Taxonomic Characterization, and Secondary Metabolite Analysis of Streptomyces triticiradicis sp. nov.: A Novel Actinomycete with Antifungal Activity

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Actinobacterial Strain
2.2. Morphological and Biochemical Characteristics
2.3. Chemotaxonomic Analysis
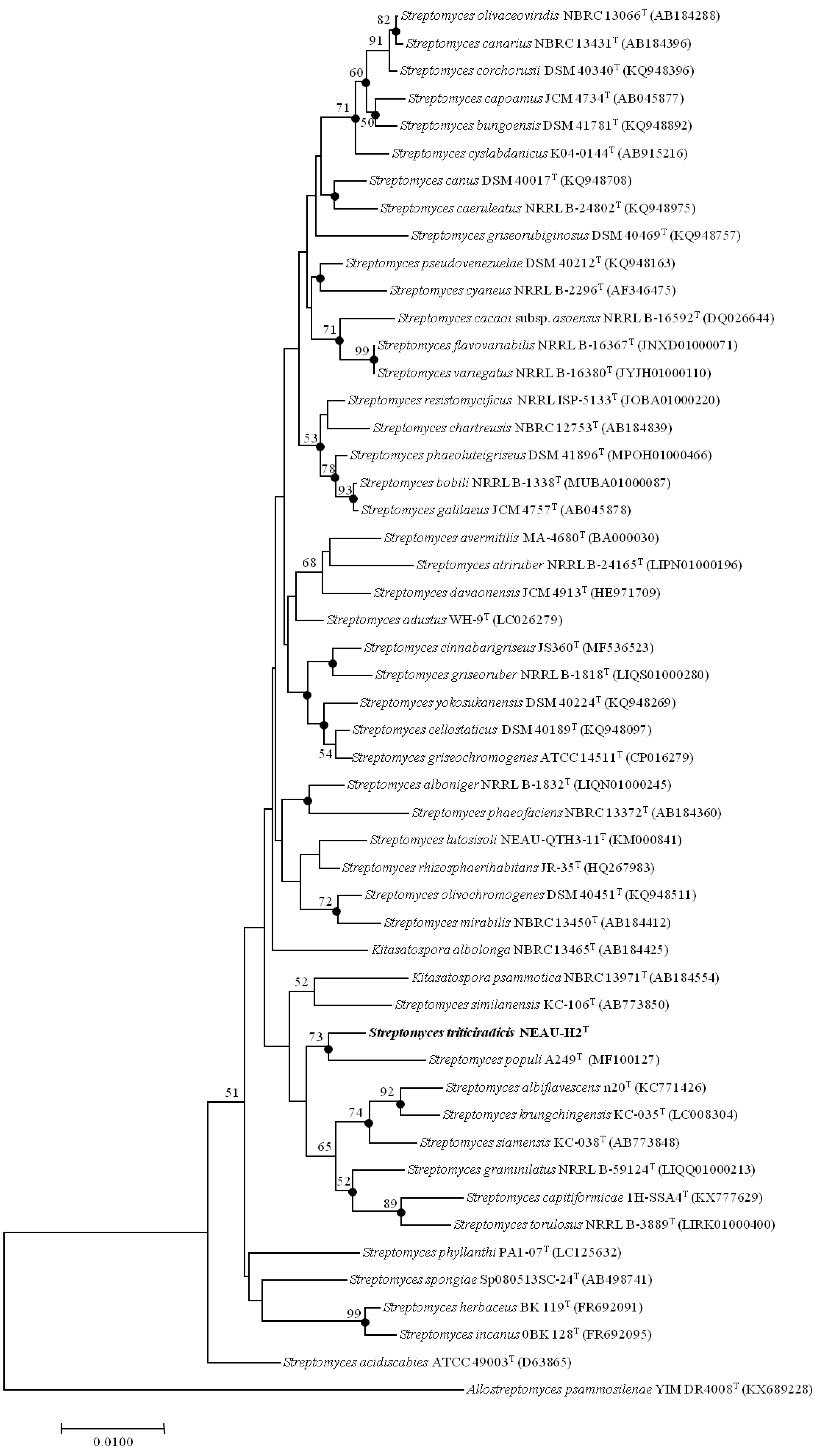
2.4. Phylogenetic Analysis
2.5. DNA–DNA Relatedness Tests
2.6. In Vitro Antifungal Activity Test
2.7. Isolation and Characterization of Secondary Metabolites
2.8. In Vitro Antifungal Activity Test of Compounds
3. Results and Discussion
3.1. Polyphasic Taxonomic Characterization of NEAU-H2T
3.2. Description of Streptomyces triticiradicis sp. nov.
3.3. Antifungal Activity Evaluation
3.4. Identified of Secondary Metabolites from Strain NEAU-H2T
3.5. Mining the Biosynthetic Potential of the Strain
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Volova, T.; Prudnikova, S.; Boyandin, A.; Zhila, N.; Kiselev, E.; Shumilova, A.; Baranovskiy, S.; Demidenko, A.; Shishatskaya, A.; Thomas, S. Constructing slow-release fungicide formulations based on poly (3-hydroxybutyrate) and natural materials as a degradable matrix. J. Agric. Food Chem. 2019, 67, 9220–9231. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.B.; Gao, Y.Q.; Nie, X.D.; Tuong, T.M.; Li, D.; Gao, J.M. Antifungal activity of griseofulvin derivatives against phytopathogenic fungi in vitro and in vivo and three-dimensional quantitative structure-activity relationship analysis. J. Agric. Food Chem. 2019, 67, 6125–6132. [Google Scholar] [CrossRef] [PubMed]
- Onaka, H. Novel antibiotic screening methods to awaken silent or cryptic secondary metabolic pathways in actinomycetes. J. Antibiot. 2017, 70, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, R.; Li, Y.A.; Li, S.Y.; Yu, J.; Zhao, B.F.; Liao, A.C.; Wang, Y.; Wang, Z.W.; Lu, A.D.; et al. Discovery of pimprinine alkaloids as novel agents against a plant virus. J. Agric. Food Chem. 2019, 67, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, D.; Qi, D.; Gao, Z.; Xie, J.; Luo, Y. Growth promotion and disease suppression ability of a Streptomyces sp. CB-75 from Banana rhizosphere soil. Front. Microbiol. 2018, 8, 2704. [Google Scholar] [CrossRef] [PubMed]
- Kemung, H.M.; Tan, L.T.; Khan, T.M.; Chan, K.G.; Pusparajah, P.; Goh, B.H.; Lee, L.H. Streptomyces as a prominent resource of future anti-MRSA drugs. Front. Microbiol. 2018, 9, 2221. [Google Scholar] [CrossRef] [PubMed]
- Janardhan, A.; Kumar, A.P.; Viswanath, B.; Saigopal, D.V.; Narasimha, G. Production of bioactive compounds by Actinomycetes and their antioxidant properties. Biotechnol. Res. Int. 2014, 2014, 217030. [Google Scholar] [CrossRef]
