Zika Virus Infection in Pregnancy: Advanced Diagnostic Approaches in Dengue-Naive and Dengue-Experienced Pregnant Women and Possible Implication for Cross-Reactivity and Cross-Protection

 ,
, 
Abstract
:1. Introduction
2. Patients and Methods
3. Results
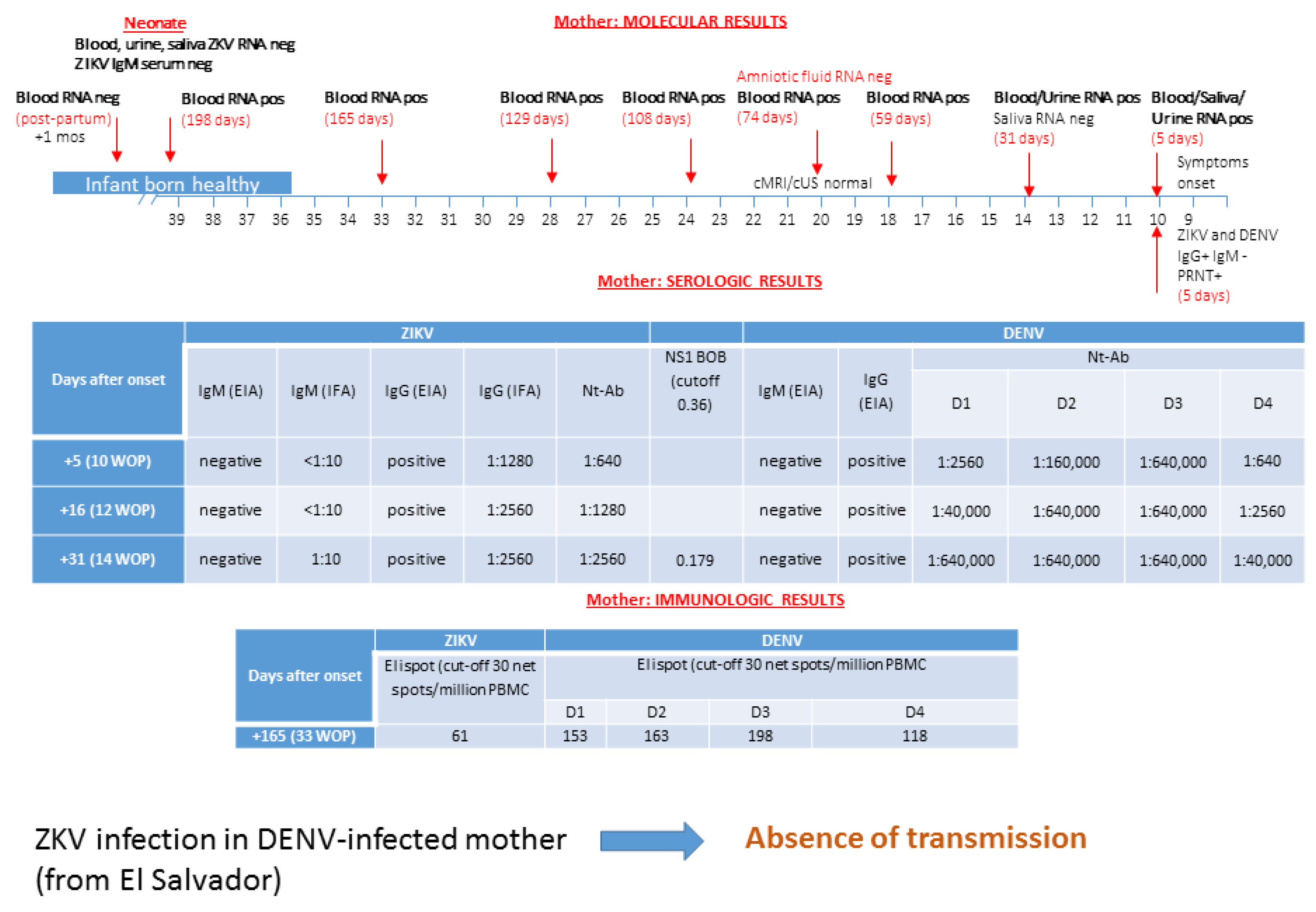
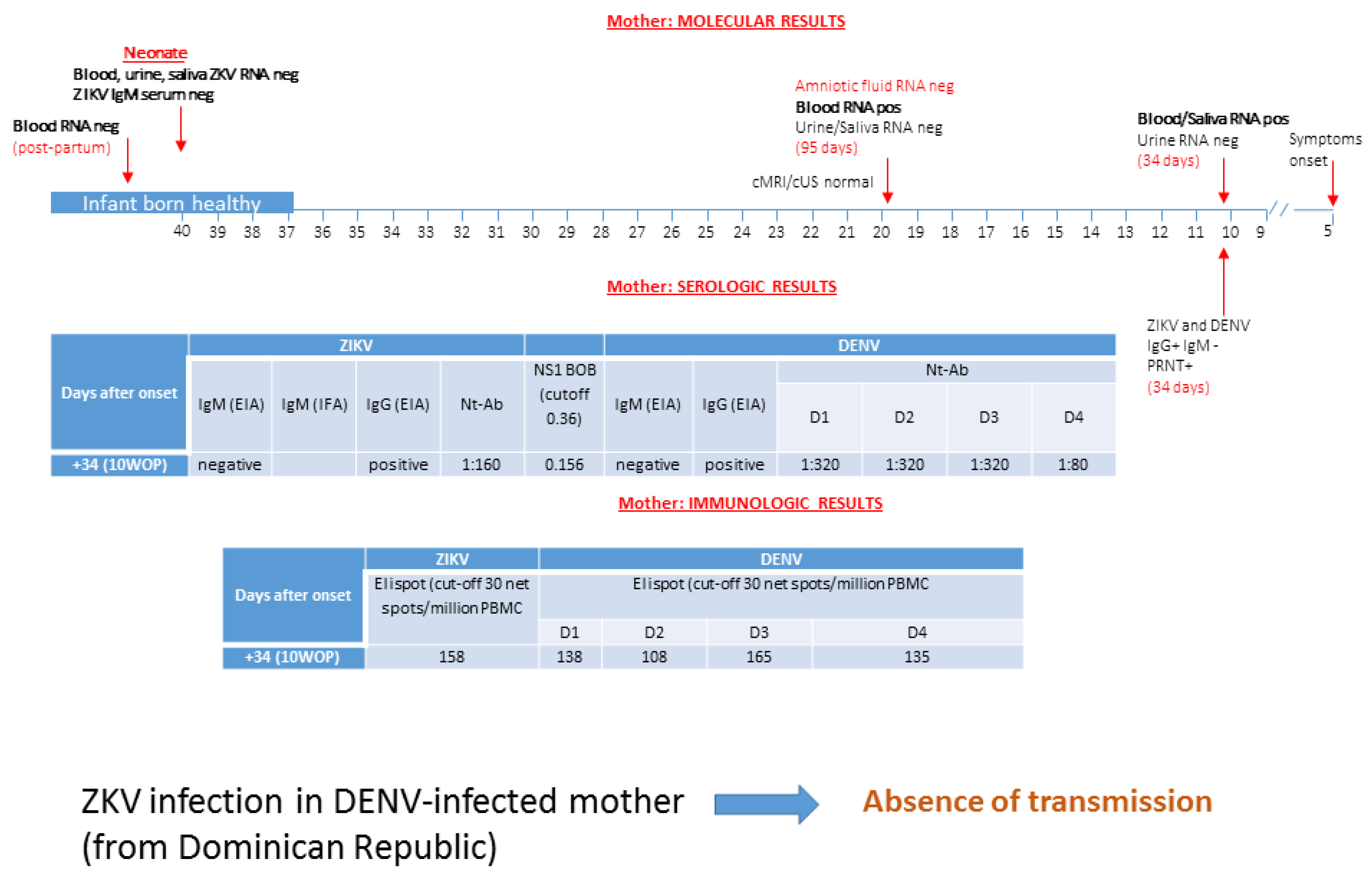
