Probiotics for the Treatment of Overweight and Obesity in Humans—A Review of Clinical Trials

Abstract
:1. Introduction
2. Methods
2.1. Data Sources and Searches
2.2. Eligibility Criteria
2.3. Quality Assessment
3. Results
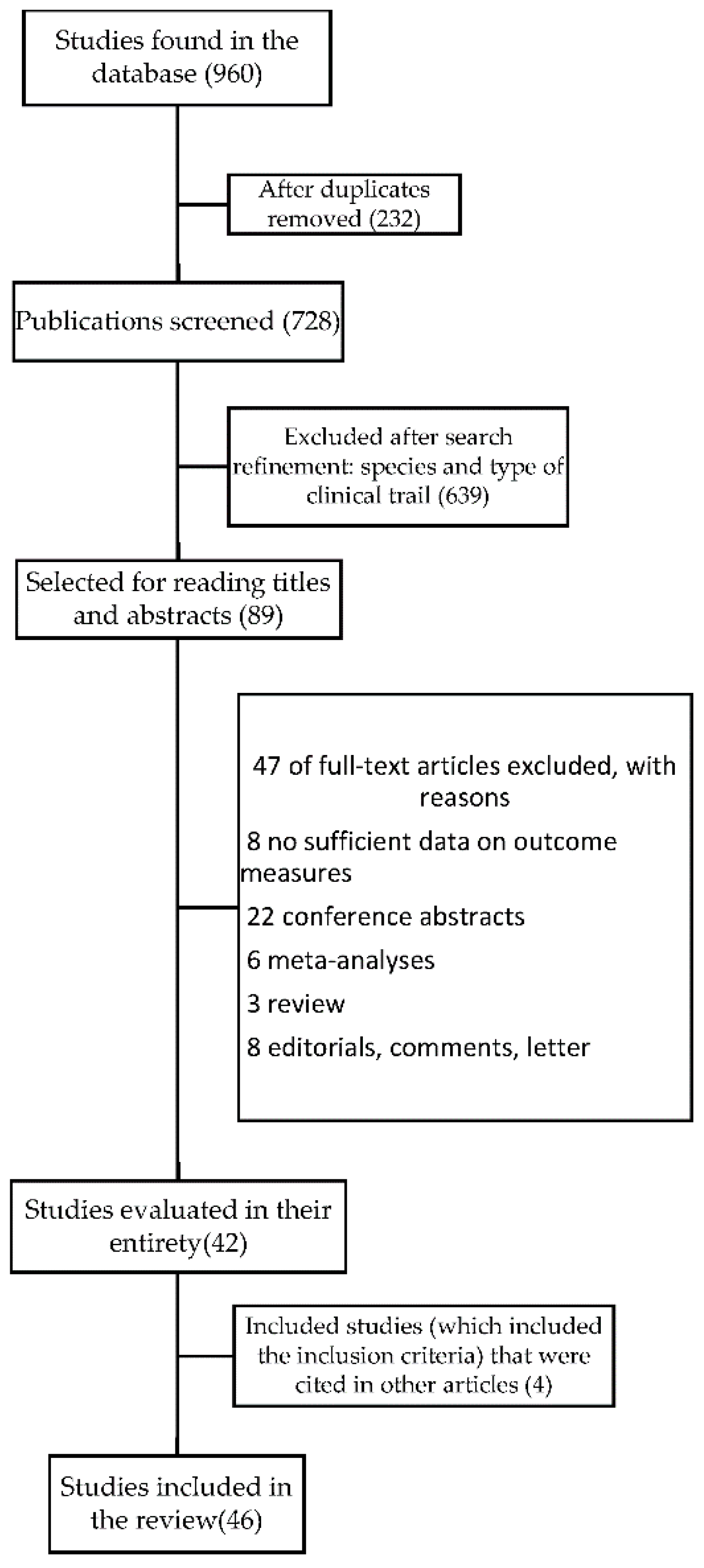
3.1. Search Results
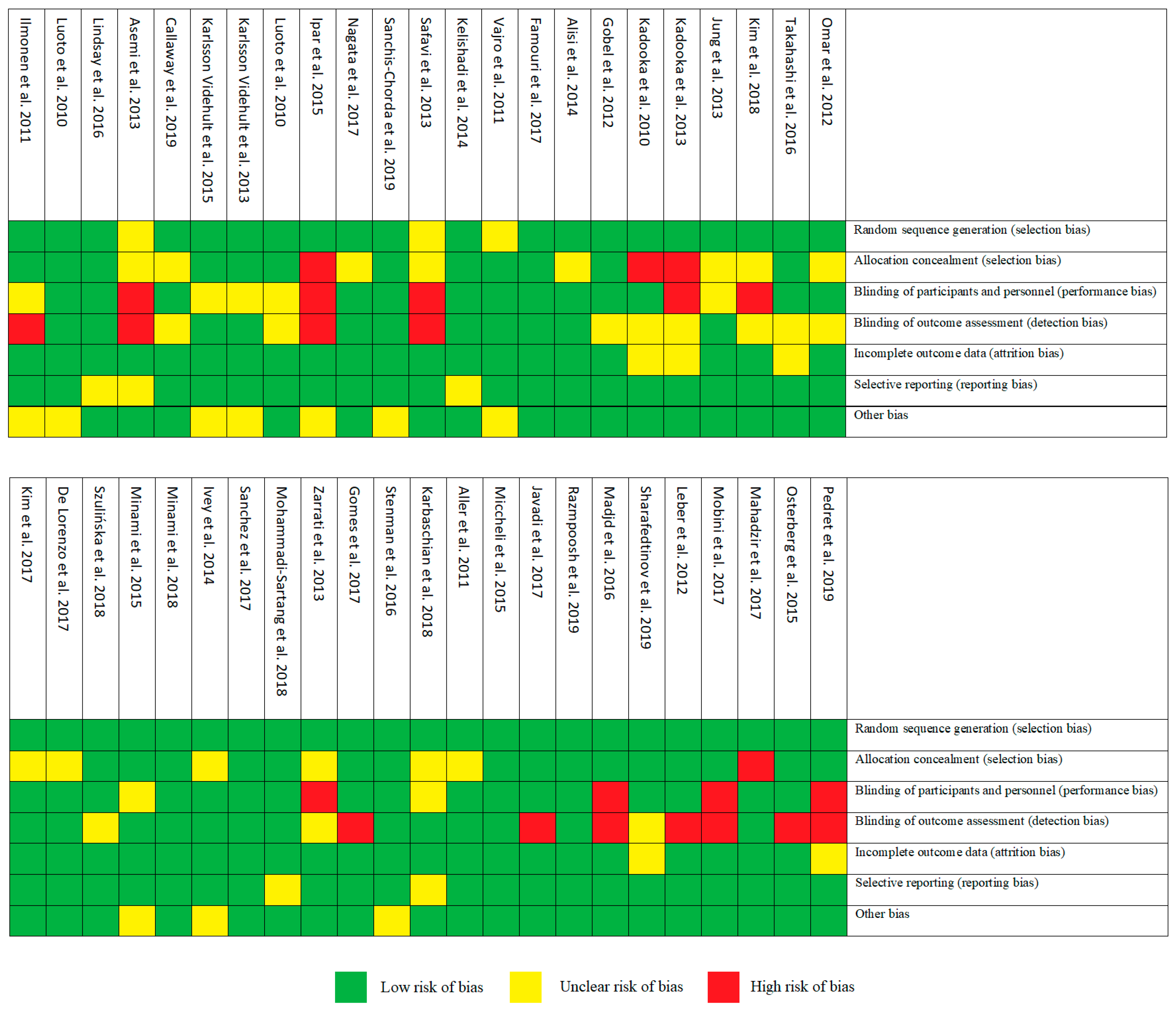
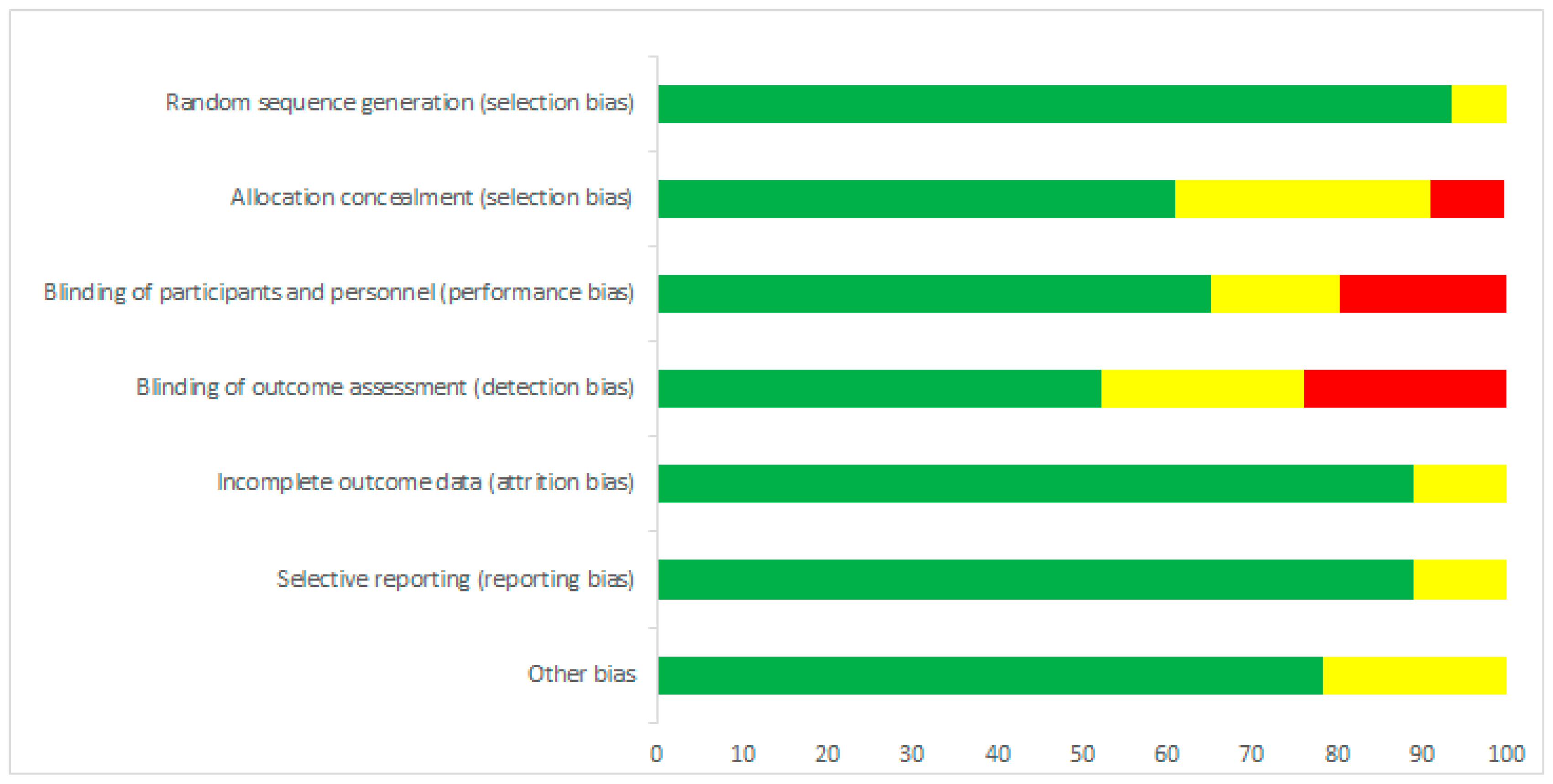
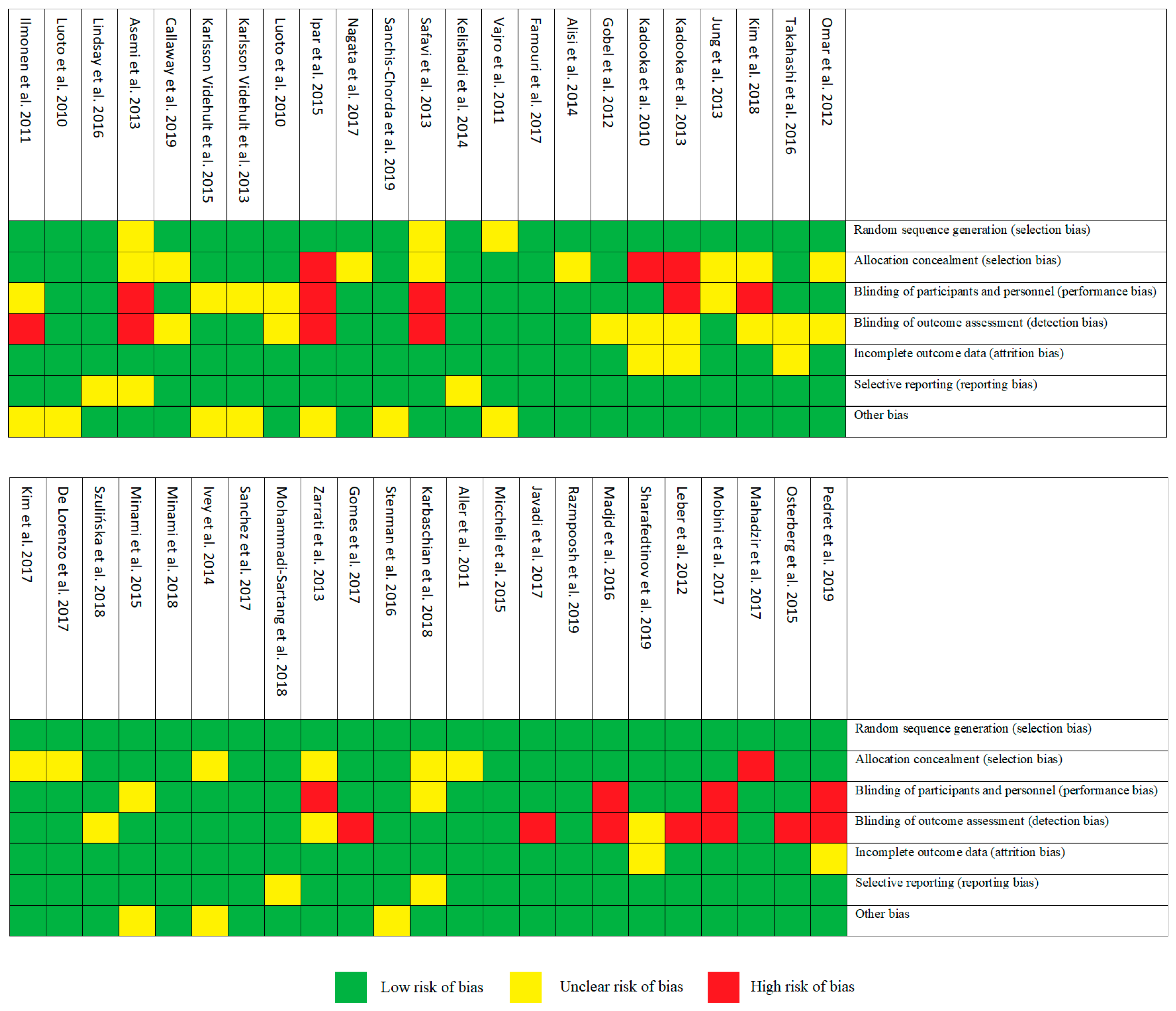
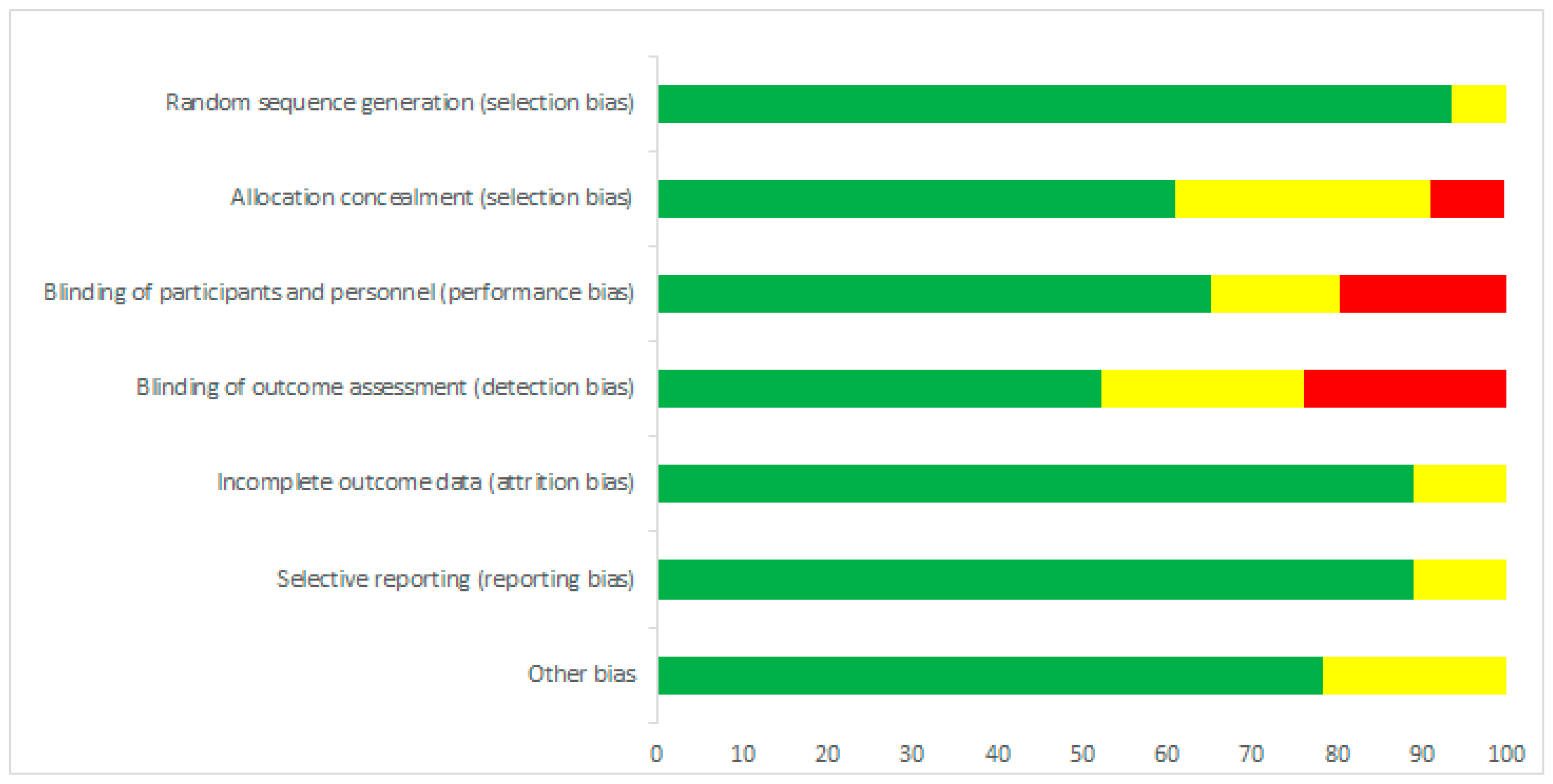
3.2. Risk of Bias Assessment
4. Obesity
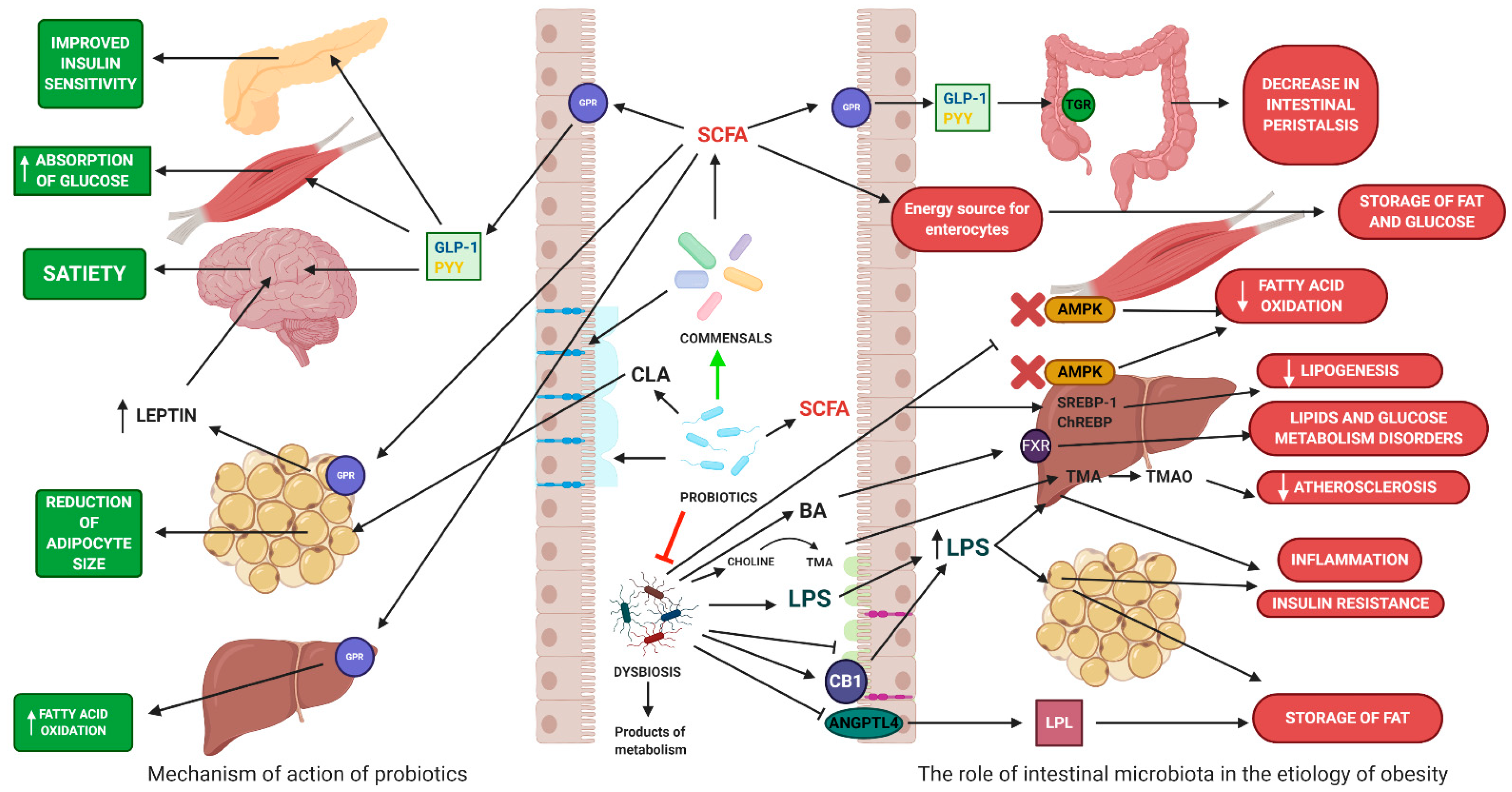
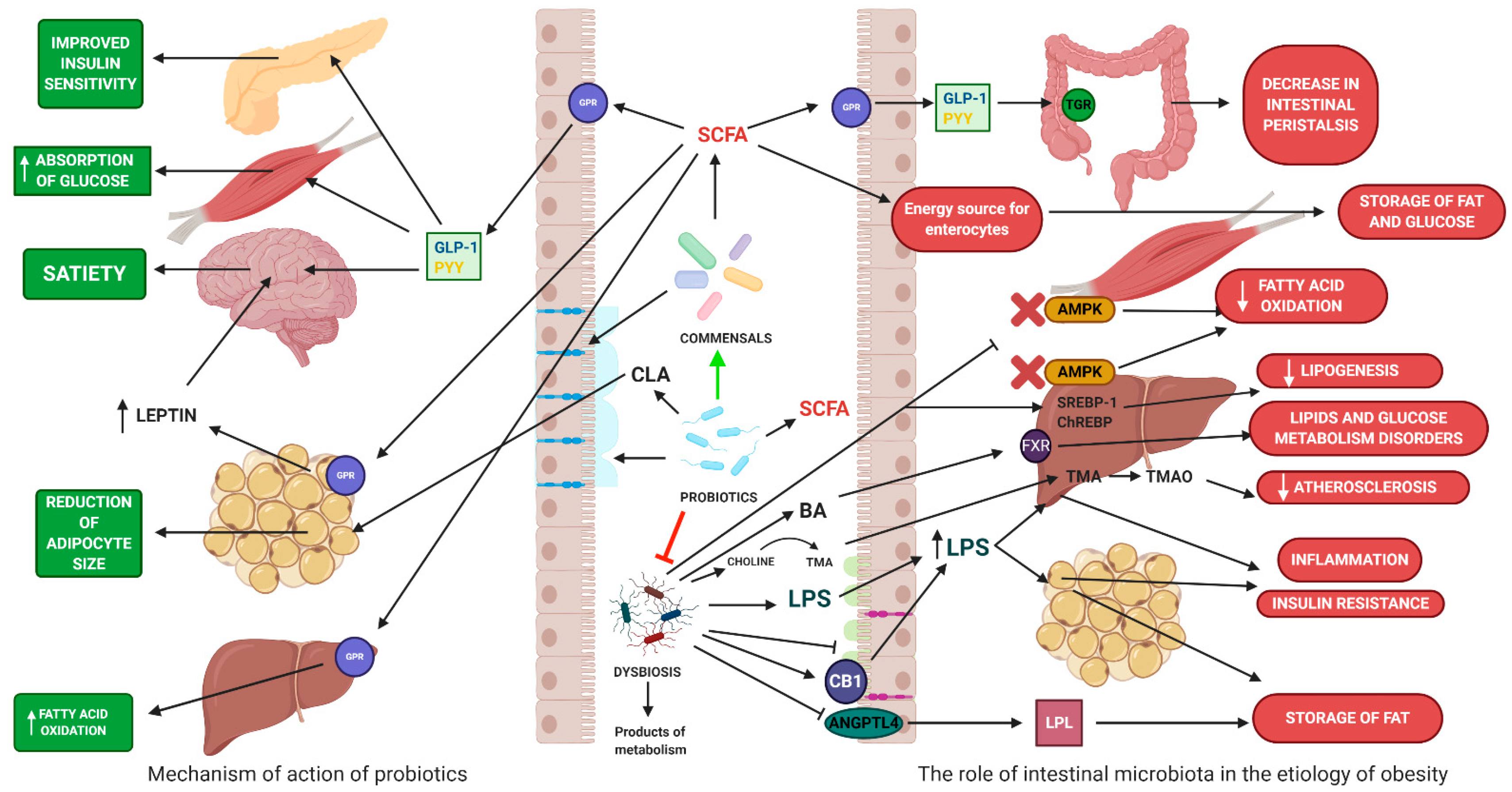
4.1. The Role of Gut Microbiota in Obesity
4.2. The Impact on Energy Homeostasis
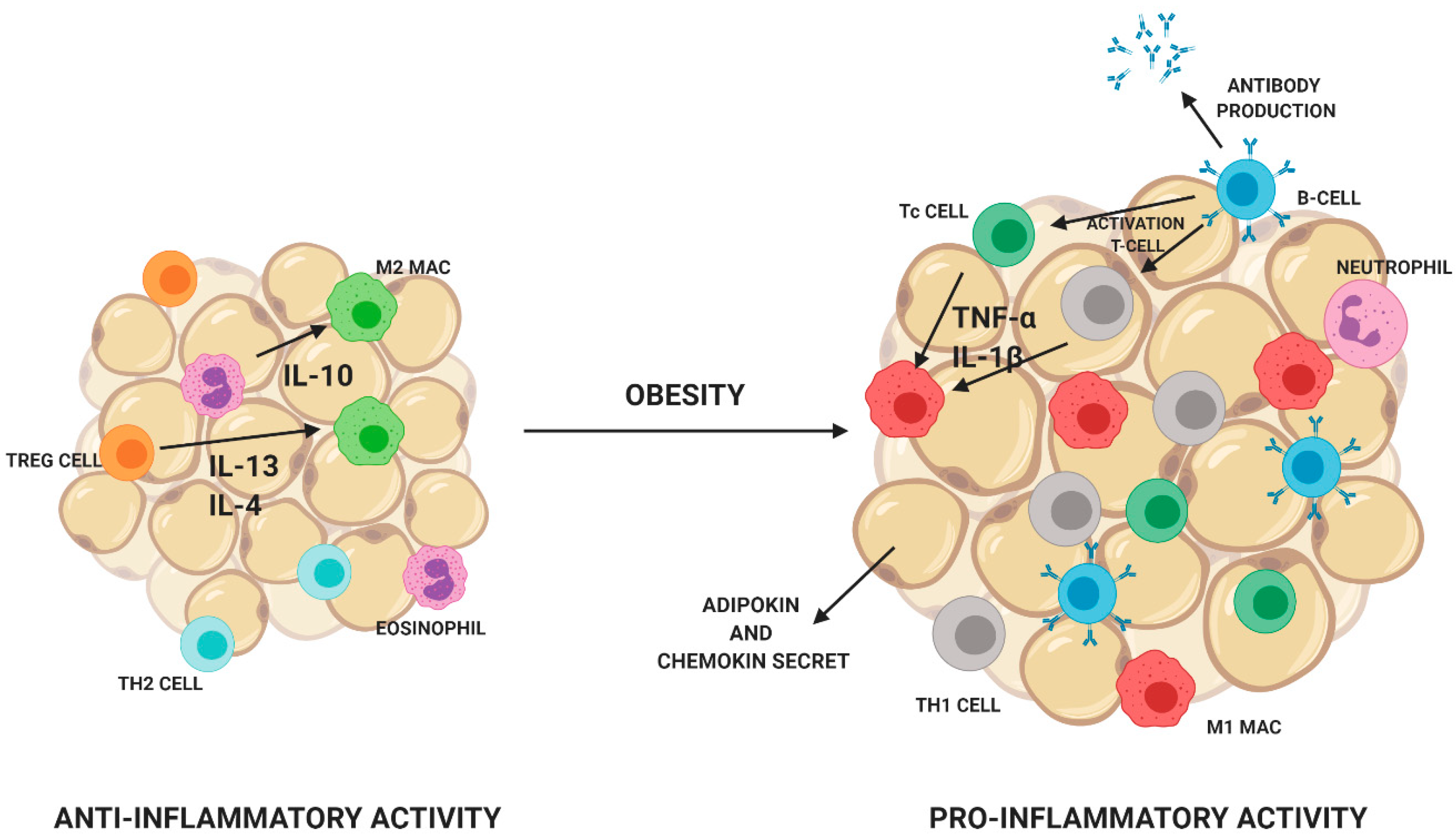
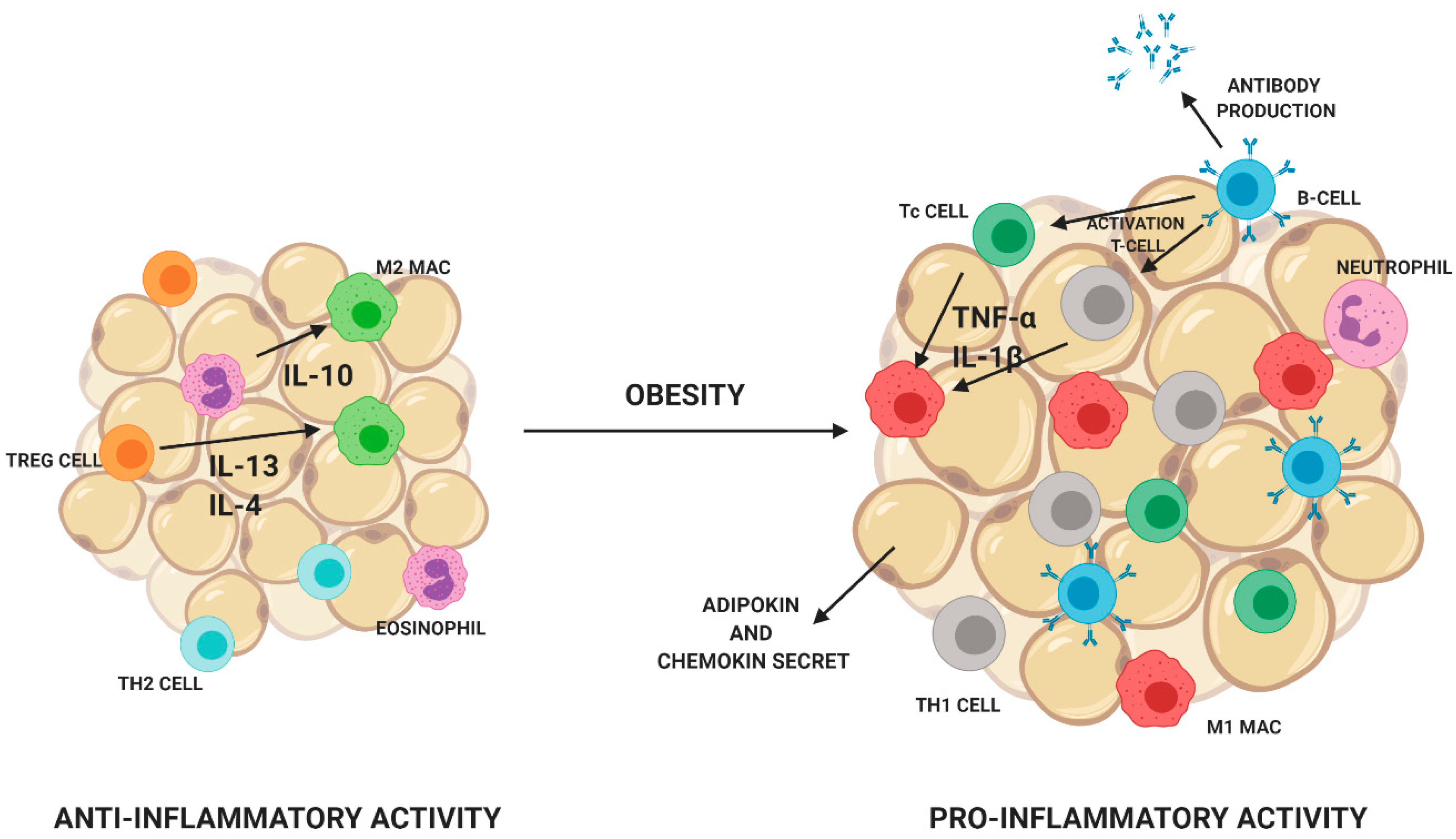
4.3. The Impact on Inflammatory Processes
4.4. The Role of Probiotics in the Treatment and Prevention of Obesity
5. Review of Clinical Studies Using Probiotics in Obesity
5.1. The Effect of Probiotic Supplementation in Pregnant Women on Body Weight
5.2. The Effect of Probiotic Supplementation in Infants in the Prevention of Overweight or Obesity
5.3. The Effect of Probiotic Supplementation in Children on Body Weight
5.4. The Effect of Probiotic Supplementation in Adults on Body Weight
5.5. The Effect of Heat Killed Probiotics on Body Weight
5.6. The Effect of Probiotic Supplementation on Weight Gain
6. Meta-Analyses
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Checchi, F.; Waldman, R.J.; Roberts, L.F.; Ager, A.; Asgary, R.; Benner, M.T.; Blanchet, K.; Burnham, G.; D’Harcourt, E.; Leaning, J.; et al. World Health Organization and emergency health: If not now, when? BMJ 2016, 352, i469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuttall, F.Q. Body Mass Index: Obesity, BMI, and Health: A Critical Review. Nutr. Today 2015, 50, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.B.; Smith, M.S. Obesity Statistics. Prim. Care Clin. Off. Pract. 2016, 43, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Altman, D.G.; Sterne, J.A.C. Cochrane Handbook for Systematic Reviews of Interventions; Higgins, J.P.T., Green, S., Eds.; Version 5.1.0; The Cochrane Collaboration: London, UK, 2011; Chapter 8. [Google Scholar]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2010, 12, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-J.J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Sherman, M.P.; Zaghouani, H.; Niklas, V. Gut microbiota, the immune system, and diet influence the neonatal gut–brain axis. Pediatr. Res. 2014, 77, 127–135. [Google Scholar] [CrossRef]
- Forbes, J.D.; Van Domselaar, G.; Bernstein, C.N. The Gut Microbiota in Immune-Mediated Inflammatory Diseases. Front. Microbiol. 2016, 7, 1081. [Google Scholar] [CrossRef] [Green Version]
- Buechler, C.; Wanninger, J.; Neumeier, M. Adiponectin, a key adipokine in obesity related liver diseases. World J. Gastroenterol. 2011, 17, 2801–2811. [Google Scholar]
- Raucci, R.; Rusolo, F.; Sharma, A.; Colonna, G.; Castello, G.; Costantini, S. Functional and structural features of adipokine family. Cytokine 2013, 61, 1–14. [Google Scholar] [CrossRef]
- Kang, Y.S. Obesity Associated Hypertension: New Insights into Mechanism. Electrolytes Blood Press. 2013, 11, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Miyata, Y.; Tanaka, H.; Shimada, A.; Sato, T.; Ito, A.; Yamanouchi, T.; Kosano, H. Regulation of adipocytokine secretion and adipocyte hypertrophy by polymethoxyflavonoids, nobiletin and tangeretin. Life Sci. 2011, 88, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Ohashi, K.; Shibata, R.; Murohara, T. Adipocytokines and obesity-linked disorders. Nagoya J. Med. Sci. 2012, 74, 19–30. [Google Scholar] [PubMed]
- Cao, H. Adipocytokines in obesity and metabolic disease. J. Endocrinol. 2014, 220, T47–T59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Ji, Y.; Kersten, S.; Qi, L. Mechanisms of inflammatory responses in obese adipose tissue. Annu. Rev. Nutr. 2012, 32, 261–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Wiciński, M.; Gębalski, J.; Mazurek, E.; Podhorecka, M.; Śniegocki, M.; Szychta, P.; Sawicka, E.; Malinowski, B. The Influence of Polyphenol Compounds on Human Gastrointestinal Tract Microbiota. Nutrients 2020, 12, 350. [Google Scholar] [CrossRef] [Green Version]
- Bibbò, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The role of diet on gut microbiota composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar]
- Hill, C.J.; De Filippis, F.; Jeffery, I.B. Diet, Health, and the Gut Microbiota. Food Microbiol. 2019, 815–829. [Google Scholar] [CrossRef]
- Carding, S.R.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 3854. [Google Scholar] [CrossRef]
- Hannson, G.C. Role of mucus layers in gut infection and inflammation. Curr. Opin. Microbiol. 2012, 15, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Aversa, F.; Tortora, A.; Ianiro, G.; Ponziani, F.R.; Annicchiarico, B.E.; Gasbarrini, A. Gut microbiota and metabolic syndrome. Intern. Emerg. Med. 2013, 8, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Sanz, Y.; Santacruz, A.; Gauffin, P. Gut microbiota in obesity and metabolic disorders. Proc. Nutr. Soc. 2010, 69, 434–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gérard, P. Gut microbiota and obesity. Cell Mol. Life Sci. 2016, 73, 147–162. [Google Scholar] [CrossRef]
- Scott, K.P.; Gratz, S.W.; Sheridan, P.; Flint, H.J.; Duncan, S.H. The influence of diet on the gut microbiota. Pharmacol. Res. 2013, 69, 52–60. [Google Scholar] [CrossRef]
- Biedermann, L.; Rogler, G. The intestinal microbiota: Its role in health and disease. Eur. J. Nucl. Med. Mol. Imaging 2015, 174, 151–167. [Google Scholar] [CrossRef]
- Besten, G.D.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Barja-Fernández, S.; Folgueira, C.; Castelao, C.; Pena-León, V.; González-Saenz, P.; Vázquez-Cobela, R.; Aguilera, C.M.; Gil-Campos, M.; Bueno, G.; Gil, A.; et al. ANGPTL-4 is Associated with Obesity and Lipid Profile in Children and Adolescents. Nutrients 2019, 11, 1340. [Google Scholar] [CrossRef] [Green Version]