- Minuto, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Control of soilborne pathogens of tomato using a commercial formulation of Streptomyces griseoviridis and solarization. Crop Protect. 2006, 25, 468–475. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant. Soil. 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Che, Q.; Zhu, T.J.; Keyzers, R.A.; Liu, X.F.; Li, J.; Gu, Q.Q.; Li, D.H. Polycyclic hybrid isoprenoids from a reed rhizosphere soil derived Streptomyces sp. CHQ-64. J. Nat. Prod. 2013, 76, 759–763. [Google Scholar] [CrossRef]
- Zhao, K.; Penttinen, P.; Chen, Q.; Guan, T.W.; Lindström, K.; Ao, X.L.; Zhang, L.L.; Zhang, X.P. The rhizospheres of traditional medicinal plants in Panxi, China, host a diverse selection of actinobacteria with antimicrobial properties. Appl. Microbiol. Biotechnol. 2012, 94, 1321–1335. [Google Scholar] [CrossRef]
- Zhao, J.W.; Shi, L.L.; Li, W.C.; Wang, J.B.; Wang, H.; Tian, Y.Y.; Xiang, W.S.; Wang, X.J. Streptomyces tritici sp. nov. a novel actinomycete isolated from rhizosphere soil of wheat (Triticum aestivum L.). Int. J. Syst. Evol. Microbiol. 2018, 68, 492–497. [Google Scholar] [CrossRef]
- Shirling, E.B.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef]
- Smibert, R.M.; Krieg, N.R. Phenotypic Characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Jin, L.Y.; Zhao, Y.; Song, W.; Duan, L.P.; Jiang, S.W.; Wang, X.J.; Zhao, J.W.; Xiang, W.S. Streptomyces inhibens sp. nov., a novel actinomycete isolated from rhizosphere soil of wheat (Triticum aestivum L.). Int. J. Syst. Evol. Microbiol. 2019, 69, 688–695. [Google Scholar] [CrossRef]
- Jones, K.L. Fresh isolates of actinomycetes in which the presence of sporogenous aerial mycelia is a fluctuating characteristic. J. Bacteriol. 1949, 57, 141–145. [Google Scholar] [CrossRef]
- Waksman, S.A. The Actinomycetes. A Summary of Current Knowledge; The Ronald Press Co.: New York, NY, USA, 1967; p. 286. [Google Scholar]
- Waksman, S.A. The Actinomycetes, Volume 2, Classification, Identification and Descriptions of Genera and Species; Williams and Wilkins Company: Philadelphia, PA, USA, 1961; p. 363. [Google Scholar]
- Kelly, K.L. Color-Name Charts Illustrated with Centroid Colors. In Inter-Society Color Council-National Bureau of Standards; U.S. National Bureau of Standards: Washington, DC, USA, 1965. [Google Scholar]
- Jia, F.Y.; Liu, C.X.; Wang, X.J.; Zhao, J.W.; Liu, Q.F.; Zhang, J.; Gao, R.X.; Xiang, W.S. Wangella harbinensis gen. nov., sp. nov., a new member of the family Micromonosporaceae. Antonie Leeuwenhoek 2013, 103, 399–408. [Google Scholar] [CrossRef]
- Zhao, J.W.; Han, L.Y.; Yu, M.Y.; Cao, P.; Li, D.; Guo, X.W.; Liu, Y.Q.; Wang, X.J.; Xiang, W.S. Characterization of Streptomyces sporangiiformans sp. nov., a novel soil actinomycete with antibacterial activity against Ralstonia solanacearum. Microorganisms 2019, 7, 360–376. [Google Scholar]