3.1. Patient 1: Absence of ZIKV Transmission in DENV-Experienced Mother
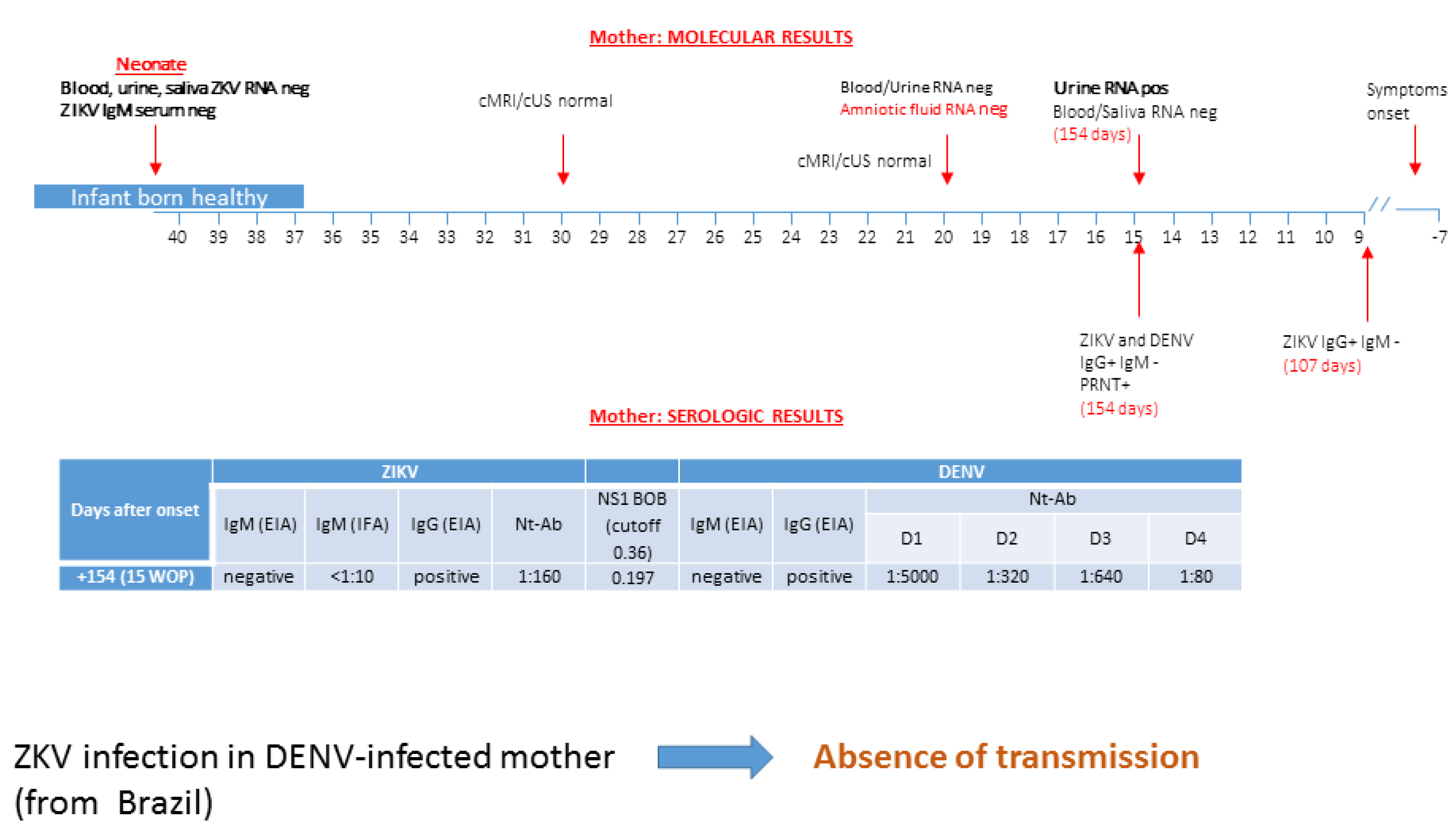
3.2. Patient 2: Absence of