- Kamada, N.; Seo, S.-U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; Reyes-Gavilán, C.G.D.L.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Layden, B.T.; Angueira, A.R.; Brodsky, M.; Durai, V.; Lowe, W.L. Short chain fatty acids and their receptors: New metabolic targets. Transl. Res. 2013, 161, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xie, C.; Wang, G.; Wu, Y.; Wu, Q.; Wang, X.; Liu, J.; Deng, Y.-Y.; Xia, J.; Chen, B.; et al. Gut microbiota and intestinal FXR mediate the clinical benefits of metformin. Nat. Med. 2018, 24, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yang, L.; Wang, Z.; Huang, W. Bile acid nuclear receptor FXR and digestive system diseases. Acta Pharm. Sin. B 2015, 5, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Düfer, M.; Hörth, K.; Krippeit-Drews, P.; Drews, G. The significance of the nuclear farnesoid X receptor (FXR) in β cell function. Islets 2012, 4, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Parséus, A.; Sommer, N.; Sommer, F.; Caesar, R.; Molinaro, A.; Ståhlman, M.; Greiner, T.U.; Perkins, R.; Bäckhed, F. Microbiota-induced obesity requires farnesoid X receptor. Gut 2016, 66, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Horai, Y.; Houten, S.M.; Morimoto, K.; Sugizaki, T.; Arita, E.; Mataki, C.; Sato, H.; Tanigawara, Y.; Schoonjans, K.; et al. Lowering Bile Acid Pool Size with a Synthetic Farnesoid X Receptor (FXR) Agonist Induces Obesity and Diabetes through Reduced Energy Expenditure. J. Biol. Chem. 2011, 286, 26913–26920. [Google Scholar] [CrossRef] [Green Version]
- Shyng, S.-L. Targeting the Gut Microbiota-FXR Signaling Axis for Glycemic Control: Does a Dietary Supplement Work Magic? Diabetes 2017, 66, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Zwickier, B.L.; Agellon, L.B. Transport and biological activities of bile acids. Int. J. Biochem. Cell Biol. 2013, 45, 1389–1398. [Google Scholar] [CrossRef]
- Cani, P.D. Gut microbiota and obesity: Lessons from the microbiome. Brief. Funct. Genom. 2013, 12, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, K.A.; Valentine, R.J.; Ruderman, N.B.; Saha, A.K. AMPK activation: A therapeutic target for type 2 diabetes? Diabetes Metab. Syndr. Obes. 2014, 7, 241–253. [Google Scholar] [PubMed] [Green Version]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nature 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Jeon, S.M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouter, K.E.; Van Raalte, D.H.; Groen, A.K.; Nieuwdorp, M. Role of the Gut Microbiome in the Pathogenesis of Obesity and Obesity-Related Metabolic Dysfunction. Gastroenterology 2017, 152, 1671–1678. [Google Scholar] [CrossRef]
- Dahiya, D.K.; Puniya, M.R.; Shandilya, U.K.; Dhewa, T.; Kumar, N.; Kumar, S.; Puniya, A.K.; Shukla, P. Gut Microbiota Modulation and Its Relationship with Obesity Using Prebiotic Fibers and Probiotics: A Review. Front. Microbiol. 2017, 8, 563. [Google Scholar] [CrossRef]
- Sato, R. Sterol metabolism and SREBP activation. Arch. Biochem. Biophys. 2010, 501, 177–181. [Google Scholar] [CrossRef]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: Mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef]
- Corbin, K.D.; Zeisel, S.H. Choline metabolism provides novel insights into nonalcoholic fatty liver disease and its progression. Curr. Opin. Gastroenterol. 2012, 28, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Spencer, M.D.; Hamp, T.J.; Reid, R.W.; Fischer, L.M.; Zeisel, S.H.; Fodor, A.A. Association Between Composition of the Human Gastrointestinal Microbiome and Development of Fatty Liver With Choline Deficiency. Gastroenterology 2011, 140, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Hur, K.Y.; Lee, M.-S. Gut Microbiota and Metabolic Disorders. Diabetes Metab. J. 2015, 39, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woting, A.; Blaut, M. The Intestinal Microbiota in Metabolic Disease. Nutrients 2016, 8, 202. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.; Cani, P.D. Interaction Between Obesity and the Gut Microbiota: Relevance in Nutrition. Annu. Rev. Nutr. 2011, 31, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborti, C.K. New-found link between microbiota and obesity. World J. Gastrointest. Pathophysiol. 2015, 6, 110–119. [Google Scholar] [CrossRef]
- Boutagy, N.E.; McMillan, R.P.; Frisard, M.; Hulver, M.W. Metabolic endotoxemia with obesity: Is it real and is it relevant? Biochimie 2015, 124, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Pereira, S.S.; Alvarez-Leite, J.I. Low-Grade Inflammation, Obesity, and Diabetes. Curr. Obes. Rep. 2014, 3, 422–431. [Google Scholar] [CrossRef]
- Zou, Z.-Y.; Hu, Y.-R.; Ma, H.; Wang, Y.-Z.; He, K.; Xia, S.; Wu, H.; Xue, D.-F.; Li, X.; Ye, X. Coptisine attenuates obesity-related inflammation through LPS/TLR-4-mediated signaling pathway in Syrian golden hamsters. Fitoterapia 2015, 105, 139–146. [Google Scholar] [CrossRef]
- Kim, K.-A.; Gu, W.; Lee, I.-A.; Joh, E.-H.; Kim, D.-H. High Fat Diet-Induced Gut Microbiota Exacerbates Inflammation and Obesity in Mice via the TLR4 Signaling Pathway. PLoS ONE 2012, 7, e47713. [Google Scholar] [CrossRef]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Alessandro, R.; Luchetti, M.M.; Milling, S.; Saieva, L.; Cypers, H.; Stampone, T.; di Benedetto, P.; et al. Dysbiosis and zonulin upregulation alter gut epithelial and vascular barriers in patients with ankylosing spondylitis. Ann. Rheum. Dis. 2017, 76, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Villa, C.R.; Ward, W.E.; Comelli, E.M. Gut microbiota-bone axis. Crit. Rev. Food Sci. Nutr. 2015, 57, 1664–1672. [Google Scholar] [CrossRef]
- Kim, K.-A.; Jeong, J.-J.; Yoo, S.-Y.; Kim, D.-H. Gut microbiota lipopolysaccharide accelerates inflamm-aging in mice. BMC Microbiol. 2016, 16, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muscogiuri, G.; Cantone, E.; Cassarano, S.; Tuccinardi, D.; Barrea, L.; Savastano, S.; Colao, A. Gut microbiota: A new path to treat obesity. Int. J. Obes. Suppl. 2019, 9, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Daniali, M.; Nikfar, S.; Abdollahi, M. A brief overview on the use of probiotics to treat overweight and obese patients. Expert Rev. Endocrinol. Metab. 2020, 15, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-B.; Xin, S.-S.; Ding, L.-N.; Ding, W.-Y.; Hou, Y.-L.; Liu, C.-Q.; Zhang, X. The Potential Role of Probiotics in Controlling Overweight/Obesity and Associated Metabolic Parameters in Adults: A Systematic Review and Meta-Analysis. Evid.-Based Complement. Altern. Med. 2019, 2019, 3862971. [Google Scholar] [CrossRef]
- Cerdó, T.; García-Santos, J.A.; García-Bermúdez, M.; Azcona-Sanjulián, M.C. The Role of Probiotics and Prebiotics in the Prevention and Treatment of Obesity. Nutrients 2019, 11, 635. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2018, 105, 16731–16736. [Google Scholar] [CrossRef] [Green Version]
