Abstract
Non-typhoidal Salmonella serovars are worldwide spread foodborne pathogens that cause diarrhea in humans and animals. Colonization of gnotobiotic piglet intestine with porcine indigenous mucinolytic Bifidobacterium boum RP36 strain and non-mucinolytic strain RP37 and their interference with Salmonella Typhimurium infection were compared. Bacterial interferences and impact on the host were evaluated by clinical signs of salmonellosis, bacterial translocation, goblet cell count, mRNA expression of mucin 2, villin, claudin-1, claudin-2, and occludin in the ileum and colon, and plasmatic levels of inflammatory cytokines IL-8, TNF-α, and IL-10. Both bifidobacterial strains colonized the intestine comparably. Neither RP36 nor RP37 B. boum strains effectively suppressed signs of salmonellosis. Both B. boum strains suppressed the growth of S. Typhimurium in the ileum and colon. The mucinolytic RP36 strain increased the translocation of S. Typhimurium into the blood, liver, and spleen.
Keywords:
Bifidobacterium boum; Salmonella Typhimurium; goblet cells; mucin; mucinolytic; germ-free; gnotobiotic; piglet 1. Introduction
The coevolution of the host and its microbiota developed a mutually-beneficial relationship between them, and the microbial community became indispensable for the host’s metabolism and health [1]. The complex body microbiota consists of viruses [2], bacteria [3], archeae [4], fungi and yeasts [5], and protozoa [6]. Relationships between hosts and their microbiota can range from both-side convenient mutualism to host-destructive parasitism [7].
Balanced microbiota is an essential prerequisite for the host’s health, and its majority is located in the intestine [8,9]. The newborn intestine is colonized depending on the way of delivery, nutrition, and use of antibiotics [10,11]. Initially, the intestinal tract of vaginally delivered newborn colonize facultative anaerobes and create suitable conditions, e.g., lower pH and consumption of oxygen, for the subsequent onset of strict anaerobic species, e.g., Bacteroides, Bifidobacterium, and Clostridium spp. [10]. Breastfeeding refines the microbiota by additional microbes, human milk oligosaccharides (HMO), and immunological proteins [12], and the microbiome of breastfed infants consists of higher Proteobacteria and lower Bacteroidetes and Firmicutes than those formula-fed [13]. Bifidobacteria are the most abundant enteric bacteria in the vaginally born infant intestine [14,15]. They confer their beneficial effect by the saccharolytic activity toward glycans abundant in the infant intestine [16]. Moreover, they suppress pathogenic microorganisms [14,17,18,19]. In contrast, infants born through Cesarean section are mainly colonized with skin and hospital environment microbes, show lower numbers of bifidobacteria, and are more often colonized with Clostridium difficile [20]. Exclusively formula-fed infants are frequently colonized with Escherichia coli, C. difficile, Bacteroides, and Lactobacillus spp., compared with breastfed infants [21]. Early life represents an opportunity to shape the intestinal microbial ecosystem [12]. Adult-like microbiota forms in children at the age of 3–5 years and is individually characteristic and relatively stable for adult life [22]. The composition of the body microbiota varies in dependence on the host’s body surface and is highly personalized [23]. The most abundant bacteria are in the lower part of the intestinal tract that comprises in adults around 2000 species of predominantly anaerobic bacteria [24,25]. According to recently corrected counts, the total number of bacterial cells is comparable with the number of the host body cells [26].
A single layer of intestinal epithelial cells forms an interface between the intestinal lumen with bacteria and the host body. This interface must tolerate the luminal microbiota and their products and simultaneously protect the host against potentially harmful dietary antigens and invading pathogens [27]. The apical parts of the adjacent enterocytes of the epithelial layer are joined by tight junction proteins as claudins, occludin, and zonula occludens proteins and create a semipermeable layer [28]. This layer is covered by high molecular weight glycoprotein mucins that produce specialized enterocytes called goblet cells [27]. Mucins, as the main component of the mucus layer, protect the intestinal epithelium from infectious and non-infectious agents [29]. Mucin 2 (MUC2) is the predominant mucin in the human colon [30].
Much translational knowledge is obtained from experimental work with animal models that are mainly laboratory rodents in contrast that they are very dissimilar to humans in the transcriptome [31], microbiome composition [32,33], and duration of sepsis [34]. Humans and pigs share related anatomy, physiology, and genetics that predetermine the pig as a suitable animal model in human gastroenterology [35], infections [36], and sepsis [37]. Additionally, a similar composition of the porcine intestinal microbiome [33,38] incites the use of the pig in host-microbiota cross-talk translation research [35].
Gnotobiotic piglets proved their suitability for transplantation of the human microbiota [39,40]. Generally, the gnotobiotic animals are germ-free or animals associated with simple defined microbiota [41,42,43]. A crucial role for the host’s colonization resistance and its health plays the composition of the microbiota. Germ-free mice colonized with intestinal microbiota from the adult mice showed higher resistance to infection with Salmonella Typhimurium that their counterparts colonized with the intestinal microbiota from young mice [42]. A consortium of appropriately selected bacterial strains protected the mouse against infection with S. Typhimurium [41]. Knowledge of the interactions among the host, its microbiota, and pathogenic bacteria helps to establish strategies for modulation of the microbiota and its effective protection of the host against infectious diseases [44].
Salmonella enterica is one of the human and animal enteric pathogens. It consists of a group of over 2500 serovars [45]. A non-typhoidal S. Typhimurium belongs to the most worldwide spread serovars [46,47]. It causes enterocolitis (salmonellosis) in humans and pigs and affects mainly the distal ileum and colon [47,48]. Clinical signs of salmonellosis are fever, anorexia, diarrhea, vomiting, lethargy, and tremor. Salmonellosis is usually a self-limiting illness. However, it can endanger immunocompromised individuals by life-threatening systemic diseases [49].
Our work aimed to study a monocolonization of the intestine of newborn germ-free piglets with mucinolytic and non-mucinolytic strains of Bifidobacterium boum, its impact on the modification of the intestinal barrier, interference of bifidobacteria with S. Typhimurium, and its influence on bacterial translocation.
2. Materials and Methods
2.1. Ethics Statement
All animal experiments were reviewed and approved by the Animal Care and Use Committee of the Czech Academy of Sciences, protocol #117/2012.
2.2. Isolation of Wild Pig Indigenous Bifidobacteria
Bifidobacteria were isolated from the fresh fecal samples of 2 wild pigs using Wilkins–Chalgren agar (Oxoid, Basingstoke, UK) supplemented with soya peptone (5 g/L; Oxoid), mupirocin (100 mg/L), and glacial acetic acid (1 mL/L) in CO2/H2/N2 (10/10/80%) atmosphere in anaerobic jars with palladium catalysts (Oxoid). Cultures were grown in anaerobic Wilkins–Chalgren broth (Oxoid) supplemented with soya peptone (5 g/L; Oxoid) [50].
2.3. Identification and Characterization of Pig B. boum Strains
Bifidobacteria species were classified using 16S rRNA gene sequencing. For this purpose, the primers fD1 and rP2 [51] were employed. The EZBioCloud 16S rRNA gene-based ID database was used for classification based on the 16S rRNA gene sequence similarity [52]. Furthermore, particular strains were biochemically characterized by the API 50CHL test strips (BioMérieux, Marcy l’Etoile, France), allowing us to evaluate the ability to utilize 48 carbohydrate substrates [53].
2.4. Mucinolytic Activity of B. boum Strains
The mucinolytic activity of the B. boum strains was tested using a medium where porcine mucin (Sigma-Aldrich, St. Luis, MO, USA) was used as a selective factor [54]. Bifidobacterial cultures were grown in Wilkins–Chalgren broth (Oxoid) supplemented with soya peptone (5 g/L; Oxoid), were twice centrifuged (5000× g for 10 min), twice flushed with sterile saline, and resuspended in the PBS buffer. The cultures were inoculated (0.1%) into the broth with mucin and incubated at 37 °C for 48 h. Mucin utilization was accompanied by a change of bromocresol purple indicator (Sigma-Aldrich) from violet to yellow color as a consequence of reduced pH value through final products (mainly acetic and lactic acid) of specific fermentative metabolism of bifidobacteria. Cultivation was done under anaerobic conditions by displacing atmospheric oxygen with purified carbon dioxide.
2.5. Salmonella Typhimurium Strain LT2
Salmonella enterica serovar Typhimurium strain LT2 (S. Typhimurium or LT2) was from a collection of the microorganisms of the Institute of Microbiology of the Czech Academy of Sciences (Novy Hradek, Czech Republic). This S. Typhimurium strain was virulent for 1-week-old germ-free piglets [55].
