Exploring the Ecology of Bifidobacteria and Their Genetic Adaptation to the Mammalian Gut

 ,
,
Abstract
1. History and Taxonomy of Bifidobacteria
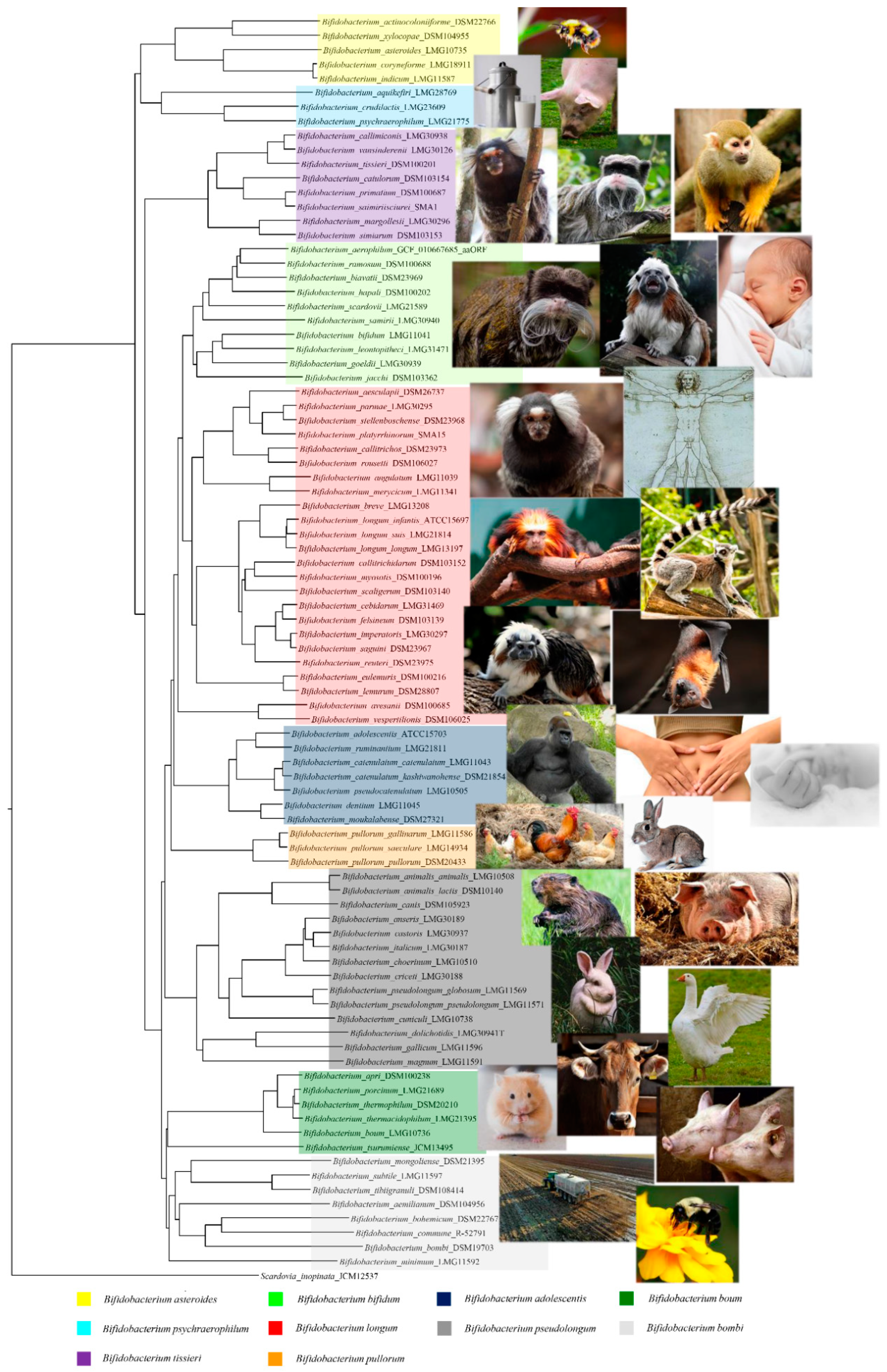
2. Ecology of Bifidobacteria
3. Bifidobacteria and Their Genetic Adaptation to the Human Gut
4. Example of Host–Bifidobacteria and Microbe–Microbe Interactions in the Human Gut
5. Microbe–Microbe Cross-Feeding Activities Influence Intestinal Immune Homeostasis and Inflammatory Response
6. Bifidobacteria as Possible Microbial Biomarkers of Health Predictor
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Van Bergeijk, D.A.; Terlouw, B.R.; Medema, M.H.; van Wezel, G.P. Ecology and genomics of Actinobacteria: New concepts for natural product discovery. Nat. Rev. Microbiol. 2020, 18, 546–558. [Google Scholar] [CrossRef]
- Ventura, M.; Canchaya, C.; Tauch, A.; Chandra, G.; Fitzgerald, G.F.; Chater, K.F.; van Sinderen, D. Genomics of Actinobacteria: Tracing the evolutionary history of an ancient phylum. Microbiol. Mol Biol. Rev. 2007, 71, 495–548. [Google Scholar] [CrossRef]
- Tissier, H. Recherches Sur la Flore Intestinale des Nourrissons: (état Normal et Pathologique). Ph.D. Thesis, G. Carré et C. Naud, Paris, France, 1900; p. 253. [Google Scholar]
- Alessandri, G.; Ossiprandi, M.C.; MacSharry, J.; van Sinderen, D.; Ventura, M. Bifidobacterial Dialogue With Its Human Host and Consequent Modulation of the Immune System. Front. Immunol. 2019, 10, 2348. [Google Scholar] [CrossRef]
- Alberoni, D.; Gaggia, F.; Baffoni, L.; Modesto, M.M.; Biavati, B.; Di Gioia, D. Bifidobacterium xylocopae sp. nov. and Bifidobacterium aemilianum sp. nov., from the carpenter bee (Xylocopa violacea) digestive tract. Syst. Appl. Microbiol. 2019, 42, 205–216. [Google Scholar] [CrossRef]
- Duranti, S.; Lugli, G.A.; Napoli, S.; Anzalone, R.; Milani, C.; Mancabelli, L.; Alessandri, G.; Turroni, F.; Ossiprandi, M.C.; van Sinderen, D.; et al. Characterization of the phylogenetic diversity of five novel species belonging to the genus Bifidobacterium: Bifidobacterium castoris sp. nov., Bifidobacterium callimiconis sp. nov., Bifidobacterium goeldii sp. nov., Bifidobacterium samirii sp. nov. and Bifidobacterium dolichotidis sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 1288–1298. [Google Scholar] [CrossRef]
- Lugli, G.A.; Mangifesta, M.; Duranti, S.; Anzalone, R.; Milani, C.; Mancabelli, L.; Alessandri, G.; Turroni, F.; Ossiprandi, M.C.; van Sinderen, D.; et al. Phylogenetic classification of six novel species belonging to the genus Bifidobacterium comprising Bifidobacterium anseris sp. nov., Bifidobacterium criceti sp. nov., Bifidobacterium imperatoris sp. nov., Bifidobacterium italicum sp. nov., Bifidobacterium margollesii sp. nov. and Bifidobacterium parmae sp. nov. Syst. Appl. Microbiol. 2018, 41, 173–183. [Google Scholar] [CrossRef]
- Trovatelli, L.D.; Crociani, F.; Pedinotti, M.; Scardovi, V. Bifidobacterium pullorum sp. nov.: A new species isolated from chicken feces and a related group of bifidobacteria isolated from rabbit feces. Arch. Microbiol. 1974, 98, 187–198. [Google Scholar] [CrossRef]
- Duranti, S.; Lugli, G.A.; Viappiani, A.; Mancabelli, L.; Alessandri, G.; Anzalone, R.; Longhi, G.; Milani, C.; Ossiprandi, M.C.; Turroni, F.; et al. Characterization of the phylogenetic diversity of two novel species belonging to the genus Bifidobacterium: Bifidobacterium cebidarum sp. nov. and Bifidobacterium leontopitheci sp. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 2288–2297. [Google Scholar] [CrossRef]
- Neuzil-Bunesova, V.; Lugli, G.A.; Modrackova, N.; Makovska, M.; Mrazek, J.; Mekadim, C.; Musilova, S.; Svobodova, I.; Spanek, R.; Ventura, M.; et al. Bifidobacterium canis sp. nov., a novel member of the Bifidobacterium pseudolongum phylogenetic group isolated from faeces of a dog (Canis lupus f. familiaris). Int. J. Syst. Evol. Microbiol. 2020, 70, 5040–5047. [Google Scholar] [CrossRef]
- Modesto, M.; Puglisi, E.; Bonetti, A.; Michelini, S.; Spiezio, C.; Sandri, C.; Sgorbati, B.; Morelli, L.; Mattarelli, P. Bifidobacterium primatium sp. nov., Bifidobacterium scaligerum sp. nov., Bifidobacterium felsineum sp. nov. and Bifidobacterium simiarum sp. nov.: Four novel taxa isolated from the faeces of the cotton top tamarin (Saguinus oedipus) and the emperor tamarin (Saguinus imperator). Syst. Appl. Microbiol. 2018, 41, 593–603. [Google Scholar] [CrossRef]
- Turroni, F.; van Sinderen, D.; Ventura, M. Genomics and ecological overview of the genus Bifidobacterium. Int. J. Food Microbiol. 2011, 149, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Philippe, H.; Douady, C.J. Horizontal gene transfer and phylogenetics. Curr. Opin. Microbiol. 2003, 6, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.; Canchaya, C.; Casale, A.D.; Dellaglio, F.; Neviani, E.; Fitzgerald, G.F.; van Sinderen, D. Analysis of bifidobacterial evolution using a multilocus approach. Int. J. Syst. Evol. Microbiol. 2006, 56, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Lugli, G.A.; Milani, C.; Duranti, S.; Alessandri, G.; Turroni, F.; Mancabelli, L.; Tatoni, D.; Ossiprandi, M.C.; van Sinderen, D.; Ventura, M. Isolation of novel gut bifidobacteria using a combination of metagenomic and cultivation approaches. Genome Biol. 2019, 20, 96. [Google Scholar] [CrossRef] [PubMed]