- Wiciński, M.; Sawicka, E.; Gębalski, J.; Kubiak, K.; Malinowski, B. Human Milk Oligosaccharides: Health Benefits, Potential Applications in Infant Formulas, and Pharmacology. Nutrients 2020, 12, 266. [Google Scholar] [CrossRef] [Green Version]
- Badehnoosh, B.; Karamali, M.; Zarrati, M.; Jamilian, M.; Bahmani, F.; Tajabadi-Ebrahimi, M.; Jafari, P.; Rahmani, E.; Asemi, Z. The effects of probiotic supplementation on biomarkers of inflammation, oxidative stress and pregnancy outcomes in gestational diabetes. J. Matern. Neonatal Med. 2017, 31, 1128–1136. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhang, Q.; Ren, Y.; Ruan, Z. Effect of probiotic Lactobacillus on lipid profile: A systematic review and meta-analysis of randomized, controlled trials. PLoS ONE 2017, 12, e0178868. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.; Wang, C.; Zhao, H.; Zhao, C.; Chen, Y.; Wang, Y.; McClain, C.; Feng, W. Enhanced AMPK phosphorylation contributes to the beneficial effects of Lactobacillus rhamnosus GG supernatant on chronic-alcohol-induced fatty liver disease. J. Nutr. Biochem. 2014, 26, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Nova, E.; De Heredia, F.P.; Gómez-Martínez, S.; Marcos, A. The Role of Probiotics on the Microbiota. Nutr. Clin. Pract. 2016, 31, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrouz, V.; Jazayeri, S.; Aryaeian, N.; Zahedi, M.J.; Hosseini, F. Effects of Probiotic and Prebiotic Supplementation on Leptin, Adiponectin, and Glycemic Parameters in Non-alcoholic Fatty Liver Disease: A Randomized Clinical Trial. Middle East J. Dig. Dis. 2017, 9, 150–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilmonen, J.; Isolauri, E.; Poussa, T.; Laitinen, K. Impact of dietary counselling and probiotic intervention on maternal anthropometric measurements during and after pregnancy: A randomized placebo-controlled trial. Clin. Nutr. 2011, 30, 156–164. [Google Scholar] [CrossRef]
- Luoto, R.; Laitinen, K.; Nermes, M.; Isolauri, E. Impact of maternal probiotic-supplemented dietary counselling on pregnancy outcome and prenatal and postnatal growth: A double-blind, placebo-controlled study. Br. J. Nutr. 2010, 103, 1792–1799. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, K.; Kennelly, M.; Culliton, M.; Smith, T.; Maguire, O.C.; Shanahan, F.; Brennan, L.; McAuliffe, F.M. Probiotics in obese pregnancy do not reduce maternal fasting glucose: A double-blind, placebo-controlled, randomized trial (Probiotics in Pregnancy Study). Am. J. Clin. Nutr. 2014, 99, 1432–1439. [Google Scholar] [CrossRef]
- Asemi, Z.; Samimi, M.; Tabassi, Z.; Rad, M.N.; Foroushani, A.R.; Khorammian, H.; Esmaillzadeh, A. Effect of daily consumption of probiotic yoghurt on insulin resistance in pregnant women: A randomized controlled trial. Eur. J. Clin. Nutr. 2012, 67, 71–74. [Google Scholar] [CrossRef]
- Callaway, L.; McIntyre, H.D.; Barrett, H.L.; Foxcroft, K.; Tremellen, A.; Lingwood, B.E.; Tobin, J.M.; Wilkinson, S.A.; Kothari, A.; Morrison, M.; et al. Probiotics for the Prevention of Gestational Diabetes Mellitus in Overweight and Obese Women: Findings From the SPRING Double-blind Randomized Controlled Trial. Diabetes Care 2019, 42, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Videhult, F.K.; Andersson, Y.; Öhlund, I.; Stenlund, H.; Hernell, O.; West, C.E. Impact of probiotics during weaning on the metabolic and inflammatory profile: Follow-up at school age. Int. J. Food Sci. Nutr. 2015, 66, 686–691. [Google Scholar] [CrossRef]
- Videhult, F.K.; Öhlund, I.; Stenlund, H.; Hernell, O.; West, C.E. Probiotics during weaning: A follow-up study on effects on body composition and metabolic markers at school age. Eur. J. Nutr. 2014, 54, 355–363. [Google Scholar] [CrossRef]
- Luoto, R.; Kalliomäki, M.; Laitinen, K.; Isolauri, E. The impact of perinatal probiotic intervention on the development of overweight and obesity: Follow-up study from birth to 10 years. Int. J. Obes. 2010, 34, 1531–1537. [Google Scholar] [CrossRef] [Green Version]
- Ipar, N.; Aydogdu, S.D.; Yildirim, G.K.; Inal, M.; Gies, I.; Vandenplas, Y.; Dinleyici, E. Effects of synbiotic on anthropometry, lipid profile and oxidative stress in obese children. Benef. Microbes 2015, 6, 775–781. [Google Scholar] [CrossRef]
- Nagata, S.; Chiba, Y.; Wang, C.; Yamashiro, Y. The effects of the Lactobacillus casei strain on obesity in children: A pilot study. Benef. Microbes 2017, 8, 535–543. [Google Scholar] [CrossRef]
- Sanchis-Chordà, J.; Del Pulgar, E.M.G.; Carrasco-Luna, J.; Benítez-Páez, A.; Sanz, Y.; Codoñer-Franch, P. Bifidobacterium pseudocatenulatum CECT 7765 supplementation improves inflammatory status in insulin-resistant obese children. Eur. J. Nutr. 2018, 58, 2789–2800. [Google Scholar] [CrossRef] [PubMed]
- Safavi, S.M.; Farajian, S.; Kelishadi, R.; Mirlohi, M.; Hashemipour, M. The effects of synbiotic supplementation on some cardio-metabolic risk factors in overweight and obese children: A randomized triple-masked controlled trial. Int. J. Food Sci. Nutr. 2013, 64, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Kelishadi, R.; Farajian, S.; Safavi, M.; Mirlohi, M.; Hashemipour, M. A randomized triple-masked controlled trial on the effects of synbiotics on inflammation markers in overweight children. J. Pediatr. 2014, 90, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Vajro, P.; Mandato, C.; Licenziati, M.R.; Franzese, A.; Vitale, D.F.; Lenta, S.; Caropreso, M.; Vallone, G.; Meli, R. Effects of Lactobacillus rhamnosus Strain GG in Pediatric Obesity-related Liver Disease. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 740–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Famouri, F.; Shariat, Z.; Hashemipour, M.; Keikha, M.; Kelishadi, R. Effects of Probiotics on Nonalcoholic Fatty Liver Disease in Obese Children and Adolescents. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 413–417. [Google Scholar] [CrossRef]
- Alisi, A.; Bedogni, G.; Baviera, G.; Giorgio, V.; Porro, E.; Paris, C.; Giammaria, P.; Reali, L.; Anania, F.; Nobili, V. Randomised clinical trial: The beneficial effects of VSL#3 in obese children with non-alcoholic steatohepatitis. Aliment. Pharmacol. Ther. 2014, 39, 1276–1285. [Google Scholar] [CrossRef]
- Gøbel, R.J.; Larsen, N.; Jakobsen, M.; Mølgaard, C.; Michaelsen, K.F. Probiotics to Adolescents With Obesity. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 673–678. [Google Scholar] [CrossRef] [Green Version]