2.6. Bacterial Suspensions
Fresh cultures of bacteria were prepared for each experiment by cultivation at 37 °C overnight. Bifidobacteria were cultivated anaerobically in vials containing 10 mL of Wilkins–Chalgren broth (Oxoid) supplemented with soya peptone (5 g/L, Oxoid). The cells were harvested by centrifugation at 4000× g for 10 min at RT, and the cell pellet was washed twice with 0.05 M phosphate buffer, pH 6.5 containing 500 mg/L cysteine (PBC). S. Typhimurium was cultivated on meat-peptone agar slopes (blood agar base; Oxoid). Bifidobacteria and S. Typhimurium were resuspended to 8 log CFU/mL density in PBC. The evaluated cell densities at 600 nm were verified by cultivation methods on appropriate agar. The cultivation of bifidobacteria was performed in 50 mm Petri dishes with Wilkins–Chalgren agar (Oxoid) supplemented with soya peptone (5 g/L, Oxoid), L-cysteine (0.5 g/L, Merck, Darmstadt, Germany), mupirocin (100 mg/L, Merck), and glacial acetic acid (1 mL/L, Merck). The Petri dishes were cultivated in an anaerostat at 37 °C for 48 h. S. Typhimurium was cultivated aerobically in a 90 mm Petri dish with MacConkey agar (Merck) at 37 °C for 24 h. The bifidobacteria and S. Typhimurium CFU were counted from the dishes optimally containing 10–100 colonies and 20–200 colonies, respectively.
2.7. Gnotobiotic Piglets
Germ-free piglets of miniature Minnesota based breed [56] (Animal Research Institute, Kostelec nad Orlici, Czech Republic) were obtained by hysterectomy on the 112th day of gestation. They were reared in fiberglass isolators with a heated floor, fed to satiety 6–7 times per day with cow’s milk-based formula by a nipple, and microbiologically tested as described in detail elsewhere [57].
2.8. Experimental Design
Each piglet group was composed of 3 independent hysterectomies. A total of 36 gnotobiotic piglets were assigned to 6 groups with 6 piglets per group (Figure 1): (i) Germ-free for the whole experimental period (GF); (ii) 1-week-old GF piglets orally infected with 6 log CFU of S. Typhimurium for 24 h (LT2); (iii) orally colonized with 8 log CFU of RP36 4 h and 24 h after hysterectomy (RP36); (iv) 1-week-old RP36 piglets orally infected with 6 log CFU of LT2 for 24 h (RP36+LT2); (v) orally colonized with 8 log CFU of RP37 4 h and 24 h after hysterectomy (RP37); (vi) 1-week-old RP37 piglets orally infected with 6 log CFU of LT2 for 24 h (RP37+LT2). The bacteria were applied orally in 5 mL of a milk diet, and GF piglets obtained 5 mL of milk only without bacteria. Twenty-four hours after the challenge with Salmonella (LT2, RP36+LT2, and RP37+LT2), the piglets were euthanized by exsanguination via cardiac puncture under isoflurane anesthesia. Their non-infected counterparts GF, RP36, and RP37, proceeded in the same way at the same age.
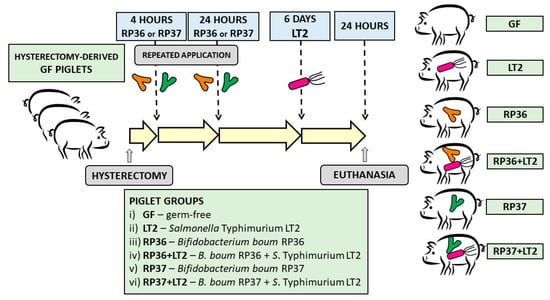
Figure 1.
Schema of the experiment. The gnotobiotic piglets (n = 36) were assigned into six groups: (i) Germ-free (GF, n = 6); (ii) infected with S. Typhimurium strain LT2 for 24 h (LT2, n = 6); (iii) colonized with Bifidobacterium boum strain RP36 (RP36, n = 6); (iv) RP36 infected with S. Typhimurium for 24 h (RP36+LT2, n = 6); (v) colonized with B. boum strain RP37 (RP37, n = 6); (vi) RP37 infected with S. Typhimurium for 24 h (RP37+LT2, n = 6).
2.9. Clinical Signs
All piglets were examined for fever, anorexia, somnolence, and diarrhea during each feeding.
2.10. Bacterial Colonization of the GIT and Translocation
Samples of peripheral blood were cultivated log 10 diluted by PBC. Jejunum (40 cm of the proximal part of the jejunum) and ileum (40 cm segment of a distal part of the small intestine containing the distal jejunum and the ileum) were cut off, filled with 2 mL of PBC, gently kneaded, and rinsed. The whole colon was cut into small pieces on a 90 mm Petri dish and lavaged in 4 mL of PBC, 0.2 g of mesenteric lymph nodes, liver, and spleen were homogenized in 0.8 mL deionized water in a 2 mL Eppendorf tube containing 23.2 mm stainless steel beads in a TissueLyser LT beadbeater (Qiagen, Hilden, Germany). The lavages, tissue homogenates, and blood were serially diluted in PBC. The cultivation for bifidobacteria and S. Typhimurium was performed as the above-described for verification of CFU counts.
2.11. Blood Plasma
A citrated blood was centrifugated at 1200× g for 10 min at 8 °C. A protease inhibitor cocktail (Roche Diagnostics, Manheim, Germany) was added to plasma and the specimens were stored at −45 °C until followed processing.
2.12. Goblet Cells in the Ileum and Colon
The terminal ileum and colon specimens were fixed in Carnoy’s fluid for 30 min, dehydrated, and embedded in paraffin. Five µm cross-sections of tissue were stained with Alcian Blue and post-stained with Nuclear Fast Red (both Diapath, Martinengo, Italy) for mucin production. The preparates were examined under an Olympus BX 40 microscope with an Olympus Camedia C-2000 digital camera (Olympus, Tokyo, Japan). The number of mucin-producing goblet cells in the ileum and colon was counted as cells stained with Alcian Blue. The density of goblet cells per area of the tunica mucosa was counted.
2.13. Total RNA Isolation and Reverse Transcription
RNA was isolated and cDNA synthesized a described previously [43]. Briefly, the RNAlater stored cross-sections of the terminal ileum, and transversal colon was homogenized with 2 mm zirconia beads (BioSpec Products, Bartlesville, OK, USA) in TissueLyser LT beadbeater (Qiagen, Hilden, Germany) and total RNA isolated by the RNeasy Plus Mini kit (Qiagen). Of total RNA, 500 ng was reverse transcribed by QuantiTect Reverse Transcription kit (Qiagen). The synthesized cDNA was 1/10 diluted with PCR quality water (Life Technologies, Carlsbad, CA, USA), and these PCR templates were stored at −25 °C until the Real-Time PCR.
2.14. Real-Time PCR
Two µl of the PCR template was added to 18 µl of the FastStart Universal Probe Master (Roche Diagnostics) containing 500 nM each of the forward and reverse primers (Generi-Biotech, Hradec Kralove, Czech Republic) and 100 nM locked nucleic acid (LNA) probe (Universal ProbeLibrary; Roche Diagnostics) (Table 1). Amplification was performed in duplicates in 45 cycles with heating at 95 °C for 15 s and 60 °C for 60 s on an iQ cycler (Bio-Rad, Hercules, CA, USA). Relative mRNA expressions were calculated by the 2-∆CT method [58] and normalized to β-actin and cyclophilin A by GenEx 6 software (MultiD Analyses AB, Gothenburg, Sweden).

Table 1.
LNA probe-based Real-Time PCR systems.
2.15. Luminex xMAP Technology
Plasma levels of IL-8, TNF-α, and IL-10 were measured by a paramagnetic sphere-based xMAP technology (Luminex Corporation, Austin, TX, USA) with a Porcine ProcartaPlex kit (Affymetrix, Santa Clara, CA, USA) on the Bio-Plex and evaluated by Bio-Plex Manager 4.01 software (Bio-Rad, Hercules, CA, USA), as described previously [43].
2.16. Statistical Analysis
Differences among the groups in parameters with normal distribution were evaluated with one-way analysis of variance (ANOVA) with Tukey’s multiple comparisons post-hoc test. Values that did not meet the normal distribution were evaluated with Kruskal–Wallis with Dunn’s multiple comparisons post-hoc test. The statistical comparisons were performed at p ˂ 0.05 by GraphPad 6 software (GraphPad Software, San Diego, CA, USA) and statistical differences depicted in figures by a letter system.
