutrophic |
| Turbio | 598000 | 3051800 | 0.58 | 0.021 | 7.8 | F | Oligotrophic |
| Escondido | 599475 | 3052650 | 0.08 | 0.022 | 4.5 | F | Ultra-oligotrophic |
| Somero | 596800 | 3052150 | 0.06 | 0.011 | 0.5 | F, B | Mesotrophic |
| Midge | 597700 | 3054150 | 0.27 | 0.054 | 8.2 | F | Ultra-oligotrophic |
| Chester | 597500 | 3053550 | 0.09 | 0.039 | 5.0 | F | Ultra-oligotrophic |
| Limnopolar | 597100 | 3052200 | 0.58 | 0.023 | 5.5 | F, B | Oligotrophic |
| Refugio S | Turbio S | Escondido S | Somero S | Midge S | Chester S | Chester D | Limnopolar S | Limnopolar D | ||
|---|---|---|---|---|---|---|---|---|---|---|
| FAPROTAX | Chao-1 | 20 | 8 | 15 | 6 | 15 | 12 | 14 | 8 | 9 |
| Shannon H | 2.00 | 1.64 | 1.53 | 0.75 | 1.75 | 1.55 | 1.97 | 1.41 | 1.56 | |
| Evenness J | 0.67 | 0.790 | 0.56 | 0.42 | 0.65 | 0.62 | 0.750 | 0.68 | 0.71 | |
| Menhinick | 1.07 | 0.22 | 0.43 | 0.17 | 0.52 | 0.47 | 0.40 | 0.240 | 0.26 | |
| C:N:S ratio | 3:0.9:1 | 60:41:1 | 65:6:1 | 15:0.8:1 | 18:4:1 | 8:3:1 | 3:0.4:1 | 30:10:1 | 3:1:1 | |
| BIOLOG | Richness S | 25 | - | - | 23 | - | - | - | 23 | 23 |
| Shannon H | 3.12 | - | - | 3.09 | - | - | - | 2.93 | 2.89 | |
| Evenness J | 0.97 | - | - | 0.99 | - | - | - | 0.93 | 0.92 | |
| AWCD | 1.32 | - | - | 1.39 | - | - | - | 0.71 | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Picazo, A.; Villaescusa, J.A.; Rochera, C.; Miralles-Lorenzo, J.; Quesada, A.; Camacho, A. Functional Metabolic Diversity of Bacterioplankton in Maritime Antarctic Lakes. Microorganisms 2021, 9, 2077. https://doi.org/10.3390/microorganisms9102077
Picazo A, Villaescusa JA, Rochera C, Miralles-Lorenzo J, Quesada A, Camacho A. Functional Metabolic Diversity of Bacterioplankton in Maritime Antarctic Lakes. Microorganisms. 2021; 9(10):2077. https://doi.org/10.3390/microorganisms9102077
Chicago/Turabian StylePicazo, Antonio, Juan Antonio Villaescusa, Carlos Rochera, Javier Miralles-Lorenzo, Antonio Quesada, and Antonio Camacho. 2021. "Functional Metabolic Diversity of Bacterioplankton in Maritime Antarctic Lakes" Microorganisms 9, no. 10: 2077. https://doi.org/10.3390/microorganisms9102077
APA StylePicazo, A., Villaescusa, J. A., Rochera, C., Miralles-Lorenzo, J., Quesada, A., & Camacho, A. (2021). Functional Metabolic Diversity of Bacterioplankton in Maritime Antarctic Lakes. Microorganisms, 9(10), 2077. https://doi.org/10.3390/microorganisms9102077