- Kadooka, Y.; Sato, M.; Imaizumi, K.; Ogawa, A.; Ikuyama, K.; Akai, Y.; Okano, M.; Kagoshima, M.; Tsuchida, T. Regulation of abdominal adiposity by probiotics (Lactobacillus gasseri SBT2055) in adults with obese tendencies in a randomized controlled trial. Eur. J. Clin. Nutr. 2010, 64, 636–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadooka, Y.; Sato, M.; Ogawa, A.; Miyoshi, M.; Uenishi, H.; Ogawa, H.; Ikuyama, K.; Kagoshima, M.; Tsuchida, T. Effect of Lactobacillus gasseri SBT2055 in fermented milk on abdominal adiposity in adults in a randomised controlled trial. Br. J. Nutr. 2013, 110, 1696–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S.-P.; Lee, K.-M.; Kang, J.-H.; Yun, S.-I.; Park, H.-O.; Moon, Y.; Kim, J.-Y. Effect ofLactobacillus gasseriBNR17 on Overweight and Obese Adults: A Randomized, Double-Blind Clinical Trial. Korean J. Fam. Med. 2013, 34, 80–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Yun, J.M.; Kim, M.K.; Kwon, O.; Cho, B. Lactobacillus gasseri BNR17 Supplementation Reduces the Visceral Fat Accumulation and Waist Circumference in Obese Adults: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Med. Food 2018, 21, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Anzawa, D.; Takami, K.; Ishizuka, A.; Mawatari, T.; Kamikado, K.; Sugimura, H.; Nishijima, T. Effect of Bifidobacterium animalis ssp. lactis GCL2505 on visceral fat accumulation in healthy Japanese adults: A randomized controlled trial. Biosci. Microbiota Food Health 2016, 35, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Omar, J.M.; Chan, Y.-M.; Jones, M.L.; Prakash, S.; Jones, P.J.H. Lactobacillus fermentum and Lactobacillus amylovorus as probiotics alter body adiposity and gut microflora in healthy persons. J. Funct. Foods 2013, 5, 116–123. [Google Scholar] [CrossRef]
- Kim, M.; Kim, M.; Kang, M.; Yoo, H.J.; Kim, M.S.; Ahn, Y.-T.; Sim, J.-H.; Jee, S.H.; Lee, J.H. Effects of weight loss using supplementation with Lactobacillus strains on body fat and medium-chain acylcarnitines in overweight individuals. Food Funct. 2017, 8, 250–261. [Google Scholar] [CrossRef]
- De Lorenzo, A.; Costacurta, M.; Merra, G.; Gualtieri, P.; Cioccoloni, G.; Marchetti, M.; Varvaras, D.; Docimo, R.; Di Renzo, L. Can psychobiotics intake modulate psychological profile and body composition of women affected by normal weight obese syndrome and obesity? A double blind randomized clinical trial. J. Transl. Med. 2017, 15, 135. [Google Scholar] [CrossRef] [Green Version]
- Szulińska, M.; Łoniewski, I.; Van Hemert, S.; Sobieska, M.; Bogdanski, P. Dose-Dependent Effects of Multispecies Probiotic Supplementation on the Lipopolysaccharide (LPS) Level and Cardiometabolic Profile in Obese Postmenopausal Women: A 12-Week Randomized Clinical Trial. Nutrients 2018, 10, 773. [Google Scholar] [CrossRef] [Green Version]
- Minami, J.-I.; Kondo, S.; Yanagisawa, N.; Odamaki, T.; Xiao, J.-Z.; Abe, F.; Nakajima, S.; Hamamoto, Y.; Saitoh, S.; Shimoda, T. Oral administration of Bifidobacterium breve B-3 modifies metabolic functions in adults with obese tendencies in a randomised controlled trial. J. Nutr. Sci. 2015, 4, e17. [Google Scholar] [CrossRef] [Green Version]
- Minami, J.; Iwabuchi, N.; Tanaka, M.; Yamauchi, K.; Xiao, J.-Z.; Abe, F.; Sakane, N. Effects of Bifidobacterium breve B-3 on body fat reductions in pre-obese adults: A randomized, double-blind, placebo-controlled trial. Biosci. Microbiota Food Health 2018, 37, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivey, K.L.; Hodgson, J.M.; A Kerr, D.; Lewis, J.R.; Thompson, P.L.; Prince, R.L. The effects of probiotic bacteria on glycaemic control in overweight men and women: A randomised controlled trial. Eur. J. Clin. Nutr. 2014, 68, 447–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, M.; Darimont, C.; Panahi, S.; Drapeau, V.; Marette, A.; Taylor, V.H.; Doré, J.; Tremblay, A. Effects of a Diet-Based Weight-Reducing Program with Probiotic Supplementation on Satiety Efficiency, Eating Behaviour Traits, and Psychosocial Behaviours in Obese Individuals. Nutrients 2017, 9, 284. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi-Sartang, M.; Bellissimo, N.; De Zepetnek, J.O.T.; Brett, N.R.; Mazloomi, S.M.; Fararouie, M.; Bedeltavana, A.; Famouri, M.; Mazloom, Z. The effect of daily fortified yogurt consumption on weight loss in adults with metabolic syndrome: A 10-week randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Zarrati, M.; Salehi, E.; Mofid, V.; Zadeh-Attar, M.J.H.; Nourijelyani, K.; Bidad, K.; Shidfar, F. Relationship between probiotic consumption and IL-10 and IL-17 secreted by PBMCs in overweight and obese people. Iran. J. Allergy Asthma Immunol. 2013, 12, 404–406. [Google Scholar]
- Gomes, A.C.; De Sousa, R.G.M.; Botelho, P.B.; Gomes, T.L.N.; Prada, P.O.; Mota, J.F. The additional effects of a probiotic mix on abdominal adiposity and antioxidant Status: A double-blind, randomized trial. Obesity 2016, 25, 30–38. [Google Scholar] [CrossRef]
- Stenman, L.K.; Lehtinen, M.J.; Meland, N.; Christensen, J.E.; Yeung, N.; Saarinen, M.T.; Courtney, M.; Burcelin, R.; Lähdeaho, M.-L.; Linros, J.; et al. Probiotic With or Without Fiber Controls Body Fat Mass, Associated With Serum Zonulin, in Overweight and Obese Adults—Randomized Controlled Trial. EBioMedicine 2016, 13, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Karbaschian, Z.; Mokhtari, Z.; Pazouki, A.; Kabir, A.; Hedayati, M.; Moghadam, S.S.; Mirmiran, P.; Hekmatdoost, A. Probiotic Supplementation in Morbid Obese Patients Undergoing One Anastomosis Gastric Bypass-Mini Gastric Bypass (OAGB-MGB) Surgery: A Randomized, Double-Blind, Placebo-Controlled, Clinical Trial. Obes. Surg. 2018, 28, 2874–2885. [Google Scholar] [CrossRef]
- Aller, R.; De Luis, D.A.; Izaola, O.; Conde, R.; Sagrado, M.G.; Primo, D.; De Fuente, B.L.; Gonzalez, J. Effect of a probiotic on liver aminotransferases in nonalcoholic fatty liver disease patients: A double blind randomized clinical trial. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 1090–1095. [Google Scholar]
- Miccheli, A.; Capuani, G.; Marini, F.; Tomassini, A.; Praticò, G.; Ceccarelli, S.; Gnani, D.; Baviera, G.; Alisi, A.; Putignani, L.; et al. Urinary 1H-NMR-based metabolic profiling of children with NAFLD undergoing VSL#3 treatment. Int. J. Obes. 2015, 39, 1118–1125. [Google Scholar] [CrossRef]
- Javadi, L.; Ghavami, M.; Khoshbaten, M.; Safaiyan, A.; Barzegari, A.; Gargari, B.P. The Effect of Probiotic and/or Prebiotic on Liver Function Tests in Patients with Nonalcoholic Fatty Liver Disease: A Double Blind Randomized Clinical Trial. Iran. Red Crescent Med. J. 2017, 19, e46017. [Google Scholar] [CrossRef]