3. Results
3.1. Isolation, Characterization, and Selection of Pig Indigenous Bifidobacterium boum Strains
In total, 17 strains of B. boum having the 16S rRNA gene similarities ≥99.71% to the B. boum type strain [59] were isolated from the fresh colonic samples of wild pigs. Then, all B. boum isolates were tested for mucinolytic activity, as described in previous studies [54,60]. B. boum designed RP36 was only one strain that showed mucinolytic activity, and its RP37 as its non-mucinolytic counterpart was taken for experiments. The RP36 strain revealed 99.71% and RP37 strain 99.85% 16S rRNA gene sequence similarity to the reference type strain. Their 16S rRNA gene sequences are deposited in the databases GenBank/EMBL/DDBJ under accession numbers MT742662 and MT742663, respectively. The resulted pattern was similar to the type strain of B. boum [61].
3.2. Clinical Signs of Salmonellosis
The germ-free (GF) and B. boum monocolonized non-infected piglets (RP36 and RP37) thrived. The piglets infected with S. Typhimurium LT2 (LT2, LT2+RP36, and LT2+RP37) suffered from anorexia, somnolence, fever, and non-bloody diarrhea.
3.3. Colonization of the Intestine with Bifidobacterium boum RP36 and RP37 Strains, Their Translocation to Mesenteric Lymph Nodes, Blood, Liver, Spleen, and Lungs and Interference with Salmonella Typhimurium
Both B. boum strains RP36 and RP37 were able to colonize the intestine of the gnotobiotic piglet in comparable densities around 4 log CFU/mL in the jejunum (Figure 2A), 6 log CFU/mL in the ileum (Figure 2B), and 9 log CFU in the colon (Figure 2C). No bacteremia or translocation into the mesenteric lymph nodes, liver, spleen, and lungs were observed (Figure 2D). The influence of S. Typhimurium on the growth of B. boum in the intestine was studied by comparison of the groups RP36 versus RP36+LT2 and RP37 versus RP37+LT2 (Figure 2A–C).
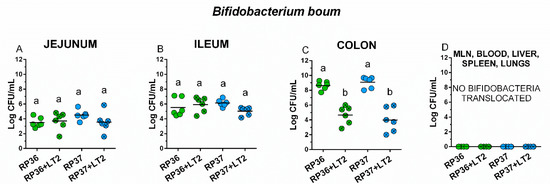
Figure 2.
Colonization of the intestine of the gnotobiotic piglets with B. boum strains RP36 and RP37. B. boum strains RP36 and RP37 colony forming units (CFU) were counted in the jejunum (A), ileum (B), colon (C), mesenteric lymph nodes (MLN), blood, liver, spleen, and lungs (all (D)) in the B. boum monocolonized piglets (RP36 and RP37) and B. boum monocolonized piglets infected with S. Typhimurium (RP36+LT2 and RP37+LT2). Interferences between both B. boum strains (RP36 and RP37) and S. Typhimurium (RP36+LT2 and RP37+LT2) were evaluated by one-way ANOVA with Tuckey’s multiple comparisons post-hoc test. Statistical differences were marked by a letter system at p < 0.05. The same letter means no statistical significance. Log CFU are depicted as individual spots with mean as a horizontal line and n = 6 for all groups.
No differences between B. boum alone (RP36 and RP37) and in the presence of S. Typhimurium (RP36+LT2 and RP37+LT2) were found in the jejunum (Figure 2A) and ileum (Figure 2B). In the colon (Figure 2C), the presence of S. Typhimurium in RP36+LT2 and RP37+LT2 groups statistically significantly decreased the counts of both B. boum strains (Figure 2C). The presence of S. Typhimurium did not cause B. boum bacteremia or its translocation into observed organs (Figure 2D).
3.4. Colonization of the Intestine with Salmonella Typhimurium, Its Translocation to Mesenteric Mesenteric Lymph Nodes, Blood, Liver, Spleen, and Lungs, and Interference with Bifidobacterium boum RP36 and RP37 Strains
Salmonella Typhimurium counts reached around 6 log CFU/mL in the jejunum, 8 log CFU/mL in the ileum, 9 log CFU/mL in colon, 6 log CFU/g in mesenteric lymph nodes, 2 log CFU/mL in blood, and 3 log CFU/g in the liver, spleen, and lungs as it is depicted in Figure 3A–H. Salmonella growth was suppressed in the jejunum, but only in the case of RP36 was this suppression statistically significant. In the ileum (Figure 3B) and colon (Figure 3C), both B. boum strains suppressed S. Typhimurium counts statistically significantly. This suppression by RP37 strain was statistically significantly higher than by the RP36 strain (Figure 3C). Neither RP36 nor RP37 influences Salmonella translocation into the mesenteric lymph nodes (Figure 3D). Bacteremia of S. Typhimurium was statistically significantly higher in the presence of the RP36 strain but non-significantly lower than in the presence of the RP37 strain (Figure 3E). The differences between Salmonella counts in the presence of RP36 and RP37 B. boum strains were not significant. The same statistical relations, as in the case of bacteremia, were in translocations to the liver (Figure 3F) and spleen (Figure 3G). The S. Typhimurium counts in the lungs were not influenced by the presence of any B. boum strain (Figure 3H).

Figure 3.
S. Typhimurium counts in the intestine, mesenteric lymph nodes (MLN), blood, liver, spleen, and lungs. S. Typhimurium strain LT2 (LT2) colony-forming units (CFU) were counted in the jejunum (A), ileum (B), colon (C), mesenteric lymph nodes (MLN; (D)), blood (E), liver (F), spleen (G), and lungs (H). S. Typhimurium (LT2) and its interferences with B. boum strains RP36 (RP36+LT2), and RP37 (RP37+LT2) were evaluated by one-way ANOVA with Tukey’s multiple comparisons post-hoc test. Statistical differences were marked by a letter system at p < 0.05. The same letter means no statistical significance. Log CFU are depicted as individual spots with mean as a horizontal line and n = 6 for all groups.
3.5. Density of Goblet Cells in the Ileum and Colon
The non-infected ileum of the GF, RP36, and RP37 groups were rich for vacuolized enterocytes (Figure 4A–C). The Salmonella infection shortened villi and reduced the number of the vacuolated enterocytes (Figure 4D–F). The infected ileum also showed edema and infiltration of neutrophils. The piglets colonized with mucinolytic B. boum RP36 and infected with S. Typhimurium (RP36+LT2) was the only group that showed a statistically significant decrease of the goblet cell density compare to the GF piglets (Figure 4G).

Figure 4.
Goblet cells density in the ileum of gnotobiotic piglets: Germ-free (GF; (A)), colonized with B. boum RP36 (RP36; (B)), colonized with B. boum RP37 (RP37; (C)), infected with S. Typhimurium LT2 (LT2; (D)), colonized with RP36 and infected with LT2 (RP36+LT2; (E)), and colonized with RP37 and infected with LT2 (RP37+LT2; (F)). Bars represent 500 μm. The goblet cell counts (G) are presented as mean + SEM. Statistical differences were calculated by one-way ANOVA with Tukey’s multiple comparison post-hoc test, and p-values < 0.05 are denoted with different letters above the columns. Six samples in each group were analyzed.
In the colon, bifidobacteria (Figure 5A–C) did not influence the number of mucin-producing cells in comparison with the GF group (Figure 5G). All Salmonella-infected groups (Figure 5D–F) showed statistically significantly lower goblet cell density (Figure 5G). Neither B. boum RP36 nor RP37 prevented a decrease of the goblet cell density (Figure 5E,F). Similar to the ileum, the infected colon was infiltrated with neutrophils.
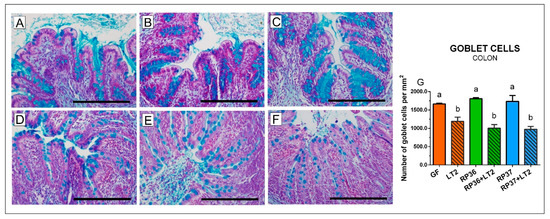
Figure 5.
Goblet cells density in the colon of gnotobiotic piglets: Germ-free (GF; (A)), colonized with B. boum RP36 (RP36; (B)), colonized with B. boum RP37 (RP37; (C)), infected with S. Typhimurium LT2 (LT2; (D)), colonized with RP36 and infected with LT2 (RP36+LT2; (E)), and colonized with RP37 and infected with LT2 (RP37+LT2; (F)). Bars represent 500 µm. The goblet cell counts (G) are presented as mean + SEM. Statistical differences were calculated by one-way ANOVA with Tukey’s multiple comparison post-hoc test, and p-values < 0.05 are denoted with different letters above the columns. Six samples in each group were analyzed.