- Killer, J.; Kopecny, J.; Mrazek, J.; Koppova, I.; Havlik, J.; Benada, O.; Kott, T. Bifidobacterium actinocoloniiforme sp. nov. and Bifidobacterium bohemicum sp. nov., from the bumblebee digestive tract. Int. J. Syst. Evol. Microbiol. 2011, 61, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G. [Comparative Studies on the Bifidus Flora in the Feces of Infants and Adults. With a Contribution to Classification and Nomenclature of Bifidus Strains]. Zentralbl. Bakteriol. Orig. 1963, 191, 486–507. [Google Scholar] [PubMed]
- Michelini, S.; Modesto, M.; Filippini, G.; Spiezio, C.; Sandri, C.; Biavati, B.; Pisi, A.; Mattarelli, P. Bifidobacterium aerophilum sp. nov., Bifidobacterium avesanii sp. nov. and Bifidobacterium ramosum sp. nov.: Three novel taxa from the faeces of cotton-top tamarin (Saguinus oedipus L.). Syst. Appl. Microbiol. 2016, 39, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Toh, H.; Yamazaki, Y.; Tashiro, K.; Kawarai, S.; Oshima, K.; Nakano, A.; Kim, C.N.; Mimura, I.; Arakawa, K.; Iriki, A.; et al. Draft Genome Sequence of Bifidobacterium aesculapii DSM 26737T, Isolated from Feces of Baby Common Marmoset. Genome Announc. 2015, 3. [Google Scholar] [CrossRef]
- Scardovi, V.; Sgorbati, B. Electrophoretic types of transaldolase, transketolase, and other enzymes in bifidobacteria. Antonie Van Leeuwenhoek 1974, 40, 427–440. [Google Scholar] [CrossRef]
- Mitsuoka, T. [Comparative studies on bifidobacteria isolated from the alimentary tract of man and animals (including descriptions of bifidobacterium thermophilum nov. spec. and bifidobacterium pseudolongum nov. spec)]. Zentralbl. Bakteriol. Orig. 1969, 210, 52–64. [Google Scholar]
- Masco, L.; Ventura, M.; Zink, R.; Huys, G.; Swings, J. Polyphasic taxonomic analysis of Bifidobacterium animalis and Bifidobacterium lactis reveals relatedness at the subspecies level: Reclassification of Bifidobacterium animalis as Bifidobacterium animalis subsp. animalis subsp. nov. and Bifidobacterium lactis as Bifidobacterium animalis subsp. lactis subsp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Pechar, R.; Killer, J.; Salmonova, H.; Geigerova, M.; Svejstil, R.; Svec, P.; Sedlacek, I.; Rada, V.; Benada, O. Bifidobacterium apri sp. nov., a thermophilic actinobacterium isolated from the digestive tract of wild pigs (Sus scrofa). Int. J. Syst. Evol. Microbiol. 2017, 67, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- Laureys, D.; Cnockaert, M.; De Vuyst, L.; Vandamme, P. Bifidobacterium aquikefiri sp. nov., isolated from water kefir. Int. J. Syst. Evol. Microbiol. 2016, 66, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Scardovi, V.; Trovatelli, L.D. New species of bifid bacteria from Apis mellifica L. and Apis indica F. A contribution to the taxonomy and biochemistry of the genus Bifidobacterium. Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg. 1969, 123, 64–88. [Google Scholar] [PubMed]
- Endo, A.; Futagawa-Endo, Y.; Schumann, P.; Pukall, R.; Dicks, L.M. Bifidobacterium reuteri sp. nov., Bifidobacterium callitrichos sp. nov., Bifidobacterium saguini sp. nov., Bifidobacterium stellenboschense sp. nov. and Bifidobacterium biavatii sp. nov. isolated from faeces of common marmoset (Callithrix jacchus) and red-handed tamarin (Saguinus midas). Syst. Appl. Microbiol. 2012, 35, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Killer, J.; Kopecny, J.; Mrazek, J.; Rada, V.; Benada, O.; Koppova, I.; Havlik, J.; Straka, J. Bifidobacterium bombi sp. nov., from the bumblebee digestive tract. Int. J. Syst. Evol. Microbiol. 2009, 59, 2020–2024. [Google Scholar] [CrossRef][Green Version]
- Scardovi, V.; Trovatelli, L.D.; Biavati, B.; Zani, G. Bifidobacterium cuniculi, Bifidobacterium choerinum, Bifido bacterium boum, and Bifidobacterium pseudocatenulatum: Four New Species and Their Deoxyribonucleic Acid Homology Relationships. Int. J. Bacteriol. 1979, 29, 291–311. [Google Scholar] [CrossRef]
- Modesto, M.; Michelini, S.; Sansosti, M.C.; De Filippo, C.; Cavalieri, D.; Qvirist, L.; Andlid, T.; Spiezio, C.; Sandri, C.; Pascarelli, S.; et al. Bifidobacterium callitrichidarum sp. nov. from the faeces of the emperor tamarin (Saguinus imperator). Int. J. Syst. Evol. Microbiol. 2018, 68, 141–148. [Google Scholar] [CrossRef]
- Scardovi, V.; Crociani, F. Bifidobacterium catenulatum, Bifidobacterium dentium, and Bifidobacterium angulatum: Three New Species and Their Deox y ribonucleic Acid Homology Relation ships. Int. J. Syst. Bacteriol. 1974, 24, 6–20. [Google Scholar] [CrossRef][Green Version]
- Morita, H.; Nakano, A.; Onoda, H.; Toh, H.; Oshima, K.; Takami, H.; Murakami, M.; Fukuda, S.; Takizawa, T.; Kuwahara, T.; et al. Bifidobacterium kashiwanohense sp. nov., isolated from healthy infant faeces. Int. J. Syst. Evol. Microbiol. 2011, 61, 2610–2615. [Google Scholar] [CrossRef][Green Version]