- Razmpoosh, E.; Javadi, A.; Ejtahed, H.S.; Mirmiran, P.; Javadi, M.; Yousefinejad, A. The effect of probiotic supplementation on glycemic control and lipid profile in patients with type 2 diabetes: A randomized placebo controlled trial. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Madjd, A.; A Taylor, M.; Mousavi, N.; Delavari, A.; Malekzadeh, R.; Macdonald, I.A.; Farshchi, H.R. Comparison of the effect of daily consumption of probiotic compared with low-fat conventional yogurt on weight loss in healthy obese women following an energy-restricted diet: A randomized controlled trial1. Am. J. Clin. Nutr. 2015, 103, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharafetdinov, K.K.; Plotnikova, O.A.; Alexeeva, R.I.; Sentsova, T.B.; Songisepp, E.; Štšepetova, J.; Smidt, I.; Mikelsaar, M. Hypocaloric diet supplemented with probiotic cheese improves body mass index and blood pressure indices of obese hypertensive patients-a randomized double-blind placebo-controlled pilot study. Nutr. J. 2013, 12, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leber, B.; Tripolt, N.J.; Blattl, D.; Eder, M.; Wascher, T.C.; Pieber, T.R.; Stauber, R.; Sourij, H.; Oettl, K.; Stadlbauer, V. The influence of probiotic supplementation on gut permeability in patients with metabolic syndrome: An open label, randomized pilot study. Eur. J. Clin. Nutr. 2012, 66, 1110–1115. [Google Scholar] [CrossRef] [Green Version]
- Mobini, R.; Tremaroli, V.; Ståhlman, M.; Karlsson, F.; Levin, M.; Ljungberg, M.; Sohlin, M.; Forslund, H.B.; Perkins, R.; Bäckhed, F.; et al. Metabolic effects ofLactobacillus reuteriDSM 17938 in people with type 2 diabetes: A randomized controlled trial. Diabetes Obes. Metab. 2017, 19, 579–589. [Google Scholar] [CrossRef]
- Mahadzir, M.D.A.; Shyam, S.; Barua, A.; Krishnappa, P.; Ramamurthy, S. Effect of Probiotic Microbial Cell Preparation (MCP) on Fasting Blood Glucose, Body Weight, Waist Circumference, and faecal short chain fatty acids among overweight Malaysian adults: A pilot randomised controlled trial of 4 weeks. Malays. J. Nutr. 2017, 23, 329–341. [Google Scholar]
- Osterberg, K.L.; Boutagy, N.E.; McMillan, R.P.; Stevens, J.R.; Frisard, M.I.; Kavanaugh, J.W.; Davy, B.M.; Davy, K.P.; Hulver, M.W. Probiotic supplementation attenuates increases in body mass and fat mass during high-fat diet in healthy young adults. Obesity 2015, 23, 2364–2370. [Google Scholar] [CrossRef]
- Pedret, A.; Valls, R.M.; Calderón-Pérez, L.; Llauradó, E.; Companys, J.; Pla-Pagà, L.; Moragas, A.; Martín-Luján, F.; Ortega, Y.; Giralt, M.; et al. Effects of daily consumption of the probiotic Bifidobacterium animalis subsp. lactis CECT 8145 on anthropometric adiposity biomarkers in abdominally obese subjects: A randomized controlled trial. Int. J. Obes. 2018, 43, 1863–1868. [Google Scholar] [CrossRef] [Green Version]
- Higashikawa, F.; Noda, M.; Awaya, T.; Danshiitsoodol, N.; Matoba, Y.; Kumagai, T.; Sugiyama, M. Antiobesity effect of Pediococcus pentosaceus LP28 on overweight subjects: A randomized, double-blind, placebo-controlled clinical trial. Eur. J. Clin. Nutr. 2016, 70, 582–587. [Google Scholar] [CrossRef]
- Jones, R.B.; Alderete, T.L.; A Martin, A.; A Geary, B.; Hwang, D.H.; Palmer, S.; Goran, M.I. Probiotic supplementation increases obesity with no detectable effects on liver fat or gut microbiota in obese Hispanic adolescents: A 16-week, randomized, placebo-controlled trial. Pediatr. Obes. 2018, 13, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Halkjær, S.I.; Nilas, L.; Carlsen, E.M.; Cortes, D.; Halldórsson, T.I.; Olsen, S.F.; Pedersen, A.E.; Krogfelt, K.A.; Petersen, A.M. Effects of probiotics (Vivomixx®) in obese pregnant women and their newborn: Study protocol for a randomized controlled trial. Trials 2016, 17, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaguarnera, M.; Vacante, M.; Antic, T.; Giordano, M.; Chisari, G.; Acquaviva, R.; Mastrojeni, S.; Malaguarnera, G.; Mistretta, A.; Volti, G.L.; et al. Bifidobacterium longum with Fructo-Oligosaccharides in Patients with Non Alcoholic Steatohepatitis. Dig. Dis. Sci. 2011, 57, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, H.; Mahmood, N.; Kumar, M.; Varikuti, S.R.; Challa, H.R.; Myakala, S.P. Effect of Probiotic (VSL#3) and Omega-3 on Lipid Profile, Insulin Sensitivity, Inflammatory Markers, and Gut Colonization in Overweight Adults: A Randomized, Controlled Trial. Mediat. Inflamm. 2014, 2014, 348959. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Bae, J.-H. Probiotics for weight loss: A systematic review and meta-analysis. Nutr. Res. 2015, 35, 566–575. [Google Scholar] [CrossRef]
- Mohammadi, H.; Ghavami, A.; Hadi, A.; Askari, G.; Symonds, M.; Miraghajani, M. Effects of pro-/synbiotic supplementation on anthropometric and metabolic indices in overweight or obese children and adolescents: A systematic review and meta-analysis. Complement. Ther. Med. 2019, 44, 269–276. [Google Scholar] [CrossRef]
- Borgeraas, H.; Johnson, L.K.; Skattebu, J.; Hertel, J.K.; Hjelmesaeth, J.; Hjelmesæth, J. Effects of probiotics on body weight, body mass index, fat mass and fat percentage in subjects with overweight or obesity: A systematic review and meta-analysis of randomized controlled trials. Obes. Rev. 2017, 19, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wu, Y.; Fei, X. Efect of probiotics on body weight and body-mass index: A systematic review and meta-analysis of randomized, controlled trials. Int. J. Food Sci. Nutr. 2016, 67, 571–580. [Google Scholar] [CrossRef]
- Koutnikova, H.; Genser, B.; Monteiro-Sepulveda, M.; Faurie, J.-M.; Rizkalla, S.; Schrezenmeir, J.; Clément, K. Impact of bacterial probiotics on obesity, diabetes and non-alcoholic fatty liver disease related variables: A systematic review and meta-analysis of randomised controlled trials. BMJ Open 2019, 9, e017995. [Google Scholar] [CrossRef]
- Dror, T.; Dickstein, Y.; Dubourg, G.; Paul, M. Microbiota manipulation for weight change. Microb. Pathog. 2017, 106, 146–161. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Short-Chain Fatty Acid (SCFA) | Receptors | Properties |
|---|---|---|
| Acetic acid | GPR 41, GPR43 | Stimulates the secretion of intestinal hormones PYY and GLP-1—appetite reduction Increased production of leptin Precursors for cholesterol and fat acids synthesis |
| Propionic acid | GPR 41, GPR43 | Stimulates the secretion of intestinal hormones PYY and GLP-1—appetite reduction Inhibition of fat accumulation in adipocytes Increased production of leptin Substrate of gluconeogenesis |