3.6. Expression of Mucin 2 and Villin mRNA in the Ileum and Colon
The expression of mRNA for mucin 2 in the ileum and colon was influenced neither by colonization with B. boum strains nor infection with S. Typhimurium (Figure 6A,B). In contrast, the villin expression was statistically significantly reduced by infection with S. Typhimurium (Figure 6C,D). Neither B. boum strain RP36 nor RP37 prevented this reduction.
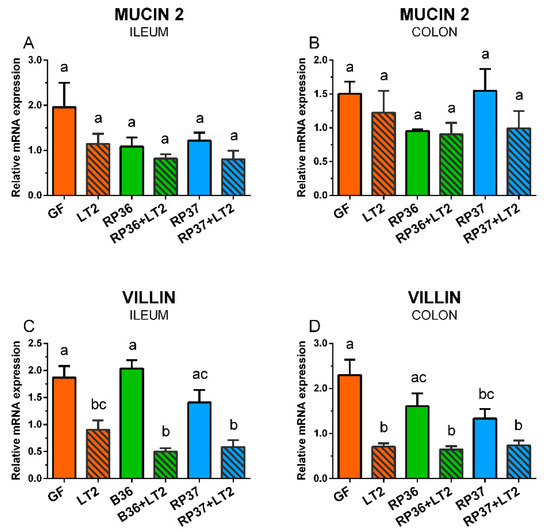
Figure 6.
Expression of mucin 2 (A,B) and villin (C,D) mRNA in the ileum (A,C) and colon (B,D) of the gnotobiotic piglets: Germ-free (GF), infected with S. Typhimurium LT2 (LT2), colonized with B. boum RP36 (RP36), colonized with RP36 and infected with LT2 (RP36+LT2), colonized with B. boum RP37 (RP37), and colonized with RP37 and infected with LT2 (RP37+LT2). The values are presented as mean + SEM. Statistical differences were calculated by one-way ANOVA with Tukey’s multiple comparison post-hoc test, and p-values < 0.05 are denoted with different letters above the columns. Six samples in each group were analyzed.
3.7. Expression of Tight Junction Protein mRNA in the Ileum and Colon
Claudin-1 was upregulated by infection with S. Typhimurium in the ileum (Figure 7A). The increase induced by S. Typhimurium alone (LT2) was statistically non-significant, but both B. boum strains emphasized these changes in the RP36+LT2 and RP37+LT2 groups. A similar pattern of the S. Typhimurium stimulatory effect was observed in the colon, but without any statistical significance compared to the GF piglets (Figure 7B). The infection did not show any visible effect on the claudin-2 in the ileum (Figure 7C). However, the Salmonella infection downregulated the expression of claudin-2 in the colon of all S. Typhimurium-infected groups (Figure 7D). In contrast to claudin-1 in the ileum, occludin was statistically significantly downregulated by S. Typhimurium in comparison with the GF piglets (Figure 7E). No statistically significant differences in the occludin expression were found in the colon (Figure 7F).
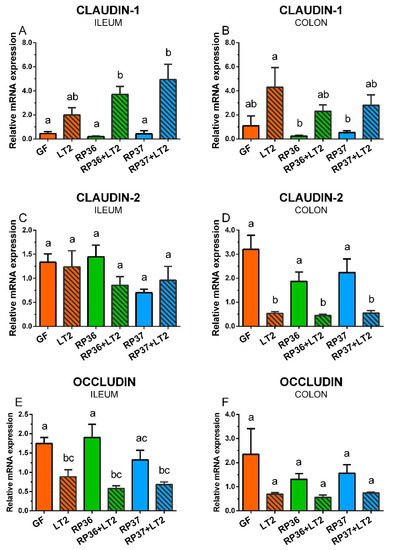
Figure 7.
Expression of claudin-1 (A,B), claudin-2 (C,D), and occludin (E,F) mRNA in the ileum (A,C,E) and colon (B,D,F) of the gnotobiotic piglets: Germ-free (GF), infected with S. Typhimurium LT2 (LT2), colonized with B. boum RP36 (RP36), colonized with RP36 and infected with LT2 (RP36+LT2), colonized with B. boum RP37 (RP37), and colonized with RP37 and infected with LT2 (RP37+LT2). The values are presented as mean + SEM. Statistical differences were calculated by one-way ANOVA with Tukey’s multiple comparison post-hoc test, and p-values < 0.05 are denoted with different letters above the columns. Six samples in each group were analyzed.
3.8. Plasmatic Levels of Inflammatory Cytokines IL-8, TNF-α, and IL-10
Plasmatic levels of IL-8 (Figure 8A) were not detectable in the GF, RP36, and RP37 groups. In contrast, they were induced in all groups infected with S. Typhimurium (LT2, RP36+LT2, and RP37+LT2). However, a statistically different increase was in the LT2 group only. Both piglet groups preliminary colonized with B. boum strains were not statistically different neither from the LT2 group nor in non-infected piglets of all three groups. TNF-α was significantly increased in the LT2 and RP36+LT2 groups but not in the RP37+LT2 group (Figure 8B). The differences among all LT2 infected groups were statistically non-significant. IL-10 showed the same statistical profile with differences among groups as TNF-α (Figure 8C). The ratio IL-10/TNF-α was statistically significantly increased in all Salmonella-infected groups (Figure 8D).

Figure 8.
Plasmatic levels of IL-8 (A), TNF-α (B), IL-10 (C), and ratio IL-10/TNF-α (D) in the gnotobiotic piglets: Germ-free (GF), infected with S. Typhimurium LT2 (LT2), colonized with B. boum RP36 (RP36), colonized with RP36 and infected with LT2 (RP36+LT2), colonized with B. boum RP37 (RP37), and colonized with RP37 and infected with LT2 (RP37+LT2). The values are presented as boxes indicating the lower and upper quartiles, the central line is the median, and the ends of the whiskers depicts the minimal and maximal values. Statistical differences were calculated by the Kruskal-Wallis test with Dunn’s multiple comparison post-hoc test, and p-values < 0.05 are denoted with different letters above the columns. Six samples in each group were analyzed.
4. Discussion
Humans and pigs show high similarity in their gastrointestinal microbiota [33,38]. Therefore, the newborn piglets can be transplanted by human microbiota [40,62,63], and preterm piglets have been used as a model of vulnerable preterm infants [57,63,64]. Bifidobacteria are the most abundant members of newborn intestinal microbiota. They are obligatory anaerobes, and their growth in oxygen presence is limited [15]. Pigs are commonly inhabited by various bifidobacterial species [65,66]. It was proved that both orally simultaneously applied Lactobacillus rhamnosus GG and Bifidobacterium animalis subsp. lactis successfully colonized the sterile intestine of germ-free piglets [67]. Other experiments showed that indigenous porcine Bifidobacterium choerinum, even alone without the support of any other oxygen-consuming bacteria, colonized the intestine of the germ-free piglets [68]. We chose for our experiments B. boum for its typical presence in the intestinal tract of domesticated and wild pigs [59,61,65,66].
The bacteria-rich intestinal lumen and sterile host body are separated by a thin cell monolayer composed of enterocytes. This layer is covered by mucus that forms high molecular mass oligomeric glycoproteins referred to as mucins. Thus, the mucus is the first sentinel that protects the host against bacterial translocation. The mucus layer is continuously secreted and moves to clear trapped material and remove it from epithelial surfaces. The main secreted gel-forming mucin of the healthy human and mouse intestine is mucin 2 (MUC2) [30]. Mucinolytic ability rarely occurs among commonly beneficial bifidobacteria, with the exception of Bifidobacterium bifidum [60]. The absence of both mucinolytic activity and translocation were chosen as criteria confirming beneficial properties of probiotic Bifidobacterium longum BB536, Bifidobacterium breve M-16V, and Bifidobacterium infantis M-63 [69]. Therefore, we aimed to isolate typical porcine indigenous bifidobacterial strain but with mucinolytic activity. The isolated B. boum strain RP36 was only one of 17 tested B. boum strains that showed mucinolytic activity. The B. boum strain RP37 was taken into experiments as the RP36 non-mucinolytic counterpart. We were interested in how the modification of the mucus can influence the intestinal barrier and possible bacterial translocation as bifidobacteria, so S. Typhimurium.