- Modesto, M.; Michelini, S.; Oki, K.; Biavati, B.; Watanabe, K.; Mattarelli, P. Bifidobacterium catulorum sp. nov., a novel taxon from the faeces of the baby common marmoset (Callithrix jacchus). Int. J. Syst. Evol. Microbiol. 2018, 68, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Praet, J.; Meeus, I.; Cnockaert, M.; Aerts, M.; Smagghe, G.; Vandamme, P. Bifidobacterium commune sp. nov. isolated from the bumble bee gut. Antonie Van. Leeuwenhoek 2015, 107, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Delcenserie, V.; Gavini, F.; Beerens, H.; Tresse, O.; Franssen, C.; Daube, G. Description of a new species, Bifidobacterium crudilactis sp. nov., isolated from raw milk and raw milk cheeses. Syst. Appl. Microbiol. 2007, 30, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Michelini, S.; Modesto, M.; Pisi, A.M.; Filippini, G.; Sandri, C.; Spiezio, C.; Biavati, B.; Sgorbati, B.; Mattarelli, P. Bifidobacterium eulemuris sp. nov., isolated from faeces of black lemurs (Eulemur macaco). Int. J. Syst. Evol. Microbiol. 2016, 66, 1567–1576. [Google Scholar] [CrossRef]
- Lauer, E. Bifidobacterium gallicum sp. nov. isolated from human feces. Int. J. Syst. Bacteriol. 1990, 40, 100–102. [Google Scholar] [CrossRef]
- Michelini, S.; Oki, K.; Yanokura, E.; Shimakawa, Y.; Modesto, M.; Mattarelli, P.; Biavati, B.; Watanabe, K. Bifidobacterium myosotis sp. nov., Bifidobacterium tissieri sp. nov. and Bifidobacterium hapali sp. nov., isolated from faeces of baby common marmosets (Callithrix jacchus L.). Int. J. Syst. Evol. Microbiol. 2016, 66, 255–265. [Google Scholar] [CrossRef]
- Modesto, M.; Watanabe, K.; Arita, M.; Satti, M.; Oki, K.; Sciavilla, P.; Patavino, C.; Camma, C.; Michelini, S.; Sgorbati, B.; et al. Bifidobacterium jacchi sp. nov., isolated from the faeces of a baby common marmoset (Callithrix jacchus). Int. J. Syst. Evol. Microbiol. 2019, 69, 2477–2485. [Google Scholar] [CrossRef]
- Modesto, M.; Michelini, S.; Stefanini, I.; Sandri, C.; Spiezio, C.; Pisi, A.; Filippini, G.; Biavati, B.; Mattarelli, P. Bifidobacterium lemurum sp. nov., from faeces of the ring-tailed lemur (Lemur catta). Int. J. Syst. Evol. Microbiol. 2015, 65, 1726–1734. [Google Scholar] [CrossRef]
- Matteuzzi, D.; Crociani, F.; Zani, G.; Trovatelli, L.D. Bifidobacterium suis n. sp.: A new species of the genus Bifidobacterium isolated from pig feces. Z. Allg. Mikrobiol. 1971, 11, 387–395. [Google Scholar] [CrossRef]
- Biavati, B.; Mattarelli, P. Bifidobacterium ruminantium sp. nov. and Bifidobacterium merycicum sp. nov. from the rumens of cattle. Int. J. Syst. Bacteriol. 1991, 41, 163–168. [Google Scholar] [CrossRef]
- Scardovi, V.; Trovatelli, L.D. Bifidobacterium animalis (Mitsuoka) comb. nov. and the “minimum” and “subtile” Groups of New Bifidobacteria Found in Sewage. Int. J. Syst. Bacteriol. 1974, 24, 21–28. [Google Scholar] [CrossRef][Green Version]
- Watanabe, K.; Makino, H.; Sasamoto, M.; Kudo, Y.; Fujimoto, J.; Demberel, S. Bifidobacterium mongoliense sp. nov., from airag, a traditional fermented mare’s milk product from Mongolia. Int. J. Syst. Evol. Microbiol. 2009, 59, 1535–1540. [Google Scholar] [CrossRef]
- Tsuchida, S.; Takahashi, S.; Nguema, P.P.M.; Fujita, S.; Kitahara, M.; Yamagiwa, J.; Ngomanda, A.; Ohkuma, M.; Ushida, K. Bifidobacterium moukalabense sp. nov., isolated from the faeces of wild west lowland gorilla (Gorilla gorilla gorilla). Int. J. Syst. Evol. Microbiol. 2014, 64, 449–455. [Google Scholar] [CrossRef]
- Modesto, M.; Satti, M.; Watanabe, K.; Scarafile, D.; Huang, C.H.; Liou, J.S.; Tamura, T.; Saito, S.; Watanabe, M.; Mori, K.; et al. Phylogenetic characterization of two novel species of the genus Bifidobacterium: Bifidobacterium saimiriisciurei sp. nov. and Bifidobacterium platyrrhinorum sp. nov. Syst. Appl. Microbiol. 2020, 43, 126111. [Google Scholar] [CrossRef] [PubMed]
- Scardovi, V.; Trovatelli, L.D.; Crociani, F.; Sgorbati, B. Bifid bacteria in bovine rumen. New species of the genus Bifidobacterium: B. globosum n.sp. and B. ruminale n.sp. Arch. Mikrobiol. 1969, 68, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Simpson, P.J.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Bifidobacterium psychraerophilum sp. nov. and Aeriscardovia aeriphila gen. nov., sp. nov., isolated from a porcine caecum. Int. J. Syst. Evol. Microbiol. 2004, 54, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Watabe, J.; Benno, Y.; Mitsuoka, T. BiJidobacterium gallinarum sp. nov.: A New Species Isolated from the Ceca of Chickens. Int. J. Syst. Bacterioly 1983, 33. [Google Scholar] [CrossRef]
- Modesto, M.; Satti, M.; Watanabe, K.; Puglisi, E.; Morelli, L.; Huang, C.H.; Liou, J.S.; Miyashita, M.; Tamura, T.; Saito, S.; et al. Characterization of Bifidobacterium species in feaces of the Egyptian fruit bat: Description of B. vespertilionis sp. nov. and B. rousetti sp. nov. Syst. Appl. Microbiol. 2019, 42, 126017. [Google Scholar] [CrossRef]
- Lugli, G.A.; Milani, C.; Turroni, F.; Duranti, S.; Ferrario, C.; Viappiani, A.; Mancabelli, L.; Mangifesta, M.; Taminiau, B.; Delcenserie, V.; et al. Investigation of the evolutionary development of the genus Bifidobacterium by comparative genomics. Appl. Environ. Microbiol. 2014, 80, 6383–6394. [Google Scholar] [CrossRef]
- Hoyles, L.; Inganas, E.; Falsen, E.; Drancourt, M.; Weiss, N.; McCartney, A.L.; Collins, M.D. Bifidobacterium scardovii sp. nov., from human sources. Int. J. Syst. Evol. Microbiol. 2002, 52, 995–999. [Google Scholar] [CrossRef]