| Butyric acid | GPR 41, GPR43, GPR109A | Differentiation of Treg-cell Decrease the production of IL-12 Increase the production of IL-10 Stimulation of fat storage in adipose tissue Promotes intestinal epithelial integrity Increase in the oxidation of fatty acids Stimulating production of mucin |
| Author | Study Design | Duration of Intervention | Intervention | Sample (Age) | Control/Placebo Groups | Clinical Outcome (vs. Control/Placebo Group) | Reference |
|---|---|---|---|---|---|---|---|
| Ilmonen et al. | Randomized placebo-controlled trial | From the first trimester, pregnancy up to 6 months after delivery | L. rhamnosus GG, B. lactis ( CFU) 1 capsule per day | 256 pregnant women; 85 diet/probiotics; 86 diet/placebo; 85 control/placebo (25–35 years) | 1 capsule containing microcrystalline cellulose and dextrose per day/without diet | ↓WC (p < 0.001) | [75] |
| Luoto et al. | Randomized, double-blind, placebo-controlled trial | From the first trimester to the end of breastfeeding | L. rhamnosus GG, B. lactis ( CFU) 1 capsule per day | 256 pregnant women; 85 diet/probiotics; 86; diet/placebo; 85 control/placebo | 1 capsule containing microcrystalline cellulose and dextrose per day/without diet | Reduction in gestational diabetes (GDM) (13% diet/probiotics; 36% diet/placebo; 34% control/placebo) (p < 0.003) | [76] |
| Lindsay et al. | Randomized, double-blind, placebo-controlled trial | 4 weeks | L. salivarius UCC118 ( CFU) 1 capsule per day | 138 women in the first trimester of pregnancy; 63 probiotic group; 75 placebo group (>18 years) | 1 capsule per day | No effect | [77] |
| Asemi et al. | Randomized single-blinded controlled trial | 9 weeks | 200 g of yogurt containing L. acidophilus LA5 and B. animalis BB12 ( CFU/g) per day | 67 women in the third trimester of pregnancy; 33 probiotic group; 37 placebo group (18–30 years) | 200 g of conventional yoghurt per day | No effect | [78] |
| Callaway et al. | Prospective double-blind RCT | From 20 weeks gestation to delivery | L. rhamnosus, B. animalis subspecies lactis ( CFU) per day | 211 obese and overweight women; 207 probiotic; 204 placebo (>18 years) | 1 capsule containing microcrystalline cellulose and dextrose per day | No effect | [79] |
| Karlsson Videhult et al. | Randomized, double-blind, placebo-controlled trial | From 4 to 13 months of age | Cereals containing of L. paracasei ssp. paracasei—LF19 ( CFU) per day | 120 children; 58 probiotic group; 62 placebo group (8–9 years) | Cereals without LF19 | Effect on anthropometric parameters (p < 0.05) | [80] |
| Karlsson Videhult et al. | Randomized, double-blind, placebo-controlled trial | From 4 to 13 months of age | Cereals containing of L. paracasei ssp. paracasei—LF19 ( CFU) per day | 120 children; 58 intervention group; 62 placebo group (8–9 years) | Cereals without LF19 | No effect | [81] |
| Luoto et al. | Randomized, double-blind, prospective follow-up | Mothers 4 weeks before delivery; infants for 6 months after birth | L. rhamnosus GG ( CFU) 1 capsule per day | 113 Mother-child; 54 probiotic group; 59 placebo group | 1 capsule containing microcrystalline cellulose per day | Probiotic administration may reduce weight gain, especially up to 4 years of age (p = 0.08) | [82] |
| Ipar et al. | Open-label, randomized, controlled trial | 4 weeks | L. acidophilus, L. rhamnosus, B. bifidum, B. longum (each 4.3 × CFU), E. faecium (8.2 × CFU), FOS 625mg, lactulose 400 mg, vit. A 6 mg, B1 1.8 mg, B2 1.6 mg, B6 2.4 mg, E 30 mg, C 75 mg 1 sachet per day | 117 children with primary obesity; 77 intervention group + calorie reduction + physical activity; 40 calorie reduction + physical activity (5–17 years) | Calorie reduction + physical activity | ↓BMI (p < 0.05); ↓ HC and WC (p < 0.05) | [83] |
| Nagata et al. | Open-label prospective | 6 months | 1 bottle of fermented milk contained L. casei Shirota (4 × ) per day | 12 obese children; 22 healthy non-obese children | Diet + physical activity | ↓ weight (p < 0.05) | [84] |
| Sanchis-Chorda et al. | Open-label prospective | 13 weeks | B. pseudocatenulatum CECT 7765 ( CFU) 1 capsule per day | 48 obese children with insulin resistance; 24 probiotic + diet; 24 diet (10–15 years) | 1 capsule per day | Improved lipid profile (p = 0.035), inflammatory markers (p = 0.026) and ↓ BMI (p = 0.001) | [85] |
| Safavi et al. | Randomized triple-blinded controlled trial | 8 weeks | L. casei, L. rhamnosus, S. thermophilus, B. breve, L. acidophilus, B. longum and L. bulgaricus (Each 2 × CFU), vit. E, A, C per day | 56 obese children and adolescents; 29 intervention group; 27 placebo (6–18 years) | 1 capsule containing maltodextrin per day | ↓ BMI (p = 0.002), ↓ WC (p = 0.001) | [86] |
| Kelishadi et al. | Triple-blinded controlled RTC | 8 weeks | L. casei, L. rhamnosus, S. thermophilus, B. breve, L. acidophilus, B. longum and L. bulgaricus (Each 2 × CFU), vit. E, A, C per day | 56 obese children and adolescents; 29 probiotic; 27 placebo (6–18 years) | 1 capsule containing maltodextrin per day | ↓ BMI (p = 0.002) | [87] |
| Vajro et al. | Double-blind placebo-controlled pilot study | 8 weeks | L. rhamnosus GG (12 billion CFU/day) | 20 obese children with persisting hypertransaminasemia and ultrasonographic bright liver; 10 probiotic; 10 placebo (10–13 years) | Placebo | No effect | [88] |
| Famouri et al. | Randomized triple-blind placebo-controlled trial | 12 weeks | L. acidophilus ATCC B3208 (3 × CFU), B. lactis DSMZ 32269 (6 × CFU), B. bifidum ATCC SD6576, L. rhamnosus DSMZ 21690 (each 2 × CFU) 1 capsule per day | 64 obese children and adults with NAFLD; 32 probiotics group; 32 placebo group (10–18 years) | 1 capsule per day | Improvement of lipid profile (p < 0.001) and NAFLD (p < 0.05); ↓ plasma liver enzymes (p = 0.02); No effect on BMI and body weight | [89] |
| Alisi et al. | Parallel double-blind RCT | 4 months | Age < 10 year 1 sachet per day of VSL#3, age > 10 year 2 sachets of VSL#3 | 44 children with NAFLD; 22 probiotic group; 22 placebo group (9–12 years) | Age < 10 year 1 sachet per day, age > 10 year 2 sachets of VSL#3 | ↓ BMI (p = 0.001); ↑ GLP-1 (p = 0.001) | [90] |
| Gobel et al. | Randomized double-blind placebo-controlled trial | 12 weeks | L. salivarius LS 33 ( CFU) 1 capsule per day | 50 obese adolescents; 27 probiotic; 23 placebo (12–15 years) | 1 capsule containing cellulose, silicon dioxide and rice-maltodextrin per day | No effect | [91] |