Both B. boum strains were applied a short time after hysterectomy and on a subsequent day. They successfully and comparably colonized the jejunum, ileum, and colon of the colostrum-free germ-free piglets. However, neither B. boum strain RP36 nor strain RP37 translocated and caused bacteremia. It testified about the safety of both bifidobacterial strains [69] for the colostrum-free gnotobiotic piglets. In contrast to commonly beneficial bifidobacteria, Salmonella enterica serovar Typhimurium is an enteric pathogen, and its infection in humans and pigs is associated with enterocolitis [47,48]. S. Typhimurium is sophisticated bacteria that can manipulate the complex conventional microbiota composition to its profit [70]. The used strain LT2 of S. Typhimurium is often called “laboratory strain” [71]. It is almost avirulent for one-week-old conventional piglets [72] but lethal for germ-free piglets of the same age [55]. We found that bifidobacterial counts of both strains of B. boum in monocolonized piglets or in the piglets subsequently infected with S. Typhimurium were comparable in the jejunum and ileum, but they were diminished in the colon. S. Typhimurium stimulates the host to produce oxidizing metabolites that suppress the growth of strictly anaerobic bifidobacteria [15,73]. The colon is the main site of bifidobacteria protective action. This protective action consists mainly of acetic acid production that prevents infection with enteric pathogens [17]. Both mucinolytic RP36 and non-mucinolytic RP37 B. boum strains did not translocate to body organs in the presence of S. Typhimurium. Despite the fact that Salmonella counts were decreased in the intestine in the preliminary colonization of the germ-free piglets with both B. boum strains, all Salmonella-infected piglets suffered diarrhea. S. Typhimurium extensively translocated into the blood and spread to the liver and spleen. The preliminary colonization with the mucinolytic RP36 B. boum strain this translocation increased, but such effect was not observed in non-mucinolytic RP37 B. boum strain. The mucinolytic B. boum probably partially disrupted the intestinal barrier and facilitated Salmonella translocation [74,75].
We paid our attention to the mucin MUC2 that is the main secreted gel-forming mucin of the healthy human and mouse intestine. For this purpose, we used Alcian blue staining of acid mucins that are composed mainly of MUC2 [30]. The mucus is composed of two layers. The inner layer of the mouse colonic mucus is tightly attached to the epithelium and prevents bacterial translocation. In contrast, the outer layer is movable and allows colonization with bacteria but also their removal in the direction of intestinal peristalsis. Nevertheless, this mechanism can be disturbed by enteric infections [30,75]. In MUC2-/- mice, bacteria reached crypts that are normally covered by mucin and were in direct contact with the epithelial cells and induced inflammation and cancer in these mice. It points to the importance of a proprietary composition of MUC2 to participate in a mucus barrier and effectively separate bacteria from the epithelia [74]. In our experiments with gnotobiotic piglets, the infection with S. Typhimurium reduced the number of the goblet cells. This reduction was more obvious in the colon than the ileum. However, neither mucinolytic nor non-mucinolytic B. boum prevented this decrease. This reduction of goblet cells can probably be accounted by the expulsion of the cellular content of mucins as the response to infection [30,75]. To target more at MUC2, we analyzed changes in MUC2 mRNA. In contrast to the reduction of the acid mucin-producing goblet cells, we did not find any obvious influence of the Salmonella infection to MUC2 mRNA expression. Thus, we speculate that pre-prepared mucins are stored in goblet cells and continuously released. It takes a longer time to activate de novo synthesis than it was possibly found within 24 h of experimental infection because the goblet cells contain mucin for immediate usage if needed [30].
To analyze possible changes in the enterocyte layer and intestinal barrier, we analyzed mRNA expression of cytoskeletal protein villin and tight junction proteins claudin-1, claudin-2, and occludin. This barrier can be disrupted directly by enteric pathogens as Salmonella [76] or by their toxins [77]. Villin is associated with the microvillar actin filaments in the apical part of the intestinal epithelial cell brush border. It is a marker of enterocyte differentiation [78], participates in the restitution of damaged epithelia [79], and its expression increases as cells migrate from the crypt to the top of villi [78]. The apical side of the enterocyte extends to the tight junctions (TJ) created by TJ proteins, e.g., claudins and occludin. They form a band around the membrane creating a semipermeable joining between adjacent epithelial cells [28]. Villin and disrupted TJ facilitate the initial steps of Salmonella invasion [77,79], and Salmonella can translocate via lymph vessels or blood to the liver, spleen, and other organs [80]. The tight junctions per se enable paracellular absorption of nutrients and water and prevent the host against bacterial translocation as well as penetration of their toxins to the body [77]. The upregulated claudin-1 in both the ileum and colon of the gnotobiotic piglets can be an attempt to seal the intestinal barrier to prevent leakage of water and electrolytes from the body [81]. The opposite trend was found in the colon but not in the ileum for claudin-2. Its colonic transcription was downregulated in the piglets infected with S. Typhimurium. This difference can explain distinct managing with water and electrolytes in the ileum and colon [82] and different functions of claudin-1 and claudin-2 as representatives of the barrier-forming and pore-forming claudins, respectively [28]. In contrast to claudins, occludin participates in macromolecule flux [83] and cell migration [84]. The infection of mice with enterotoxigenic Escherichia coli downregulated occludin mRNA in the jejunum [85] as the infections with S. Typhimurium in our experiments. S. Typhimurium altered TJ by dephosphorylation of occludin by the induction of inflammatory cytokines tumor necrosis (TNF)-α. This alteration of the intestinal barrier resulted in 10 times higher Salmonella translocation and almost 2 times increased recruitment of neutrophils [86]. Similarly, S. Typhimurium induced neutrophil recruitment to the inflamed intestine in streptomycin-treated mice [87]. The neutrophil recruitment in the intestine of gnotobiotic piglets induced by avirulent rough mutants with incomplete lipopolysaccharide (LPS) of S. Infantis strain 1326/28 [88] or S. Typhimurium strain SF1591 [89] protected the piglets against subsequent infection with virulent S. Typhimurium F98 or S. Typhimurium strain LT2, respectively. The relation of virulence and LPS completeness in the gnotobiotic piglets was reported [55,90]. Lowered virulence of rough Salmonella mutants can reflect their lowered ability to bind to mucus and penetrate deeply, as was documented in experiments with streptomycin-treated mice [91].
Cytokines are important for the normal physiological processes of the host organism [92]. However, their exacerbated unregulated production upon some stimulus, e.g., infection or mechanical trauma, can inflict multiple organ dysfunction and host death [93,94]. The elevated systemic levels of inflammatory cytokines are used as markers of infection and sepsis [95]. Inflammatory cytokines, e.g., TNF-α change the permeability of the intestinal barrier by alteration of TJ protein expression [77]. We analyzed systemic levels of cytokines with different actions—chemotactic interleukin (IL)-8 that attracts neutrophils into the inflammatory site and activate them, TNF-α, which is a potent mediator of inflammation and immunoregulatory IL-10 that prevents an excessive inflammatory reaction [96]. The used cytokines are suitable to indicate the severity of enteric infections in the gnotobiotic pigs [97], and their plasmatic levels dramatically increased in the gnotobiotic piglets infected with necrotoxigenic Escherichia coli O55 [98] that obviously suffered from infection compare to their counterparts that thrived [99]. The overproduction of IL-10 was the main predictor of severity and fatal outcome of sepsis [96,100], and an increased ratio of IL-10/TNF-α was found in non-survived human patients [100]. The appearance of detectable levels of IL-10 in plasma also predicts a bad prognosis of surviving in the piglets [55,68,99]. The infection with S. Typhimurium induced the appearance of all inflammatory cytokines in blood plasma and allowed to count the IL-10/TNF-α ratio in contrast to all groups of non-infected piglets with nonmeasurable cytokines. The colonization with B. boum strains did not prevent the appearance of detectable levels of the analyzed inflammatory cytokines, but the colonization with B. boum RP37 prevented the increase plasmatic IL-10 to the comparable levels in both other Salmonella-infected piglet groups. This alleviating effect of B. boum RP37 may be important for the outcome of the infection with the S. Typhimurium LT2 strain.
5. Conclusions
Bifidobacteria are an important component of complex intestinal microbiota that can increase the resistance of the immunocompromised host to enteric infections. Therefore, we colonized the gnotobiotic piglets with intestinal barrier-disrupting mucinolytic B. boum strain RP36 and harmless non-mucinolytic strain RP37. Both B. boum strains reduced S. Typhimurium counts in the intestine, but the mucinolytic RP36 strain increased S. Typhimurium bacteremia and its translocation to the liver and spleen that was facilitated probably by disruption of the mucus layer, changes in the tight junctions, and the epithelial layer remodeling. Sepsis caused by S. Typhimurium was evaluated by the systemic levels of the analyzed inflammatory cytokines. Other studies are needed to induce a higher protective effect of the newborn intestine’s bacterial colonization with defined bacteria to increase resistance against infection with enteric pathogens.