- Zhu, L.; Li, W.; Dong, X. Species identification of genus Bifidobacterium based on partial HSP60 gene sequences and proposal of Bifidobacterium thermacidophilum subsp. porcinum subsp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1619–1623. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Xin, Y.; Jian, W.; Liu, X.; Ling, D. Bifidobacterium thermacidophilum sp. nov., isolated from an anaerobic digester. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 1, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Eckel, V.P.L.; Ziegler, L.M.; Vogel, R.F.; Ehrmann, M. Bifidobacterium tibiigranuli sp. nov. isolated from homemade water kefir. Int. J. Syst. Evol. Microbiol. 2020, 70, 1562–1570. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Benno, Y.; Leung, K.P.; Maeda, N. Bifidobacterium tsurumiense sp. nov., from hamster dental plaque. Int. J. Syst. Evol. Microbiol. 2008, 58, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Duranti, S.; Mangifesta, M.; Lugli, G.A.; Turroni, F.; Anzalone, R.; Milani, C.; Mancabelli, L.; Ossiprandi, M.C.; Ventura, M. Bifidobacterium vansinderenii sp. nov., isolated from faeces of emperor tamarin (Saguinus imperator). Int. J. Syst. Evol. Microbiol. 2017, 67, 3987–3995. [Google Scholar] [CrossRef]
- Alessandri, G.; Milani, C.; Mancabelli, L.; Mangifesta, M.; Lugli, G.A.; Viappiani, A.; Duranti, S.; Turroni, F.; Ossiprandi, M.C.; van Sinderen, D.; et al. The impact of human-facilitated selection on the gut microbiota of domesticated mammals. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef]
- Milani, C.; Mangifesta, M.; Mancabelli, L.; Lugli, G.A.; James, K.; Duranti, S.; Turroni, F.; Ferrario, C.; Ossiprandi, M.C.; van Sinderen, D.; et al. Unveiling bifidobacterial biogeography across the mammalian branch of the tree of life. ISME J. 2017, 11, 2834–2847. [Google Scholar] [CrossRef]
- Wong, C.B.; Odamaki, T.; Xiao, J.Z. Insights into the reason of Human-Residential Bifidobacteria (HRB) being the natural inhabitants of the human gut and their potential health-promoting benefits. FEMS Microbiol. Rev. 2020, 44, 369–385. [Google Scholar] [CrossRef]
- Bottacini, F.; Ventura, M.; van Sinderen, D.; O’Connell Motherway, M. Diversity, ecology and intestinal function of bifidobacteria. Microb. Cell. Fact. 2014, 13 (Suppl. 1), S4. [Google Scholar] [CrossRef]
- Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Sanchez, B.; Margolles, A. Bifidobacteria and Their Molecular Communication with the Immune System. Front. Microbiol. 2017, 8, 2345. [Google Scholar] [CrossRef]
- Turroni, F.; Peano, C.; Pass, D.A.; Foroni, E.; Severgnini, M.; Claesson, M.J.; Kerr, C.; Hourihane, J.; Murray, D.; Fuligni, F.; et al. Diversity of bifidobacteria within the infant gut microbiota. PLoS. ONE 2012, 7, e36957. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the human infant intestinal microbiota. PLoS Biol. 2007, 5, e177. [Google Scholar] [CrossRef] [PubMed]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4578–4585. [Google Scholar] [CrossRef] [PubMed]
- Avershina, E.; Lundgard, K.; Sekelja, M.; Dotterud, C.; Storro, O.; Oien, T.; Johnsen, R.; Rudi, K. Transition from infant- to adult-like gut microbiota. Environ. Microbiol. 2016, 18, 2226–2236. [Google Scholar] [CrossRef]
- Nuriel-Ohayon, M.; Neuman, H.; Koren, O. Microbial Changes during Pregnancy, Birth, and Infancy. Front. Microbiol. 2016, 7, 1031. [Google Scholar] [CrossRef]
- Rautava, S.; Luoto, R.; Salminen, S.; Isolauri, E. Microbial contact during pregnancy, intestinal colonization and human disease. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 565–576. [Google Scholar] [CrossRef]
- Milani, C.; Mancabelli, L.; Lugli, G.A.; Duranti, S.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Ferretti, P.; Gorfer, V.; et al. Exploring Vertical Transmission of Bifidobacteria from Mother to Child. Appl. Environ. Microbiol. 2015, 81, 7078–7087. [Google Scholar] [CrossRef]
- Sela, D.A.; Chapman, J.; Adeuya, A.; Kim, J.H.; Chen, F.; Whitehead, T.R.; Lapidus, A.; Rokhsar, D.S.; Lebrilla, C.B.; German, J.B.; et al. The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc. Natl. Acad. Sci. USA 2008, 105, 18964–18969. [Google Scholar] [CrossRef]
- Ishikawa, E.; Matsuki, T.; Kubota, H.; Makino, H.; Sakai, T.; Oishi, K.; Kushiro, A.; Fujimoto, J.; Watanabe, K.; Watanuki, M.; et al. Ethnic diversity of gut microbiota: Species characterization of Bacteroides fragilis group and genus Bifidobacterium in healthy Belgian adults, and comparison with data from Japanese subjects. J. Biosci. Bioeng. 2013, 116, 265–270. [Google Scholar] [CrossRef]
- Odamaki, T.; Bottacini, F.; Kato, K.; Mitsuyama, E.; Yoshida, K.; Horigome, A.; Xiao, J.Z.; van Sinderen, D. Genomic diversity and distribution of Bifidobacterium longum subsp. longum across the human lifespan. Sci. Rep. 2018, 8, 85. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Foroni, E.; Pizzetti, P.; Giubellini, V.; Ribbera, A.; Merusi, P.; Cagnasso, P.; Bizzarri, B.; de’Angelis, G.L.; Shanahan, F.; et al. Exploring the diversity of the bifidobacterial population in the human intestinal tract. Appl. Environ. Microbiol. 2009, 75, 1534–1545. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and Their Role as Members of the Human Gut Microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef]
- Jacob, R.A. Folate, DNA methylation, and gene expression: Factors of nature and nurture. Am. J. Clin. Nutr. 2000, 72, 903–904. [Google Scholar] [CrossRef] [PubMed]
- Lucock, M. Folic acid: Nutritional biochemistry, molecular biology, and role in disease processes. Mol. Genet. Metab. 2000, 71, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, B.; Weimer, B.C.; Pinzon, J.; Dao Kong, N.; Rompato, G.; Brothersen, C.; McMahon, D.J. Probiotic bacteria survive in Cheddar cheese and modify populations of other lactic acid bacteria. J. Appl. Microbiol. 2014, 116, 1642–1656. [Google Scholar] [CrossRef] [PubMed]