| Kadooka et al. | Multi-centre, double-blind, parallel group RCT | 12 weeks | 200 g of fermented milk containing L. gasseri SBT2055 ( CFU/g) per day) | 87 obese adults; 43 probiotic group, 44 placebo group (33–63 years) | 200 g of fermented milk without probiotic per day | ↓ Weight, BMI, HC and WC (p = 0.01) | [92] |
| Kadooka et al. | Multi-centre, double-blind, parallel group RCT | 12 weeks | 200 g of fermented milk containing L. gasseri SBT2055 ( or CFU/g) per day | 210 obese adults; 69 high dose probiotic, 71 low dose probiotic, 70 placebo (35–60 years) | 200 g of fermented milk without probiotic per day | ↓BMI, HC, WC and body fat mass (p = 0.01) | [93] |
| Jung et al. | Randomized, double-blind, placebo-controlled trial | 12 weeks | L. gasseri BNR17 ( CFU) 6 capsules per day | 57 overweight or obese adults; 28 probiotic group; 29 placebo group (19–60 years) | 6 capsules containing trehalose, skim milk, and fructooligosaccharide per day | ↓ WC and HC (p = 0.015); ↓ body weight (p = 0.008) | [94] |
| Kim et al. | Randomized, double-blind, placebo-controlled trial | 12 weeks | L. gasseri BNR17 ( CFU) 2 capsuls per day | 90 overweight or obese adults; 30 high dose probiotic; 30 low dose probiotic; 30 placebo (20–75 years) | 2 capsule containing maltodextrin, crystalline cellulose, and magnesium stearate per day | High and low probiotic doses reduce body weight and waist circumferences (p < 0.05); A high dose of probiotic reduces visceral adipose tissue (p < 0.05) | [95] |
| Takahashi et al. | Multicenter, randomized, double-blind, placebo-controlled intervention trial | 12 weeks | 100g of fermented milk containing B. animalis ssp. lactis GCL2505 (8 × CFU) per day | 137 overweight and obese adults; 69 probiotic; 68 placebo (20–65 years) | 100 g of fermented milk without probiotic | ↓ visceral fat (p = 0.05) | [96] |
| Omar et al. | Randomized double-blind placebo-controlled trial | 43 days | 100 g of yogurt containing L. amylovorus (1.39 × CFU) or 100 g of yogurt containing L. fermentum (1.08 × CFU) per day | 28 overweight and obese adults; 14 probiotic LA; 12 probiotic LF; 12 placebo (16–60 years) | 100 g of control yogurt per day | No effect | [97] |
| Kim et al. | Randomized, double-blind, placebo-controlled trial | 12 weeks | 2 g of powder containing L. plantarum KY1032 and L. curvatus (2.5 × CFU) twice a day | 65 overweight adults; 32 probiotic; 34 placebo (25–75 years) | 2 g of powder without probiotic twice a day | ↓ body weight (p < 0.05) | [98] |
| De Lorenzo et al. | Randomized, double-blinded controlled trial | 3 weeks | S. thermophilus SGSt01, B. animalis subsp. Lactis SGB06, S. thermophiles, B. bifidum SGB02, L. delbrueckii spp. Bulgaricus DSM 20081, L. lactis subsp. Lactis SGLc01, L. acidophilus SGL11, L. plantarum SGL07, L. reuteri SGL01 (1.5 × CFU) 1 sachet per day | 48 women with normal weight, overweight or obesity; 24 probiotics; 24 placebo (20–65 years) | 1 sachet containing maltodextrin from corn and silica per day | ↓ BMI, and fat mass (p < 0.05) | [99] |
| Szulińska et al. | Randomized, double-blind, placebo-controlled trial | 12 weeks | 2 g of powder containing B. bifidum W23, B. lactis W51, B. lactis W52, L. acidophilus W37, L. brevis W63, L. casei W56, L. salivarius W24, L. lactis W19, and L. lactis W58 (High dose CFU or low dose 2.5 × CFU per day divided in two equal doses) | 71 obese women; 23 high dose probiotic; 24 low dose probiotic; 24 placebo (45–70 years) | 2 g of powder containing only the excipients, i.e., maize starch and maltodextrins | Improving lipid profile and glucose metabolism (p < 0.05); ↓ visceral fat, waist circumference (p < 0.05); High probiotic doses ↓ LPS levels (p < 0.05) | [100] |
| Minami et al. | Randomized, double-blind, parallel-group comparative trial | 12 weeks | B. breve B-3 (5 × CFU, 3 capsules per day) | 44 overweight adults with diabetes; 19 probiotic group; 25 placebo group (40–69 years) | 3 capsules per day containing maize starch | Improving liver function (p < 0.05); Reduction of metabolic disorders (p < 0.05); Reduction of fat mass (p < 0.05) | [101] |
| Minami et al. | Randomized, double-blind, placebo-controlled trial | 12 weeks | B. breve B-3 (2 × CFU) 2 capsuls per day | 80 healthy pre-obese adults; 40 probiotics; 40 placebo (20–64 years) | 2 capsules per day containing corn starch | ↓ body fat (p < 0.05); No effect on lipid profile | [102] |
| Ivey et al. | Randomized double-blinded parallel study | 6 weeks | Both the probiotic yogurt and the probiotic capsule contained L. acidophilus La5, B. animalis subsp. lactis Bb12 (3 × CFU per day) | 156 overweight men and women; 40 probiotic yoghurt + probiotic capsules; 37 probiotic yoghurt + placebo capsules; 39 control milk + probiotic capsules; 40 control milk + placebo capsules (over 55 years) | Control milk | No effect | [103] |
| Sanchez et al. | Randomized, double-blind, placebo-controlled trial | 12 weeks | L. rhamnosus CGMCC1.3724 (3.24 × CFU) + 90 g inulin and 210 g FOS, 2 capsules per day | 125 obese adults; 62 intervention group, 63 placebo group (18-55 years) | 2 capsules containing maltodextrin and magnesium stearate per day | ↓ weight (p = 0.02); ↓ appetite (p = 0.02) | [104] |
| Mohammadi-Sartang et al. | Randomized double-blinded controlled trial | 10 weeks | 250 g of fortified yogurt (FSY) with whey protein, calcium, vit. D, fiber and probiotic (S. thermophiles, L. bulgaricus and at least B. lactis Bb-12) 250 g twice a day | 87 obese adults with metabolic syndrome; 44 fortified yogurt + diet; 43 low-fat conventional yogurt + diet (20–65 years) | 250 g of low fat plain yogurt with St. thermophiles and L. bulgaricus (LFY) twice a day | ↓ body fat mass (p < 0.05); ↓ WC (p < 0.05), ↓ TG ↓ HOMAR-IR value (p < 0.05); ↑HDL (p < 0.05) | [105] |
| Zarrati et al. | Randomized double-blind placebo-controlled trial | 8 weeks | 200 g of yogurt containing L. acidophilus La5, B. BB12 and L. casei DN001 CFU/g | 75 obese adults; 25 regular yogurt + low calorie diet; 25 probiotic yogurt + low calorie diet; 25 probiotic yogurt without low calorie diet | 200 g of yogurt without probiotics | ↓ body mass and inflammation (p < 0.001) | [106] |
| Gomes et al. | Randomized, double-blind, placebo-controlled trial | 8 weeks | L. acidophilus LA-14, L. casei LC-11, L. lactis LL-23, B. bifidum BB-06, B. lactis BL-4 (each CFU) 4 sachets per day + diet | 43 overweight or obese women; 21 probiotics; 22 placebo (20–59 years) | 4 sachets per day of placebo + diet | ↓ abdominal adiposity (p = 0.02) | [107] |