- Gordon, R.E.; Barnett, D.A.; Handerhan, J.E.; Pang, C. Nocardia coeliaca, Nocardia autotrophica, and the nocardin strain. Int. J. Syst. Bacteriol. 1974, 24, 54–63. [Google Scholar] [CrossRef]
- Yokota, A.; Tamura, T.; Hasegawa, T.; Huang, L.H. Catenuloplanes japonicas gen. nov., sp. nov., nom. rev., a new genus of the order actinomycetales. Int. J. Syst. Bacteriol. 1993, 43, 805–812. [Google Scholar] [CrossRef]
- McKerrow, J.; Vagg, S.; McKinney, T.; Seviour, E.M.; Maszenan, A.M.; Brooks, P.; Sevious, R.J. A simple HPLC method for analysing diaminopimelic acid diastereomers in cell walls of Gram-positive bacteria. Lett. Appl. Microbiol. 2000, 30, 178–182. [Google Scholar] [CrossRef]
- Lechevalier, M.P.; Lechevalier, H.A. The Chemotaxonomy of Actinomycetes. In Actinomycete Taxonomy; Dietz, A., Thayer, D.W., Eds.; Special Publication for Society of Industrial Microbiology: Arlington, TX, USA, 1980; pp. 227–291. [Google Scholar]
- Minnikin, D.E.; O’Donnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J.H. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Methods. 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Collins, M.D. Isoprenoid Quinone Analyses in Bacterial Classification and Identification. In Chemical Methods in Bacterial Systematics; Goodfellow, M., Minnikin, D.E., Eds.; Academic Press: Cambridge, MA, USA, 1985; pp. 267–284. [Google Scholar]
- Wu, C.; Lu, X.; Qin, M.; Wang, Y.; Ruan, J. Analysis of menaquinone compound in microbial cells by HPLC. Microbiology 1989, 16, 76–178. [Google Scholar]
- Xiang, W.S.; Liu, C.X.; Wang, X.J.; Du, J.; Xi, L.J.; Huang, Y. Actinoalloteichus nanshanensis sp. nov., isolated from the rhizosphere of a fig tree (Ficus religiosa). Int. J. Syst. Evol. Microbiol. 2011, 61, 1165–1169. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Bruns, M.A.; Tiedje, J.M. DNA recovery from soils of diverse composition. Appl. Environ. Microbiol. 1996, 62, 316–322. [Google Scholar]
- Yi, R.K.; Tan, F.; Liao, W.; Wang, Q.; Mu, J.F.; Zhou, X.R.; Yang, Z.N.; Zhao, X. Isolation and identification of Lactobacillus plantarum HFY05 from natural fermented Yak Yogurt and Its effect on alcoholic liver injury in mice. Microorganisms 2019, 7, 530. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 83–791. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Q.; Zhu, H.M.; Ruan, J.; Qian, W.B.; Fang, X.D.; Shi, Z.B.; Li, Y.R.; Li, S.T.; Shan, G.; Kristiansen, K.; et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef]
- Ley, J.D.; Cattoir, H.; Reynaerts, A. The quantitative measurement of DNA hybridization from renaturation rates. Eur. J. Biochem. 1970, 12, 133–142. [Google Scholar] [CrossRef]