- Heller, K.J. Probiotic bacteria in fermented foods: Product characteristics and starter organisms. Am. J. Clin. Nutr. 2001, 73, 374S–379S. [Google Scholar] [CrossRef]
- Huang, J.S.; Bousvaros, A.; Lee, J.W.; Diaz, A.; Davidson, E.J. Efficacy of probiotic use in acute diarrhea in children: A meta-analysis. Dig. Dis. Sci. 2002, 47, 2625–2634. [Google Scholar] [CrossRef]
- Allen, S.J.; Okoko, B.; Martinez, E.; Gregorio, G.; Dans, L.F. Probiotics for treating infectious diarrhoea. Cochrane Database Syst. Rev. 2004, CD003048. [Google Scholar] [CrossRef]
- Saavedra, J.M.; Bauman, N.A.; Oung, I.; Perman, J.A.; Yolken, R.H. Feeding of Bifidobacterium bifidum and Streptococcus thermophilus to infants in hospital for prevention of diarrhoea and shedding of rotavirus. Lancet 1994, 344, 1046–1049. [Google Scholar] [CrossRef]
- Turroni, F.; Bottacini, F.; Foroni, E.; Mulder, I.; Kim, J.H.; Zomer, A.; Sanchez, B.; Bidossi, A.; Ferrarini, A.; Giubellini, V.; et al. Genome analysis of Bifidobacterium bifidum PRL2010 reveals metabolic pathways for host-derived glycan foraging. Proc. Natl. Acad. Sci. USA 2010, 107, 19514–19519. [Google Scholar] [CrossRef] [PubMed]
- Duranti, S.; Milani, C.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Sanchez, B.; Margolles, A.; et al. Evaluation of genetic diversity among strains of the human gut commensal Bifidobacterium adolescentis. Sci. Rep. 2016, 6, 23971. [Google Scholar] [CrossRef] [PubMed]
- Duranti, S.; Milani, C.; Lugli, G.A.; Turroni, F.; Mancabelli, L.; Sanchez, B.; Ferrario, C.; Viappiani, A.; Mangifesta, M.; Mancino, W.; et al. Insights from genomes of representatives of the human gut commensal Bifidobacterium bifidum. Environ. Microbiol. 2015, 17, 2515–2531. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.E.; Gerdes, S.; Evans, K.C.; Nedveck, D.; Singles, S.K.; Volk, B.; Budinoff, C. Strain-specific strategies of 2’-fucosyllactose, 3-fucosyllactose, and difucosyllactose assimilation by Bifidobacterium longum subsp. infantis Bi-26 and ATCC 15697. Sci. Rep. 2020, 10, 15919. [Google Scholar] [CrossRef]
- O’Connell Motherway, M.; Fitzgerald, G.F.; Neirynck, S.; Ryan, S.; Steidler, L.; van Sinderen, D. Characterization of ApuB, an extracellular type II amylopullulanase from Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 2008, 74, 6271–6279. [Google Scholar] [CrossRef]
- Turroni, F.; Milani, C.; Duranti, S.; Mahony, J.; van Sinderen, D.; Ventura, M. Glycan Utilization and Cross-Feeding Activities by Bifidobacteria. Trends Microbiol. 2018, 26, 339–350. [Google Scholar] [CrossRef]
- Duranti, S.; Turroni, F.; Lugli, G.A.; Milani, C.; Viappiani, A.; Mangifesta, M.; Gioiosa, L.; Palanza, P.; van Sinderen, D.; Ventura, M. Genomic characterization and transcriptional studies of the starch-utilizing strain Bifidobacterium adolescentis 22L. Appl. Environ. Microbiol. 2014, 80, 6080–6090. [Google Scholar] [CrossRef]
- Ventura, M.; Turroni, F.; Motherway, M.O.; MacSharry, J.; van Sinderen, D. Host-microbe interactions that facilitate gut colonization by commensal bifidobacteria. Trends Microbiol. 2012, 20, 467–476. [Google Scholar] [CrossRef]
- Bottacini, F.; van Sinderen, D.; Ventura, M. Omics of bifidobacteria: Research and insights into their health-promoting activities. Biochem. J. 2017, 474, 4137–4152. [Google Scholar] [CrossRef]
- Tremaroli, V.; Backhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Odamaki, T.; Xiao, J.Z. Production of Indole-3-Lactic Acid by Bifidobacterium Strains Isolated fromHuman Infants. Microorganisms 2019, 7, 340. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.B.; Tanaka, A.; Kuhara, T.; Xiao, J.Z. Potential Effects of Indole-3-Lactic Acid, a Metabolite of Human Bifidobacteria, on NGF-induced Neurite Outgrowth in PC12 Cells. Microorganisms 2020, 8, 398. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Ding, Y.; Saedi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut Microbiota-Derived Tryptophan Metabolites Modulate Inflammatory Response in Hepatocytes and Macrophages. Cell Rep. 2018, 23, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.K.; Yoo, K.Y.; Li, H.; Park, O.K.; Lee, C.H.; Choi, J.H.; Jeong, Y.G.; Lee, Y.L.; Kim, Y.M.; Kwon, Y.G.; et al. Indole-3-propionic acid attenuates neuronal damage and oxidative stress in the ischemic hippocampus. J. Neurosci. Res. 2009, 87, 2126–2137. [Google Scholar] [CrossRef] [PubMed]
- Foroni, E.; Serafini, F.; Amidani, D.; Turroni, F.; He, F.; Bottacini, F.; O’Connell Motherway, M.; Viappiani, A.; Zhang, Z.; Rivetti, C.; et al. Genetic analysis and morphological identification of pilus-like structures in members of the genus Bifidobacterium. Microb. Cell Fact. 2011, 10 (Suppl. 1), S16. [Google Scholar] [CrossRef]
- Turroni, F.; Serafini, F.; Foroni, E.; Duranti, S.; O’Connell Motherway, M.; Taverniti, V.; Mangifesta, M.; Milani, C.; Viappiani, A.; Roversi, T.; et al. Role of sortase-dependent pili of Bifidobacterium bifidum PRL2010 in modulating bacterium-host interactions. Proc. Natl. Acad. Sci. USA 2013, 110, 11151–11156. [Google Scholar] [CrossRef]