| Stenman et al. | Randomized, double-blind, placebo-controlled, multi-center clinical trial | 6 months | B. animalis ssp. lactis 420 ( CFU) with or without 12g of LU per day | 134 overweight or obese adults; 25 probiotic; 36 LU (12 g); 37 probiotic + LU (12 g); 36 placebo (18–65 years) | 1 sachet (12 g) containing microcrystalline cellulose per day | ↓ fat mass and WC (p = 0.02) | [108] |
| Karbaschian et al. | Randomized, double-blind, placebo-controlled trial | 4 weeks before surgery and 12 weeks after surgery | L. casei (3.5 × CFU/g), L. rhamnosus (7.5 × CFU/g), S. thermophilus ( CFU/g), B. breve ( CFU/g), L. acidophilus ( CFU/g), B. longum (3.5 × CFU/g), and L. bulgaricus ( CFU/g) and 38.5 mg FOS 1 capsule per day | 46 Obese women; 23 probiotics; 23 placebo (18–60 years) | 1 capsule containing maltodextrin per day | ↓ body weight (p < 0.05) | [109] |
| Aller et al. | Randomized, double-blind, parallel-group comparative trial | 3 months | L. bulgaricus and St. thermophilus (500 million of probiotic, 1 tablet per day) | 28 adults with NAFLD; 14 probiotic group; 14 placebo group (39–60 years) | 1 tablet contained 120 mg of starch | Improving glucose metabolism (p < 0.05); Improvement of liver function (p < 0.05); No effect on body weight | [110] |
| Miccheli et al. | Parallel-arm, double-blind RCT | 4 months | (112.5 × CFU) - VSL#3 (age < 10 year 1 sachet per day of VSL#3; age > 10 year 2 sachets of VSL#3) + diet + physical activity | 31 children with NAFLD; 15 probiotic group; 16 placebo (9–12 years) | 1 sachet placebo | ↓ BMI (p = 0.045) and liver function (p = 0.026) | [111] |
| Javadi et al. | Randomized double-blind placebo-controlled trial | 3 months | B. longum, L. acidophilus (each 2 × CFU per day) | 75 obese adults: 20 probiotic; 19 prebiotic; 17 probiotic + prebiotic; 19 placebo (20–60 years) | Supplementation with probiotic and/or prebiotic improves liver function (p < 0.05); ↓ body mass and BMI (p < 0.05) | [112] | |
| Razmpoosh | Randomized, double-blind, placebo-controlled trial | 6 weeks | L. acidophilus (2 × CFU), L. casei, B. longum (each 7 × CFU), L. rhamnosus (1.5 × CFU), L. bulgaricus (2 × CFU), B. breve (3 × CFU), S. thermophilus (1.5 × CFU), 100 mg FOS 2 capsules per day | 60 adults with diabetes; 30 probiotic group, 30 placebo group (30–75 years) | 2 capsules per day containing fructooligosaccharide and magnesium stearate | ↓ fasting glucose (p = 0.001); ↑ HDL (p = 0.002); No effect on anthropometric parameters and insulin levels | [113] |
| Madjd et al. | Randomized, single-blind, controlled trial | 12 weeks | 200 g of probiotic yogurt (S. thermophiles, L. bulgaricus, L. acidophilus LA5, B. lactis BB12 - CFU) twice a day | 89 healthy overweight and obese women; 45 probiotic; 44 placebo (18–50 years) | 200 g low-fat yogurt twice a day | No effect | [114] |
| Sharafedtinov et al. | Randomized, double-blind, placebo-controlled trial | 3 weeks | 50 g cheese with L. plantarum TENSIA per day | 40 obese adults with hypertension; 25 probiotic group; 15 placebo group (30–69 years) | 50 g cheese without probiotic per day | ↓ BMI and morning blood pressure (p = 0.001) | [115] |
| Leber et al. | Open label, randomized pilot study | 3 months | 1 bottles (65 mL) containing L. casei Shirota (6.5 × CFU) three times a day | 28 adults with MetS; 13 probiotic; 15 placebo (24–66 years) | 1 bottles (65 mL) of placebo three times a day | No effect | [116] |
| Mobini et al. | Randomized double-blind placebo-controlled trial | 12 weeks | Powder containing L. reuteri DSM 17938 low dose ( CFU) or high dose ( CFU) | 44 patients with type 2 diabetes; 15 low dose; 14 high dose; 15 placebo (50–75 years) | Powder without probiotic | No effect | [117] |
| Mahadzir et al. | Randomized triple-blind placebo-controlled trial | 4 weeks | L. acidophilus BCMC 12130, L. casei subsp. BCMC 12313, L. lactis BCMC 12451, B. bifidum BCMC 02290, B. longum BCMC 02120 and B. infantis BCMC 02129 (3 × CFU per day) twice a day | 24 overweight adults; 12 probiotics; 12 placebo (18–50 years) | 2 sachets without probiotic | No effect | [118] |
| Osterberg et al. | Randomized, double-blind, parallel-group comparative trial | 4 weeks | VLS#3 (4.5 × ) 2 sachets per day | 20 healthy non-obese; 9 probiotic; 11 placebo group (18–30 years) | 2 sachets per day | ↓ body mass (p = 0.023) | [119] |
| Pedret et al. | Randomized, parallel, double-blind, placebo-controlled trial | 12 weeks | B. animalis subsp. lactis CECT 8145 ( CFU) 1 capsul per day; heat-killed B. animalis subsp. lactis CECT 8145 ( CFU) 1 capsule per day | 126 abdominally obese adults, 42 probiotic; 44 heat killed from probiotic; 40 placebo group (>18 years) | 1 capsule containing 300 mg of maltodextrose | ↓ BMI and WC (p < 0.05) | [120] |
| Higashikawa et al. | Randomized double-blind placebo-controlled trial | 12 weeks | Powder containing living Pediococcus pentosaceus LP28 ( CFU) with dextrin per day; Powder containing heat-kill Pediococcus pentosaceus LP28 ( CFU) with dextrin per day | 62 overweight adults; 21 living LP28; 21 heat-killed LP28; 20 placebo group (20–70 years) | Powder containing dextrin | ↓ body mass (p = 0.004), BMI (p = 0.035), WC (p = 0.009) in group after Heat-killed LP28 | [121] |
| Jones et al. | Randomized double-blind placebo-controlled trial | 16 weeks | VLS#3 3 sachets per day | 19 obese adolescents; 8 probiotic; 11 placebo group (12–18 years) | 3 sachets per day | ↑ total adiposity (p = 0.01) | [122] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiciński, M.; Gębalski, J.; Gołębiewski, J.; Malinowski, B. Probiotics for the Treatment of Overweight and Obesity in Humans—A Review of Clinical Trials. Microorganisms 2020, 8, 1148. https://doi.org/10.3390/microorganisms8081148
Wiciński M, Gębalski J, Gołębiewski J, Malinowski B. Probiotics for the Treatment of Overweight and Obesity in Humans—A Review of Clinical Trials. Microorganisms. 2020; 8(8):1148. https://doi.org/10.3390/microorganisms8081148
Chicago/Turabian StyleWiciński, Michał, Jakub Gębalski, Jakub Gołębiewski, and Bartosz Malinowski. 2020. "Probiotics for the Treatment of Overweight and Obesity in Humans—A Review of Clinical Trials" Microorganisms 8, no. 8: 1148. https://doi.org/10.3390/microorganisms8081148
APA StyleWiciński, M., Gębalski, J., Gołębiewski, J., & Malinowski, B. (2020). Probiotics for the Treatment of Overweight and Obesity in Humans—A Review of Clinical Trials. Microorganisms, 8(8), 1148. https://doi.org/10.3390/microorganisms8081148