Author Contributions
Conceptualization, I.S., E.V., and A.S.; methodology, I.S., R.P., J.K., Z.S., V.N.B., E.V., H.S.S., and A.S.; validation, I.S. and A.S.; formal analysis, I.S., R.P., J.K., V.N.B., E.V., H.S.S., and A.S.; investigation, I.S., R.P., J.K., Z.S., E.V., H.S.S., and A.S.; resources, I.S. and E.V.; data curation, I.S., E.V., and A.S.; writing—original draft preparation, I.S., and A.S.; writing—review and editing, I.S., J.K., E.V., and A.S.; supervision, I.S. and A.S.; project administration, I.S. and E.V.; funding acquisition, I.S. and E.V. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded from the grant 13-08803S of the Czech Science Foundation and the Institutional Research concept RVO 61388971 of the Institute of Microbiology of the Czech Academy of Sciences and European Regional Development Fund-Project NutRisk Centre, No. CZ.02.1.01/0.0/0.0/16_019/0000845.
Acknowledgments
We are grateful to Jana Machova, Hana Sychrovska, Jarmila Jarkovska, and Vera Jenistova for animal care, microbiological work, and a procession of samples. Special thanks are given to professor V. Rada from the University of Life Sciences in Prague for fruitful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sommer, F.; Backhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Scarpellini, E.; Ianiro, G.; Attili, F.; Bassanelli, C.; De, S.A.; Gasbarrini, A. The human gut microbiota and virome: Potential therapeutic implications. Dig. Liver Dis. 2015, 47, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Moissl-Eichinger, C.; Pausan, M.; Taffner, J.; Berg, G.; Bang, C.; Schmitz, R.A. Archaea are interactive components of complex microbiomes. Trends Microbiol. 2018, 26, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Kapitan, M.; Niemiec, M.J.; Steimle, A.; Frick, J.S.; Jacobsen, I.D. Fungi as Part of the Microbiota and Interactions with Intestinal Bacteria. Curr. Top. Microbiol. Immunol. 2019, 422, 265–301. [Google Scholar] [CrossRef] [PubMed]
- Chabe, M.; Lokmer, A.; Segurel, L. Gut Protozoa: Friends or Foes of the Human Gut Microbiota? Trends Parasitol. 2017, 33, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Swain Ewald, H.A.; Ewald, P.W. Natural Selection, The Microbiome, and Public Health. Yale J. Biol. Med. 2018, 91, 445–455. [Google Scholar]
- Schroeder, B.O.; Backhed, F. Signals from the gut microbiota to distant organs in physiology and disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef]
- Turroni, F.; Milani, C.; Duranti, S.; Lugli, G.A.; Bernasconi, S.; Margolles, A.; Di Pierro, F.; van Sinderen, D.; Ventura, M. The infant gut microbiome as a microbial organ influencing host well-being. Ital. J. Pediatr. 2020, 46, 16. [Google Scholar] [CrossRef]
- Rutayisire, E.; Huang, K.; Liu, Y.; Tao, F. The mode of delivery affects the diversity and colonization pattern of the gut microbiota during the first year of infants’ life: A systematic review. BMC. Gastroenterol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Wang, M.; Radlowski, E.C.; Monaco, M.H.; Fahey, G.C., Jr.; Gaskins, H.R.; Donovan, S.M. Mode of delivery and early nutrition modulate microbial colonization and fermentation products in neonatal piglets. J. Nutr. 2013, 143, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Donovan, S.M. Evolution of the gut microbiome in infancy within an ecological context. Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.C.; Dinsmoor, A.M.; Wang, M.; Donovan, S.M. Microbiome composition in pediatric populations from birth to adolescence: Impact of diet and prebiotic and probiotic interventions. Dig. Dis. Sci. 2020, 65, 706–722. [Google Scholar] [CrossRef]
- Trebichavsky, I.; Rada, V.; Splichalova, A.; Splichal, I. Cross-talk of human gut with bifidobacteria. Nutr. Rev. 2009, 67, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Peano, C.; Pass, D.A.; Foroni, E.; Severgnini, M.; Claesson, M.J.; Kerr, C.; Hourihane, J.; Murray, D.; Fuligni, F.; et al. Diversity of bifidobacteria within the infant gut microbiota. PLoS ONE 2012, 7, e36957. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Milani, C.; Duranti, S.; Ferrario, C.; Lugli, G.A.; Mancabelli, L.; van Sinderen, D.; Ventura, M. Bifidobacteria and the infant gut: An example of co-evolution and natural selection. Cell Mol. Life Sci. 2018, 75, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Barba-Vidal, E.; Castillejos, L.; Lopez-Colom, P.; Rivero, U.M.; Moreno Munoz, J.A.; Martin-Orue, S.M. Evaluation of the probiotic strain Bifidobacterium longum subsp. infantis CECT 7210 capacities to improve health status and fight digestive pathogens in a piglet model. Front. Microbiol. 2017, 8, 533. [Google Scholar] [CrossRef]
- Delcaru, C.; Alexandru, I.; Podgoreanu, P.; Cristea, V.C.; Bleotu, C.; Chifiriuc, M.C.; Bezirtzoglou, E.; Lazar, V. Antagonistic activities of some Bifidobacterium sp. strains isolated from resident infant gastrointestinal microbiota on Gram-negative enteric pathogens. Anaerobe 2016, 39, 39–44. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef]
- Ficara, M.; Pietrella, E.; Spada, C.; Della Casa, M.E.; Lucaccioni, L.; Iughetti, L.; Berardi, A. Changes of intestinal microbiota in early life. J. Matern. Fetal Neonatal Med. 2020, 33, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A new genomic blueprint of the human gut microbiota. Nature 2019, 568, 499–504. [Google Scholar] [CrossRef]
- Nayfach, S.; Shi, Z.J.; Seshadri, R.; Pollard, K.S.; Kyrpides, N.C. New insights from uncultivated genomes of the global human gut microbiome. Nature 2019, 568, 505–510. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Are we really vastly outnumbered? Revisiting the ratio of bacterial to host cells in humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef]
- Allaire, J.M.; Crowley, S.M.; Law, H.T.; Chang, S.Y.; Ko, H.J.; Vallance, B.A. The intestinal epithelium: Central coordinator of mucosal immunity. Trends Immunol. 2018, 1. [Google Scholar] [CrossRef]
- Gunzel, D.; Fromm, M. Claudins and other tight junction proteins. Compr. Physiol. 2012, 2, 1819–1852. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, D.; Ha, Y.; Cho, K.D.; Lee, B.H.; Seo, I.W.; Kim, S.H.; Chae, C. Expression of mucins and trefoil factor family protein-1 in the colon of pigs naturally infected with Salmonella typhimurium. J. Comp. Pathol. 2009, 140, 38–42. [Google Scholar] [CrossRef]
- Linden, S.K.; Florin, T.H.; McGuckin, M.A. Mucin dynamics in intestinal bacterial infection. PLoS ONE 2008, 3, e3952. [Google Scholar] [CrossRef]
- Breschi, A.; Gingeras, T.R.; Guigo, R. Comparative transcriptomics in human and mouse. Nat. Rev. Genet. 2017, 18, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Feng, Q.; Liang, S.; Sonne, S.B.; Xia, Z.; Qiu, X.; Li, X.; Long, H.; Zhang, J.; Zhang, D.; et al. A catalog of the mouse gut metagenome. Nat. Biotechnol. 2015, 33, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Cavaillon, J.M.; Singer, M.; Skirecki, T. Sepsis therapies: Learning from 30 years of failure of translational research to propose new leads. EMBO Mol. Med. 2020, 12, e10128. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.; Sangild, P.T.; Stoll, B.; Thymann, T.; Buddington, R.; Marini, J.; Olutoye, O.; Shulman, R.J. Translational advances in pediatric nutrition and gastroenterology: New insights from pig models. Annu. Rev. Anim. Biosci. 2020, 8, 321–354. [Google Scholar] [CrossRef]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef]