- Huss, V.A.R.; Festl, H.; Schleifer, K.H. Studies on the spectrometric determination of DNA hybridisation from renaturation rates. Syst. Appl. Microbiol. 1983, 4, 184–192. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Goker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Duran, H.G.S.; Santos, E.L.C.D.L.; Kim, H.U.; Nave, M.; et al. Antismash 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef]
- Liu, C.X.; Zhuang, X.X.; Yu, Z.Y.; Wang, Z.Y.; Wang, Y.J.; Guo, X.W.; Xiang, W.S.; Huang, S.X. Community structures and antifungal activity of root-associated endophytic actinobacteria of healthy and diseased soybean. Microorganisms 2019, 7, 243. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.J.; Yan, Y.J.; Jiang, L.; Wang, J.D.; Li, B.J.; Xiang, W.S. Isolation and identification of 5-hydroxyl-5-methyl-2-hexenoic acid from Actinoplanes sp. HBDN08 with antifungal activity. Bioresour. Technol. 2010, 101, 8383–8388. [Google Scholar] [CrossRef]
- Yu, Z.Y.; Wang, L.; Yang, J.; Zhang, F.; Sun, Y.; Yu, M.M.; Yan, Y.J.; Ma, Y.T.; Huang, S.X. A new antifungal macrolide from Streptomyces sp. KIB-H869 and structure revision of halichomycin. Tetrahedron Lett. 2016, 57, 1375–1378. [Google Scholar] [CrossRef]
- Kämpfer, P.; Genus, I. Streptomyces Waksman and Henrici 1943, 339 AL. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Springer: New York, NY, USA, 2012; pp. 1679–1680. [Google Scholar]
- Lee, H.J.; Whang, K.S. Streptomyces rhizosphaerihabitans sp. nov. and Streptomyces adustus sp. nov. isolated from bamboo forest soil. Int. J. Syst. Microbiol. 2016, 66, 3573–3578. [Google Scholar]
- Wang, Z.K.; Jiang, B.J.; Li, X.G.; Gan, L.Z.; Long, X.F.; Zhang, Y.Q.; Tian, Y.Q. Streptomyces populi sp. nov. a novel endophytic actinobacterium isolated from stem of Populus adenopoda Maxim. Int. J. Syst. Microbiol. 2018, 68, 2568–2573. [Google Scholar]
- Sripreechasak, P.; Matsumoto, A.; Suwanborirux, K.; Inahashi, Y.; Shiomi, K.; Tanasupawat, S.; Takahashi, Y. Streptomyces siamensis sp. nov., and Streptomyces similanensis sp. nov., isolated from Thai soils. J. Antibiot. 2013, 66, 633–640. [Google Scholar]
- Kim, S.B.; Lonsdale, J.; Seong, C.N.; Goodfellow, M. Streptacidiphilus gen. nov., acidophilic actinomycetes with wall chemotype I and emendation of the family Streptomycetaceae (Waksman and Henrici (1943) AL) emend. Rainey et al. 1997. Antonie. Leeuwenhoek. 2003, 83, 107–116. [Google Scholar] [CrossRef]
- Kroppenstedt, R.M. Fatty Acid and Menaquinone Analysis of Actinomycetes and Related Organisms. In Chemical Methods in Bacterial Systematics; Goodfellow, M., Minnikin, D.E., Eds.; Academic Press: London, UK, 1985; pp. 173–199. [Google Scholar]