- O’Connell Motherway, M.; Zomer, A.; Leahy, S.C.; Reunanen, J.; Bottacini, F.; Claesson, M.J.; O’Brien, F.; Flynn, K.; Casey, P.G.; Munoz, J.A.; et al. Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl. Acad. Sci. USA 2011, 108, 11217–11222. [Google Scholar] [CrossRef]
- Scott, J.R.; Zahner, D. Pili with strong attachments: Gram-positive bacteria do it differently. Mol. Microbiol. 2006, 62, 320–330. [Google Scholar] [CrossRef]
- Milani, C.; Mangifesta, M.; Mancabelli, L.; Lugli, G.A.; Mancino, W.; Viappiani, A.; Faccini, A.; van Sinderen, D.; Ventura, M.; Turroni, F. The Sortase-Dependent Fimbriome of the Genus Bifidobacterium: Extracellular Structures with Potential To Modulate Microbe-Host Dialogue. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef]
- Tomich, M.; Planet, P.J.; Figurski, D.H. The tad locus: Postcards from the widespread colonization island. Nat. Rev. Microbiol. 2007, 5, 363–375. [Google Scholar] [CrossRef] [PubMed]
- O’Connell Motherway, M.; Houston, A.; O’Callaghan, G.; Reunanen, J.; O’Brien, F.; O’Driscoll, T.; Casey, P.G.; de Vos, W.M.; van Sinderen, D.; Shanahan, F. A Bifidobacterial pilus-associated protein promotes colonic epithelial proliferation. Mol. Microbiol. 2019, 111, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Potempa, J.; Korzus, E.; Travis, J. The serpin superfamily of proteinase inhibitors: Structure, function, and regulation. J. Biol. Chem. 1994, 269, 15957–15960. [Google Scholar] [PubMed]
- Turroni, F.; Foroni, E.; O’Connell Motherway, M.; Bottacini, F.; Giubellini, V.; Zomer, A.; Ferrarini, A.; Delledonne, M.; Zhang, Z.; van Sinderen, D.; et al. Characterization of the serpin-encoding gene of Bifidobacterium breve 210B. Appl. Environ. Microbiol. 2010, 76, 3206–3219. [Google Scholar] [CrossRef]
- Alvarez-Martin, P.; O’Connell Motherway, M.; Turroni, F.; Foroni, E.; Ventura, M.; van Sinderen, D. A two-component regulatory system controls autoregulated serpin expression in Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 2012, 78, 7032–7041. [Google Scholar] [CrossRef]
- Alvarez-Martin, P.; Fernandez, M.; O’Connell-Motherway, M.; O’Connell, K.J.; Sauvageot, N.; Fitzgerald, G.F.; MacSharry, J.; Zomer, A.; van Sinderen, D. A conserved two-component signal transduction system controls the response to phosphate starvation in Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 2012, 78, 5258–5269. [Google Scholar] [CrossRef]
- Ivanov, D.; Emonet, C.; Foata, F.; Affolter, M.; Delley, M.; Fisseha, M.; Blum-Sperisen, S.; Kochhar, S.; Arigoni, F. A serpin from the gut bacterium Bifidobacterium longum inhibits eukaryotic elastase-like serine proteases. J. Biol. Chem. 2006, 281, 17246–17252. [Google Scholar] [CrossRef]
- Ferrario, C.; Milani, C.; Mancabelli, L.; Lugli, G.A.; Duranti, S.; Mangifesta, M.; Viappiani, A.; Turroni, F.; Margolles, A.; Ruas-Madiedo, P.; et al. Modulation of the eps-ome transcription of bifidobacteria through simulation of human intestinal environment. FEMS Microbiol. Ecol. 2016, 92, fiw056. [Google Scholar] [CrossRef]
- Turroni, F.; Ventura, M.; Butto, L.F.; Duranti, S.; O’Toole, P.W.; Motherway, M.O.; van Sinderen, D. Molecular dialogue between the human gut microbiota and the host: A Lactobacillus and Bifidobacterium perspective. Cell. Mol. Life Sci. 2014, 71, 183–203. [Google Scholar] [CrossRef]
- Horn, N.; Wegmann, U.; Dertli, E.; Mulholland, F.; Collins, S.R.; Waldron, K.W.; Bongaerts, R.J.; Mayer, M.J.; Narbad, A. Spontaneous mutation reveals influence of exopolysaccharide on Lactobacillus johnsonii surface characteristics. PLoS ONE 2013, 8, e59957. [Google Scholar] [CrossRef]
- Fanning, S.; Hall, L.J.; van Sinderen, D. Bifidobacterium breve UCC2003 surface exopolysaccharide production is a beneficial trait mediating commensal-host interaction through immune modulation and pathogen protection. Gut Microbes 2012, 3, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Ruas-Madiedo, P.; Gueimonde, M.; Arigoni, F.; de los Reyes-Gavilan, C.G.; Margolles, A. Bile affects the synthesis of exopolysaccharides by Bifidobacterium animalis. Appl. Environ. Microbiol. 2009, 75, 1204–1207. [Google Scholar] [CrossRef] [PubMed]
- Fanning, S.; Hall, L.J.; Cronin, M.; Zomer, A.; MacSharry, J.; Goulding, D.; Motherway, M.O.; Shanahan, F.; Nally, K.; Dougan, G.; et al. Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc. Natl. Acad. Sci. USA 2012, 109, 2108–2113. [Google Scholar] [CrossRef] [PubMed]
- Altmann, F.; Kosma, P.; O’Callaghan, A.; Leahy, S.; Bottacini, F.; Molloy, E.; Plattner, S.; Schiavi, E.; Gleinser, M.; Groeger, D.; et al. Genome Analysis and Characterisation of the Exopolysaccharide Produced by Bifidobacterium longum subsp. longum 35624. PLoS ONE 2016, 11, e0162983. [Google Scholar] [CrossRef]
- Schiavi, E.; Gleinser, M.; Molloy, E.; Groeger, D.; Frei, R.; Ferstl, R.; Rodriguez-Perez, N.; Ziegler, M.; Grant, R.; Moriarty, T.F.; et al. The Surface-Associated Exopolysaccharide of Bifidobacterium longum 35624 Plays an Essential Role in Dampening Host Proinflammatory Responses and Repressing Local TH17 Responses. Appl. Environ. Microbiol. 2016, 82, 7185–7196. [Google Scholar] [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Ratajczak, W.; Ryl, A.; Mizerski, A.; Walczakiewicz, K.; Sipak, O.; Laszczynska, M. Immunomodulatory potential of gut microbiome-derived short-chain fatty acids (SCFAs). Acta Biochim. Pol. 2019, 66, 1–12. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de Los Reyes-Gavilan, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Riviere, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Riviere, A.; Gagnon, M.; Weckx, S.; Roy, D.; De Vuyst, L. Mutual Cross-Feeding Interactions between Bifidobacterium longum subsp. longum NCC2705 and Eubacterium rectale ATCC 33656 Explain the Bifidogenic and Butyrogenic Effects of Arabinoxylan Oligosaccharides. Appl. Environ. Microbiol. 2015, 81, 7767–7781. [Google Scholar] [CrossRef]