- Waterhouse, A.; Leslie, D.C.; Bolgen, D.E.; Lightbown, S.; Dimitrakakis, N.; Cartwright, M.J.; Seiler, B.; Lightbown, K.; Smith, K.; Lombardo, P.; et al. Modified clinical monitoring assesment criteria for multi-organ failure during bacteremia and sepsis progression in a pig model. Adv. Crit. Care Med. 2018, 1, 2. [Google Scholar]
- Xiao, L.; Estelle, J.; Kiilerich, P.; Ramayo-Caldas, Y.; Xia, Z.; Feng, Q.; Liang, S.; Pedersen, A.O.; Kjeldsen, N.J.; Liu, C.; et al. A reference gene catalogue of the pig gut microbiome. Nat. Microbiol. 2016, 1, 16161. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Paim, F.C.; Kandasamy, S.; Alhamo, M.A.; Fischer, D.D.; Langel, S.N.; Deblais, L.; Kumar, A.; Chepngeno, J.; Shao, L.; et al. Protein malnutrition modifies innate immunity and gene expression by intestinal epithelial cells and human rotavirus infection in neonatal gnotobiotic pigs. mSphere 2017, 2. [Google Scholar] [CrossRef]
- Wang, M.; Donovan, S.M. Human microbiota-associated swine: Current progress and future opportunities. ILAR J. 2015, 56, 63–73. [Google Scholar] [CrossRef]
- Brugiroux, S.; Beutler, M.; Pfann, C.; Garzetti, D.; Ruscheweyh, H.J.; Ring, D.; Diehl, M.; Herp, S.; Lotscher, Y.; Hussain, S.; et al. Genome-guided design of a defined mouse microbiota that confers colonization resistance against Salmonella enterica serovar Typhimurium. Nat. Microbiol. 2016, 2, 16215. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Sakamoto, K.; Seo, S.U.; Pickard, J.M.; Gillilland, M.G., III; Pudlo, N.A.; Hoostal, M.; Li, X.; Wang, T.D.; Feehley, T.; et al. Neonatal acquisition of Clostridia species protects against colonization by bacterial pathogens. Science 2017, 356, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Splichal, I.; Donovan, S.M.; Splichalova, Z.; Bunesova, V.N.; Vlkova, E.; Jenistova, V.; Killer, J.; Svejstil, R.; Skrivanova, E.; Splichalova, A. Colonization of germ-free piglets with commensal Lactobacillus amylovorus, Lactobacillus mucosae, and probiotic E. coli Nissle 1917 and their interference with Salmonella Typhimurium. Microorganisms 2019, 7, 273. [Google Scholar] [CrossRef] [PubMed]
- Baumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef]
- Hurley, D.; McCusker, M.P.; Fanning, S.; Martins, M. Salmonella-host interactions—Modulation of the host innate immune system. Front. Immunol. 2014, 5, 481. [Google Scholar] [CrossRef]
- Campos, J.; Mourao, J.; Peixe, L.; Antunes, P. Non-typhoidal Salmonella in the pig production chain: A comprehensive analysis of Its impact on human health. Pathogens 2019, 8, 19. [Google Scholar] [CrossRef]
- Kaiser, P.; Hardt, W.D. Salmonella typhimurium diarrhea: Switching the mucosal epithelium from homeostasis to defense. Curr. Opin. Immunol. 2011, 23, 456–463. [Google Scholar] [CrossRef]
- Zhang, S.; Kingsley, R.A.; Santos, R.L.; Andrews-Polymenis, H.; Raffatellu, M.; Figueiredo, J.; Nunes, J.; Tsolis, R.M.; Adams, L.G.; Baumler, A.J. Molecular pathogenesis of Salmonella enterica serotype Typhimurium-induced diarrhea. Infect. Immun. 2003, 71, 1–12. [Google Scholar] [CrossRef]
- Wen, S.C.; Best, E.; Nourse, C. Non-typhoidal Salmonella infections in children: Review of literature and recommendations for management. J. Paediatr. Child. Health 2017, 53, 936–941. [Google Scholar] [CrossRef]
- Rada, V.; Petr, J. A new selective medium for the isolation of glucose non-fermenting bifidobacteria from hen caeca. J. Microbiol. Methods 2000, 43, 127–132. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Killer, J.; Sedlacek, I.; Rada, V.; Havlik, J.; Kopecny, J. Reclassification of Bifidobacterium stercoris Kim et al. 2010 as a later heterotypic synonym of Bifidobacterium adolescentis. Int. J. Syst. Evol. Microbiol. 2013, 63, 4350–4353. [Google Scholar] [CrossRef] [PubMed]
- Pechar, R.; Rada, V.; Parafati, L.; Musilova, S.; Bunesova, V.; Vlkova, E.; Killer, J.; Mrazek, J.; Kmet, V.; Svejstil, R. Mupirocin-mucin agar for selective enumeration of Bifidobacterium bifidum. Int. J. Food Microbiol. 2014, 191, 32–35. [Google Scholar] [CrossRef]
- Splichal, I.; Rychlik, I.; Splichalova, I.; Karasova, D.; Splichalova, A. Toll-like receptor 4 signaling in the ileum and colon of gnotobiotic piglets infected with Salmonella Typhimurium or Its isogenic rfa mutants. Toxins 2020, 12, 545. [Google Scholar] [CrossRef]
- Mandel, L.; Travnicek, J. The minipig as a model in gnotobiology. Nahrung 1987, 31, 613–618. [Google Scholar] [CrossRef]
- Splichalova, A.; Slavikova, V.; Splichalova, Z.; Splichal, I. Preterm life in sterile conditions: A study on preterm, germ-free piglets. Front. Immunol. 2018, 9, 220. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Pechar, R.; Killer, J.; Mekadim, C.; Geigerova, M.; Rada, V. Classification of culturable bifidobacterial population from colonic samples of wild pigs (Sus scrofa) based on three molecular genetic methods. Curr. Microbiol. 2017, 74, 1324–1331. [Google Scholar] [CrossRef]
- Killer, J.; Marounek, M. Fermentation of mucin by bifidobacteria from rectal samples of humans and rectal and intestinal samples of animals. Folia Microbiol. 2011, 56, 85–89. [Google Scholar] [CrossRef]
- Pechar, R.; Killer, J.; Salmonova, H.; Geigerova, M.; Svejstil, R.; Svec, P.; Sedlacek, I.; Rada, V.; Benada, O. Bifidobacterium apri sp. nov., a thermophilic actinobacterium isolated from the digestive tract of wild pigs (Sus scrofa). Int. J. Syst. Evol. Microbiol. 2017, 67, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.D.; Kandasamy, S.; Paim, F.C.; Langel, S.N.; Alhamo, M.A.; Shao, L.; Chepngeno, J.; Miyazaki, A.; Huang, H.C.; Kumar, A.; et al. Protein Malnutrition Alters Tryptophan and Angiotensin-Converting Enzyme 2 Homeostasis and Adaptive Immune Responses in Human Rotavirus-Infected Gnotobiotic Pigs with Human Infant Fecal Microbiota Transplant. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef] [PubMed]
- Brunse, A.; Martin, L.; Rasmussen, T.S.; Christensen, L.; Skovsted, C.M.; Wiese, M.; Khakimov, B.; Pieper, R.; Nielsen, D.S.; Sangild, P.T.; et al. Effect of fecal microbiota transplantation route of administration on gut colonization and host response in preterm pigs. ISME J. 2019, 13, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Jiang, P.; Frokiaer, H.; Heegaard, P.M.; Thymann, T.; Sangild, P.T. Delayed development of systemic immunity in preterm pigs as a model for preterm infants. Sci. Rep. 2016, 6, 36816. [Google Scholar] [CrossRef]
- Lamendella, R.; Santo Domingo, J.W.; Kelty, C.; Oerther, D.B. Bifidobacteria in feces and environmental waters. Appl. Environ. Microbiol. 2008, 74, 575–584. [Google Scholar] [CrossRef]
- Killer, J.; Mrazek, J.; Bunesova, V.; Havlik, J.; Koppova, I.; Benada, O.; Rada, V.; Kopecny, J.; Vlkova, E. Pseudoscardovia suis gen. nov., sp. nov., a new member of the family Bifidobacteriaceae isolated from the digestive tract of wild pigs (Sus scrofa). Syst. Appl. Microbiol. 2013, 36, 11–16. [Google Scholar] [CrossRef]
- Chattha, K.S.; Vlasova, A.N.; Kandasamy, S.; Rajashekara, G.; Saif, L.J. Divergent immunomodulating effects of probiotics on T cell responses to oral attenuated human rotavirus vaccine and virulent human rotavirus infection in a neonatal gnotobiotic piglet disease model. J. Immunol. 2013, 191, 2446–2456. [Google Scholar] [CrossRef]
- Splichalova, A.; Trebichavsky, I.; Rada, V.; Vlkova, E.; Sonnenborn, U.; Splichal, I. Interference of Bifidobacterium choerinum or Escherichia coli Nissle 1917 with Salmonella Typhimurium in gnotobiotic piglets correlates with cytokine patterns in blood and intestine. Clin. Exp. Immunol. 2011, 163, 242–249. [Google Scholar] [CrossRef]