- Lechevalier, M.P.; Lechevalier, H. Chemical composition as a criterion in the classification of aerobic actinomycetes. Int. J. Syst. Bacteriol. 1970, 20, 435–443. [Google Scholar] [CrossRef]
- Rong, X.Y.; Huang, Y. Taxonomic evaluation of the Streptomyces hygroscopicus clade using multilocus sequence analysis and DNA-DNA hybridization, validating the MLSA scheme for systematics of the whole genus. Syst. Appl. Microbiol. 2012, 35, 7–18. [Google Scholar] [CrossRef]
- Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; Starr, M.P.; Trper, H.G. International Committee on Systematic Bacteriology. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 1987, 37, 463–464. [Google Scholar]
- Ye, X.W.; Chai, W.Y.; Lian, X.Y.; Zhang, Z.Z. Novel propanamide analogue and antiproliferative diketopiperazines from mangrove Streptomyces sp. Q 24. Nat. Prod. Res. 2017, 31, 1390–1396. [Google Scholar] [CrossRef]
- Domondon, D.L.; He, W.; De Kimpe, N.; Höfte, M.; Poppe, J. β-Adenosine, a bioactive compound in grass chaff stimulating mushroom production. Phytochemistry 2004, 65, 181–187. [Google Scholar] [CrossRef]
- Cheng, Y.X.; Zhou, J.; Teng, R.W.; Tan, N.H. Nitrogen-containing compounds from Brachystemma calycinum. Acta Bot. Yunnanica 2001, 23, 527–530. [Google Scholar]
- Katsuyama, Y. Mining novel biosynthetic machineries of secondary metabolites from actinobacteria. Biosci. Biotechnol. Biochem. 2019, 83, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.P.; Yuan, P.H.; Wang, Y.Y.; Liu, X.F.; Zhou, Z.X.; Bu, Q.T.; Yu, P.; Jiang, H.; Li, Y.Q. Generation of the natamycin analogs by gene engineering of natamycin biosynthetic genes in Streptomyces chattanoogensis L10. Microbiol. Res. 2015, 173, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.L.; Li, S.Z.; Zhou, Z.; Chen, S.F.; Fan, W.M.; Li, Y.Q. The pleitropic regulator AdpAch is required for natamycin biosynthesis and morphological differentiation in Streptomyces chattanoogensis. Microbiology 2011, 157, 1300–1311. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Spore chain | Spiral | Straight ‡ | Straight † | Spiral § |
| Spore surface | Spiny | Hairy ‡ | Rough † | Smooth § |
| Growth temperature range (°C) | 4–40 | 10–40 | 10–37 | 10–40 |
| Growth pH range | 5–10 | 5–11 | 6–12 | 5–11 |
| NaCl tolerance range (w/v, %) | 0–9.0 | 0–10.0 | 0–4.0 | 0–10.0 |
| Cellulose decomposition | + | + | – | – |
| Gelatin liquefaction | – | + | + | – |
| Catalase production | – | + | + | – |
| H2S production | + | – | + | + |
| Milk coagulation | + | + | + | – |
| Nitrate reduction | – | – | + | – |
| Starch hydrolysis | + | – | – | – |
| Tween 20 hydrolysis | + | + | + | – |
| Tween 80 hydrolysis | + | + | – | + |