- Falony, G.; Calmeyn, T.; Leroy, F.; De Vuyst, L. Coculture fermentations of Bifidobacterium species and Bacteroides thetaiotaomicron reveal a mechanistic insight into the prebiotic effect of inulin-type fructans. Appl. Environ. Microbiol. 2009, 75, 2312–2319. [Google Scholar] [CrossRef] [PubMed]
- Rios-Covian, D.; Sanchez, B.; Salazar, N.; Martinez, N.; Redruello, B.; Gueimonde, M.; de Los Reyes-Gavilan, C.G. Different metabolic features of Bacteroides fragilis growing in the presence of glucose and exopolysaccharides of bifidobacteria. Front. Microbiol. 2015, 6, 825. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Gueimonde, M.; Duncan, S.H.; Flint, H.J.; de los Reyes-Gavilan, C.G. Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.; O’Connell Motherway, M.; Ventura, M.; van Sinderen, D. Metabolism of sialic acid by Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 2014, 80, 4414–4426. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Ozcan, E.; Milani, C.; Mancabelli, L.; Viappiani, A.; van Sinderen, D.; Sela, D.A.; Ventura, M. Glycan cross-feeding activities between bifidobacteria under in vitro conditions. Front. Microbiol. 2015, 6, 1030. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Milani, C.; Duranti, S.; Mancabelli, L.; Mangifesta, M.; Viappiani, A.; Lugli, G.A.; Ferrario, C.; Gioiosa, L.; Ferrarini, A.; et al. Deciphering bifidobacterial-mediated metabolic interactions and their impact on gut microbiota by a multi-omics approach. ISME J. 2016, 10, 1656–1668. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef]
- Moens, E.; Veldhoen, M. Epithelial barrier biology: Good fences make good neighbours. Immunology 2012, 135, 1–8. [Google Scholar] [CrossRef]
- Baumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Nunez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Tojo, R.; Suarez, A.; Clemente, M.G.; de los Reyes-Gavilan, C.G.; Margolles, A.; Gueimonde, M.; Ruas-Madiedo, P. Intestinal microbiota in health and disease: Role of bifidobacteria in gut homeostasis. World J. Gastroenterol. 2014, 20, 15163–15176. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Lewis, J.D. Analysis of the human gut microbiome and association with disease. Clin. Gastroenterol. Hepatol. 2013, 11, 774–777. [Google Scholar] [CrossRef] [PubMed]
- Mangifesta, M.; Mancabelli, L.; Milani, C.; Gaiani, F.; de’Angelis, N.; de’Angelis, G.L.; van Sinderen, D.; Ventura, M.; Turroni, F. Mucosal microbiota of intestinal polyps reveals putative biomarkers of colorectal cancer. Sci. Rep. 2018, 8, 13974. [Google Scholar] [CrossRef] [PubMed]
- Khailova, L.; Mount Patrick, S.K.; Arganbright, K.M.; Halpern, M.D.; Kinouchi, T.; Dvorak, B. Bifidobacterium bifidum reduces apoptosis in the intestinal epithelium in necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1118–G1127. [Google Scholar] [CrossRef]
- Bozzi Cionci, N.; Baffoni, L.; Gaggia, F.; Di Gioia, D. Therapeutic Microbiology: The Role of Bifidobacterium breve as Food Supplement for the Prevention/Treatment of Paediatric Diseases. Nutrients 2018, 10, 1723. [Google Scholar] [CrossRef] [PubMed]
- Braga, T.D.; da Silva, G.A.; de Lira, P.I.; de Carvalho Lima, M. Efficacy of Bifidobacterium breve and Lactobacillus casei oral supplementation on necrotizing enterocolitis in very-low-birth-weight preterm infants: A double-blind, randomized, controlled trial. Am. J. Clin. Nutr. 2011, 93, 81–86. [Google Scholar] [CrossRef]
- Duranti, S.; Gaiani, F.; Mancabelli, L.; Milani, C.; Grandi, A.; Bolchi, A.; Santoni, A.; Lugli, G.A.; Ferrario, C.; Mangifesta, M.; et al. Elucidating the gut microbiome of ulcerative colitis: Bifidobacteria as novel microbial biomarkers. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef]
- Leslie, A.; Carey, F.A.; Pratt, N.R.; Steele, R.J. The colorectal adenoma-carcinoma sequence. Br. J. Surg. 2002, 89, 845–860. [Google Scholar] [CrossRef]


{kind=link}
| Bifidobacterium Strains | Isolation | References |
|---|---|---|
| B. actinocoloniiforme DSM 22766 | Bumblebee digestive tract | [16] |
| B. adolescentis ATCC 15703 | Intestine of human adult | [17] |
| B. aemilianum XV10 | Carpenter bee digestive tract | [5] |
| B. aerophilum DSM 100689 | Feces of cotton-top tamarin | [18] |
| B. aesuclapii DSM 26737 | Feces of baby common marmoset | [19] |
| B. angulatum LMG 11039 | Feces of human | [20] |
| B. animalis subsp. animalis LMG 10508 | Feces of rat | [21] |
| B. animalis subsp. lactis DSM 10140 | Fermented milk | [22] |
| B. anseris LMG 30189 | Feces of domestic goose | [7] |
| B. apri DSM 100238 | Digestive tract of wild pig | [23] |
| B. aquikefiri LMG 28769 | Water kefir | [24] |
| B. asteroides LMG 10735 | Hindgut of honeybee | [25] |
| B. avesanii DSM 100685 | Feces of cotton-top tamarin | [18] |
| B. biavatii DSM 23969 | Feces of tamarin | [26] |
| B. bifidum LMG 11041 | Feces of breast-fed infant | [3] |