- Abe, F.; Muto, M.; Yaeshima, T.; Iwatsuki, K.; Aihara, H.; Ohashi, Y.; Fujisawa, T. Safety evaluation of probiotic bifidobacteria by analysis of mucin degradation activity and translocation ability. Anaerobe 2010, 16, 131–136. [Google Scholar] [CrossRef]
- Arguello, H.; Estelle, J.; Zaldivar-Lopez, S.; Jimenez-Marin, A.; Carvajal, A.; Lopez-Bascon, M.A.; Crispie, F.; O’Sullivan, O.; Cotter, P.D.; Priego-Capote, F.; et al. Early Salmonella Typhimurium infection in pigs disrupts microbiome composition and functionality principally at the ileum mucosa. Sci. Rep. 2018, 8, 7788. [Google Scholar] [CrossRef]
- McClelland, M.; Sanderson, K.E.; Spieth, J.; Clifton, S.W.; Latreille, P.; Courtney, L.; Porwollik, S.; Ali, J.; Dante, M.; Du, F.; et al. Complete genome sequence of Salmonella enterica serovar Typhimurium LT2. Nature 2001, 413, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.C.; Gyles, C.L. Virulence of wild and mutant strains of Salmonella typhimurium in ligated intestinal segments of calves, pigs, and rabbits. Am. J. Vet. Res. 1987, 48, 504–510. [Google Scholar] [PubMed]
- Perez-Lopez, A.; Behnsen, J.; Nuccio, S.P.; Raffatellu, M. Mucosal immunity to pathogenic intestinal bacteria. Nat. Rev. Immunol. 2016, 16, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.; Phillipson, M.; Petersson, J.; Velcich, A.; Holm, L.; Hansson, G.C. The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 15064–15069. [Google Scholar] [CrossRef] [PubMed]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the mucosal barrier to infection. Mucosal. Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Griffiths, G.; Repnik, U.; Hornef, M. Seeing is understanding: Salmonella’s way to penetrate the intestinal epithelium. Int. J. Med. Microbiol. 2018, 308, 97–106. [Google Scholar] [CrossRef]
- Viswanathan, V.K.; Hodges, K.; Hecht, G. Enteric infection meets intestinal function: How bacterial pathogens cause diarrhoea. Nat. Rev. Microbiol. 2009, 7, 110–119. [Google Scholar] [CrossRef]
- West, A.B.; Isaac, C.A.; Carboni, J.M.; Morrow, J.S.; Mooseker, M.S.; Barwick, K.W. Localization of villin, a cytoskeletal protein specific to microvilli, in human ileum and colon and in colonic neoplasms. Gastroenterology 1988, 94, 343–352. [Google Scholar] [CrossRef]
- Lhocine, N.; Arena, E.T.; Bomme, P.; Ubelmann, F.; Prevost, M.C.; Robine, S.; Sansonetti, P.J. Apical invasion of intestinal epithelial cells by Salmonella typhimurium requires villin to remodel the brush border actin cytoskeleton. Cell Host Microbe 2015, 17, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Galen, J.E.; Buskirk, A.D.; Tennant, S.M.; Pasetti, M.F. Live attenuated human Salmonella vaccine candidates: Tracking the pathogen in natural infection and stimulation of host immunity. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Gunzel, D.; Yu, A.S. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [PubMed]
- Kiela, P.R.; Ghishan, F.K. Physiology of intestinal absorption and secretion. Best. Pract. Res. Clin. Gastroenterol. 2016, 30, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Khatib, K.; Guo, S.; Ye, D.; Youssef, M.; Ma, T. Occludin regulates macromolecule flux across the intestinal epithelial tight junction barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G1054–G1064. [Google Scholar] [CrossRef] [PubMed]
- Edelblum, K.L.; Shen, L.; Weber, C.R.; Marchiando, A.M.; Clay, B.S.; Wang, Y.; Prinz, I.; Malissen, B.; Sperling, A.I.; Turner, J.R. Dynamic migration of gammadelta intraepithelial lymphocytes requires occludin. Proc. Natl. Acad. Sci. USA 2012, 109, 7097–7102. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Zhang, F.; He, Y.; Wang, X.; Jin, X.; Zhang, P.; Bi, D. Enterococcus faecium HDRsEf1 elevates the intestinal barrier defense against enterotoxigenic Escherichia coli and regulates occludin expression via activation of TLR-2 and PI3K signalling pathways. Lett. Appl. Microbiol. 2018, 67, 520–527. [Google Scholar] [CrossRef]
- Kohler, H.; Sakaguchi, T.; Hurley, B.P.; Kase, B.A.; Reinecker, H.C.; McCormick, B.A. Salmonella enterica serovar Typhimurium regulates intercellular junction proteins and facilitates transepithelial neutrophil and bacterial passage. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G178–G187. [Google Scholar] [CrossRef]
- Loetscher, Y.; Wieser, A.; Lengefeld, J.; Kaiser, P.; Schubert, S.; Heikenwalder, M.; Hardt, W.D.; Stecher, B. Salmonella transiently reside in luminal neutrophils in the inflamed gut. PLoS ONE 2012, 7, e34812. [Google Scholar] [CrossRef]
- Foster, N.; Lovell, M.A.; Marston, K.L.; Hulme, S.D.; Frost, A.J.; Bland, P.; Barrow, P.A. Rapid protection of gnotobiotic pigs against experimental salmonellosis following induction of polymorphonuclear leukocytes by avirulent Salmonella enterica. Infect. Immun. 2003, 71, 2182–2191. [Google Scholar] [CrossRef]
- Splichal, I.; Trebichavsky, I.; Splichalova, A.; Barrow, P.A. Protection of gnotobiotic pigs against Salmonella enterica serotype Typhimurium by rough mutant of the same serotype is accompanied by the change of local and systemic cytokine response. Vet. Immunol. Immunopathol. 2005, 103, 155–161. [Google Scholar] [CrossRef]
- Splichalova, A.; Splichalova, Z.; Karasova, D.; Rychlik, I.; Trevisi, P.; Sinkora, M.; Splichal, I. Impact of the lipopolysaccharide chemotype of Salmonella enterica serovar Typhimurium on virulence in gnotobiotic piglets. Toxins 2019, 11, 534. [Google Scholar] [CrossRef]
- Nevola, J.J.; Laux, D.C.; Cohen, P.S. In vivo colonization of the mouse large intestine and in vitro penetration of intestinal mucus by an avirulent smooth strain of Salmonella typhimurium and its lipopolysaccharide-deficient mutant. Infect. Immun. 1987, 55, 2884–2890. [Google Scholar] [CrossRef]
- Dinarello, C.A. Historical insights into cytokines. Eur. J. Immunol. 2007, 37 (Suppl. 1), S34–S45. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Cole, E.; Gillespie, S.; Vulliamy, P.; Brohi, K. Multiple organ dysfunction after trauma. Br. J. Surg. 2020, 107, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Pierrakos, C.; Vincent, J.L. Sepsis biomarkers: A review. Crit. Care 2010, 14, R15. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, H.; Zhou, J.; Zhong, Y.; Ali, M.M.; McGuire, F.; Nagarkatti, P.S.; Nagarkatti, M. Role of cytokines as a double-edged sword in sepsis. In Vivo 2013, 27, 669–684. [Google Scholar] [PubMed]
- Splichal, I.; Splichalova, A. Experimental enteric bacterial infections in pigs. J. Infect. Dis. 2018, 218, 504–505. [Google Scholar] [CrossRef] [PubMed]
- Splichalova, A.; Splichal, I.; Sonnenborn, U.; Rada, V. A modified MacConkey agar for selective enumeration of necrotoxigenic E. coli O55 and probiotic E. coli Nissle 1917. J. Microbiol. Methods 2014, 104, 82–86. [Google Scholar] [CrossRef]
- Splichalova, A.; Splichal, I. Local and systemic occurrences of HMGB1 in gnotobiotic piglets infected with E. coli O55 are related to bacterial translocation and inflammatory cytokines. Cytokine 2012, 60, 597–600. [Google Scholar] [CrossRef]
- Gogos, C.A.; Drosou, E.; Bassaris, H.P.; Skoutelis, A. Pro- versus anti-inflammatory cytokine profile in patients with severe sepsis: A marker for prognosis and future therapeutic options. J. Infect. Dis. 2000, 181, 176–180. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).