| Nitrogen source utilization | ||||
| l-serine | – | + | + | + |
| l-threonine | + | + | – | + |
| l-tyrosine | – | + | + | + |
| Carbon source utilization | ||||
| l-arabinose | – | + | + | + |
| Dulcitol | – | – | – | + |
| meso-inositol | – | + | + | + |
| d-mannitol | – | + | – | + |
| l-rhamnose | + | – | + | – |
| d-ribose | – | + | – | + |
| d-sorbitol | – | + | – | + |
| d-xylose | – | + | + | – |
| Phospholipids * | DPG, PE, PL, PI, PIM | AL, DPG, GL, PE, PG, PI, PIM, 2PLs ‡ | AL, APL, DPG, 2Ls, PE, PL, PIM † | DPG, PE, PG, PI, PL § |
| Menaquinones | MK-9(H6), MK-9(H8), MK-9(H4) | MK-9(H6), MK-9(H8) ‡ | MK-9(H6), MK-9(H8), MK-9(H2), MK-9(H4) † | MK-9(H6), MK-9(H4), MK-9(H8) § |
| Whole cell-wall sugars | Glucose | Glucose, ribose ‡ | Xylose, galactose † | ND |
| No. | δC | δH (mult, J in Hz) | 1H-1H COSY | HMBC (H→C) |
|---|---|---|---|---|
| 2 | 135.8 | 8.29 (s, 1H) | C-3, 3a, 7a, 8 | |
| 3 | 116.3 | |||
| 3a | 127.3 | |||
| 4 | 122.9 | 8.27 (d, J = 7.3 Hz, 1H) | H-5 | C-6 |
| 5 | 123.4 | 7.21 (td, J = 7.2, 1.2 Hz, 1H) | H-5, H-6 | C-4, 6, 3a, 7 |
| 6 | 124.5 | 7.24 (td, J = 7.2, 1.2 Hz, 1H) | H-5, H-7 | C-4, 7a |
| 7 | 112.9 | 7.46 (d, J = 7.4 Hz, 1H) | H-6 | C-5, 3a |
| 7a | 138.2 | |||
| 8 | 197.2 | |||
| 9 | 79.7 | 4.74 (d, J = 4.8 Hz, 1H) | H-10 | C-11, 10, 8 |
| 10 | 71.1 | 4.10 (m, 1H) | H-11, H-9 | C-11, 8 |
| 11 | 17.9 | 1.16 (d, J = 6.4 Hz, 3H) | H-10 | C-10, 9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Han, C.; Yu, B.; Zhao, J.; Yan, Y.; Huang, S.; Liu, C.; Xiang, W. Taxonomic Characterization, and Secondary Metabolite Analysis of Streptomyces triticiradicis sp. nov.: A Novel Actinomycete with Antifungal Activity. Microorganisms 2020, 8, 77. https://doi.org/10.3390/microorganisms8010077
Yu Z, Han C, Yu B, Zhao J, Yan Y, Huang S, Liu C, Xiang W. Taxonomic Characterization, and Secondary Metabolite Analysis of Streptomyces triticiradicis sp. nov.: A Novel Actinomycete with Antifungal Activity. Microorganisms. 2020; 8(1):77. https://doi.org/10.3390/microorganisms8010077
Chicago/Turabian StyleYu, Zhiyin, Chuanyu Han, Bing Yu, Junwei Zhao, Yijun Yan, Shengxiong Huang, Chongxi Liu, and Wensheng Xiang. 2020. "Taxonomic Characterization, and Secondary Metabolite Analysis of Streptomyces triticiradicis sp. nov.: A Novel Actinomycete with Antifungal Activity" Microorganisms 8, no. 1: 77. https://doi.org/10.3390/microorganisms8010077
APA StyleYu, Z., Han, C., Yu, B., Zhao, J., Yan, Y., Huang, S., Liu, C., & Xiang, W. (2020). Taxonomic Characterization, and Secondary Metabolite Analysis of Streptomyces triticiradicis sp. nov.: A Novel Actinomycete with Antifungal Activity. Microorganisms, 8(1), 77. https://doi.org/10.3390/microorganisms8010077