| B. bohemicum DSM22767 | Bumblebee digestive tract | [16] |
| B. bombi DSM 19703 | Bumblebee digestive tract | [27] |
| B. boum LMG 10736 | Rumen of bovine | [28] |
| B. breve LMG 13208 | Infant stool | [17] |
| B. callimiconis LMG 30938 | Feces of Goeldi’s marmoset | [6] |
| B. callitrichidarum DSM 103152 | Feces of emperor tamarin | [29] |
| B. callitrichos DSM 23973 | Feces of common marmoset | [26] |
| B. canis DSM105923 | Feces of dog | [10] |
| B. castoris LMG 30937 | Feces of beaver | [6] |
| B. catenulatum LMG 11043 | Adult intestine | [30] |
| B. catenulatum subsp. kashiwanohense DSM21854 | Infant feces | [31] |
| B. catulorum DSM103154 | Feces of common marmoset | [32] |
| B. cebidarum LMG31469 | Feces of golden-headed tamarin | [9] |
| B. choerinum LMG 10510 | Feces of piglet | [28] |
| B. commune LMG28292 | Bumblebee gut | [33] |
| B. coryneforme LMG 18911 | Hindgut of honeybee | [25] |
| B. criceti LMG 30188 | Feces of European hamster | [7] |
| B. crudilactis LMG 23609 | Raw cow milk | [34] |
| B. cuniculi LMG 10738 | Feces of rabbit | [28] |
| B. dentium LMG 11045 | Oral cavity | [30] |
| B. dolichotidis LMG 30941 | Feces of Patagonian mara | [6] |
| B. eulemuris DSM 100216 | Feces of black lemur | [35] |
| B. felsineum DSM103139 | Feces of cotton-top tamarin | [11] |
| B. gallicum LMG 11596 | Adult intestine | [36] |
| B. goeldii LMG 30939 | Feces of Goeldi’s marmoset | [6] |
| B. hapali DSM 100202 | Feces of baby common marmoset | [37] |
| B. imperatoris LMG 30297 | Feces of emperor tamarin | [7] |
| B. indicum LMG 11587 | Insect | [25] |
| B. italicum LMG 30187 | Feces of European rabbit | [7] |
| B. jacchi DSM 103362 | Feces of baby common marmoset | [38] |
| B. lemurum DSM 28807 | Feces of ring-tailed lemur | [39] |
| B. leontopitechi LMG 31471 | Feces of Goeldi’s monkey | [9] |
| B. longum subsp. infantis ATCC 15697 | Intestine of infant | [17] |
| B. longum subsp. longum LMG 13197 | Adult intestine | [17] |
| B. longum subsp. suis LMG 21814 | Feces of pig | [40] |
| B. magnum LMG 11591 | Feces of rabbit | [30] |
| B. margollesii LMG 30296 | Feces of pygmy marmoset | [7] |
| B. meryciucm LMG 11341 | Rumen of bovine | [41] |
| B. minimum LMG 11592 | Sewage | [42] |
| B. mongoliense DSM 21395 | Fermented mare’s milk | [43] |
| B. moukabalense DSM 27321 | Feces of gorilla | [44] |
| B. myosotis DSM 100196 | Feces of common marmoset | [37] |
| B. parmae LMG 30295 | Feces of pygmy marmoset | [7] |
| B. platyrrhinorum SMA15 | Feces of squirrel monkey | [45] |
| B. primatium DSM 100687 | Feces of cotton-top tamarin | [11] |
| B. pseudocatenulatum LMG 10505 | Infant feces | [28] |
| B. pseudolongum subsp. globosum LMG 11596 | Rumen of bovine | [46] |
| B. pseudolongum subsp. pseudolongum LMG 11571 | Feces of swine | [21] |
| B. psychraerophilum LMG 21775 | Caecum of pig | [47] |
| B. pullorum subsp. gallinarum LMG 11586 | Caecum of chicken | [48] |
| B. pullorum subsp. pullorum LMG 21816 | Feces of chicken | [8] |
| B. ramosum DSM 100688 | Feces of cotton-top tamarin | [18] |
| B. reuteri DSM 23975 | Feces of common marmoset | [26] |
| B. rousetti BCRC 81136 | Feces of Egyptian fruit bat | [49] |
| B. ruminantium LMG 21811 | Rumen of bovine | [41] |
| B. pullorum subsp. saeculare LMG 14934 | Feces of rabbit | [50] |
| B. saguini LMG 23967 | Feces of tamarin | [26] |
| B. saimiriisciurei SMA1 | Feces of squirrel monkey | [45] |
| B. saimirii LMG 30940 | Feces of Bolivian saimiri | [6] |
| B. scaligerum DSM 103140 | Feces of cotton-top tamarin | [11] |
| B. scardovii LMG 21589 | Blood | [51] |
| B. simiarum DSM 103153 | Feces of emperor tamarin | [11] |
| B. stellenboschense DSM 23968 | Feces of tamarin | [26] |
| B. subtile LMG 11597 | Sewage | [42] |
| B. porcinum LMG 21689 | Feces of piglet | [52] |
| B. thermacidophilum LMG 21395 | Anaerobic digester | [53] |
| B. termophilum JCM 7027 | Rumen of bovine | [21] |
| B. tibiigranuli LMG 31086 | Water kefir | [54] |
| B. tissieri DSM 100201 | Feces of baby common marmoset | [37] |
| B. tsurumiense JCM 13495 | Hamster dental plaque | [55] |
| B. vansinderenii LMG 30126 | Feces of emperor tamarin | [56] |
| B. vespertilionis DSM 106025 | Feces of Egyptian fruit bat | [49] |
| B. xylocopae DSM104955 | Carpenter bee digestive tract | [5] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duranti, S.; Longhi, G.; Ventura, M.; van Sinderen, D.; Turroni, F. Exploring the Ecology of Bifidobacteria and Their Genetic Adaptation to the Mammalian Gut. Microorganisms 2021, 9, 8. https://doi.org/10.3390/microorganisms9010008
Duranti S, Longhi G, Ventura M, van Sinderen D, Turroni F. Exploring the Ecology of Bifidobacteria and Their Genetic Adaptation to the Mammalian Gut. Microorganisms. 2021; 9(1):8. https://doi.org/10.3390/microorganisms9010008
Chicago/Turabian StyleDuranti, Sabrina, Giulia Longhi, Marco Ventura, Douwe van Sinderen, and Francesca Turroni. 2021. "Exploring the Ecology of Bifidobacteria and Their Genetic Adaptation to the Mammalian Gut" Microorganisms 9, no. 1: 8. https://doi.org/10.3390/microorganisms9010008
APA StyleDuranti, S., Longhi, G., Ventura, M., van Sinderen, D., & Turroni, F. (2021). Exploring the Ecology of Bifidobacteria and Their Genetic Adaptation to the Mammalian Gut. Microorganisms, 9(1), 8. https://doi.org/10.3390/microorganisms9010008