ZIKV Transmission in DENV-Experienced Mother
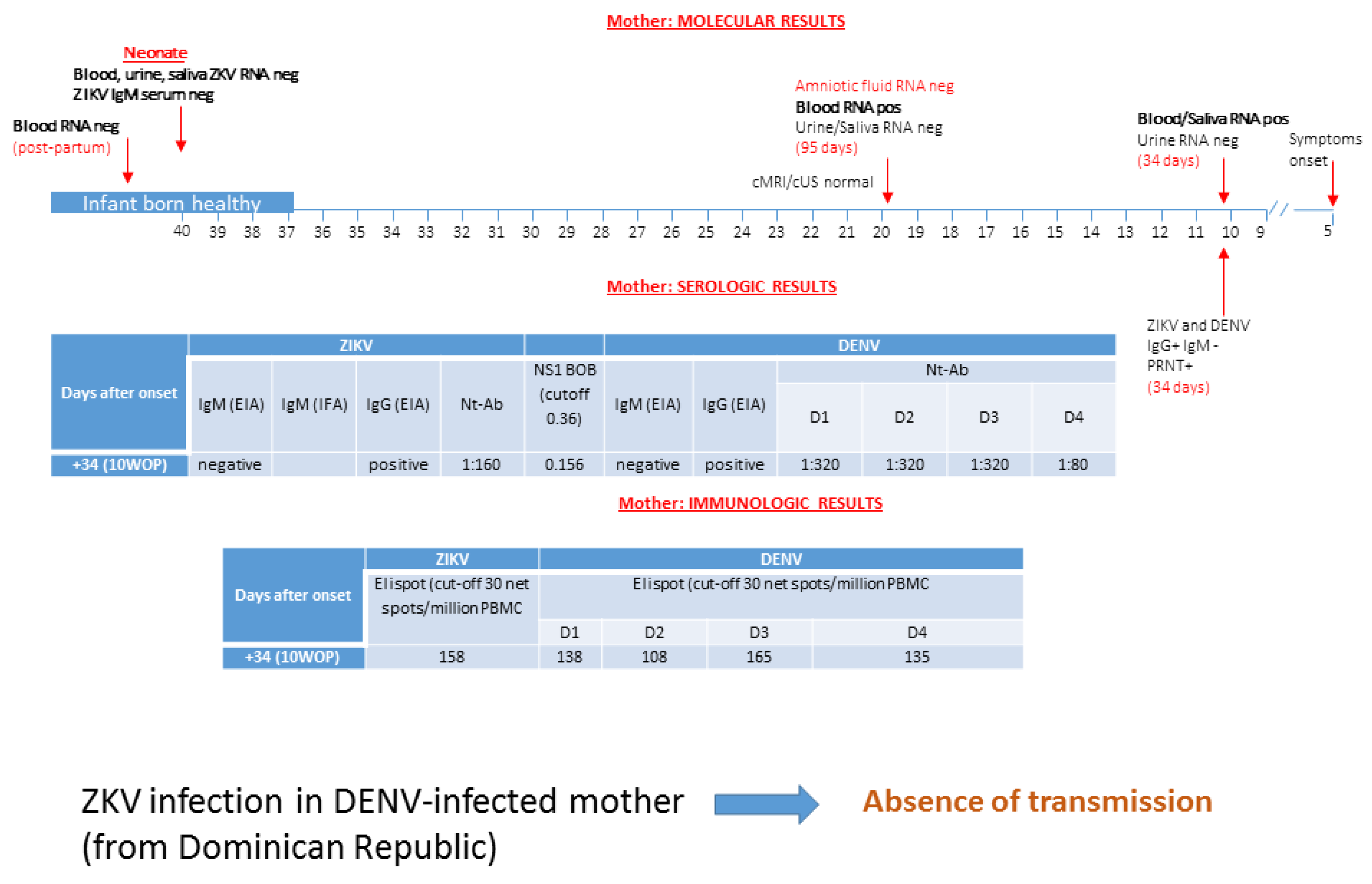
3.3. Patient 3: Absence of ZIKV Transmission in DENV-Experienced Mother
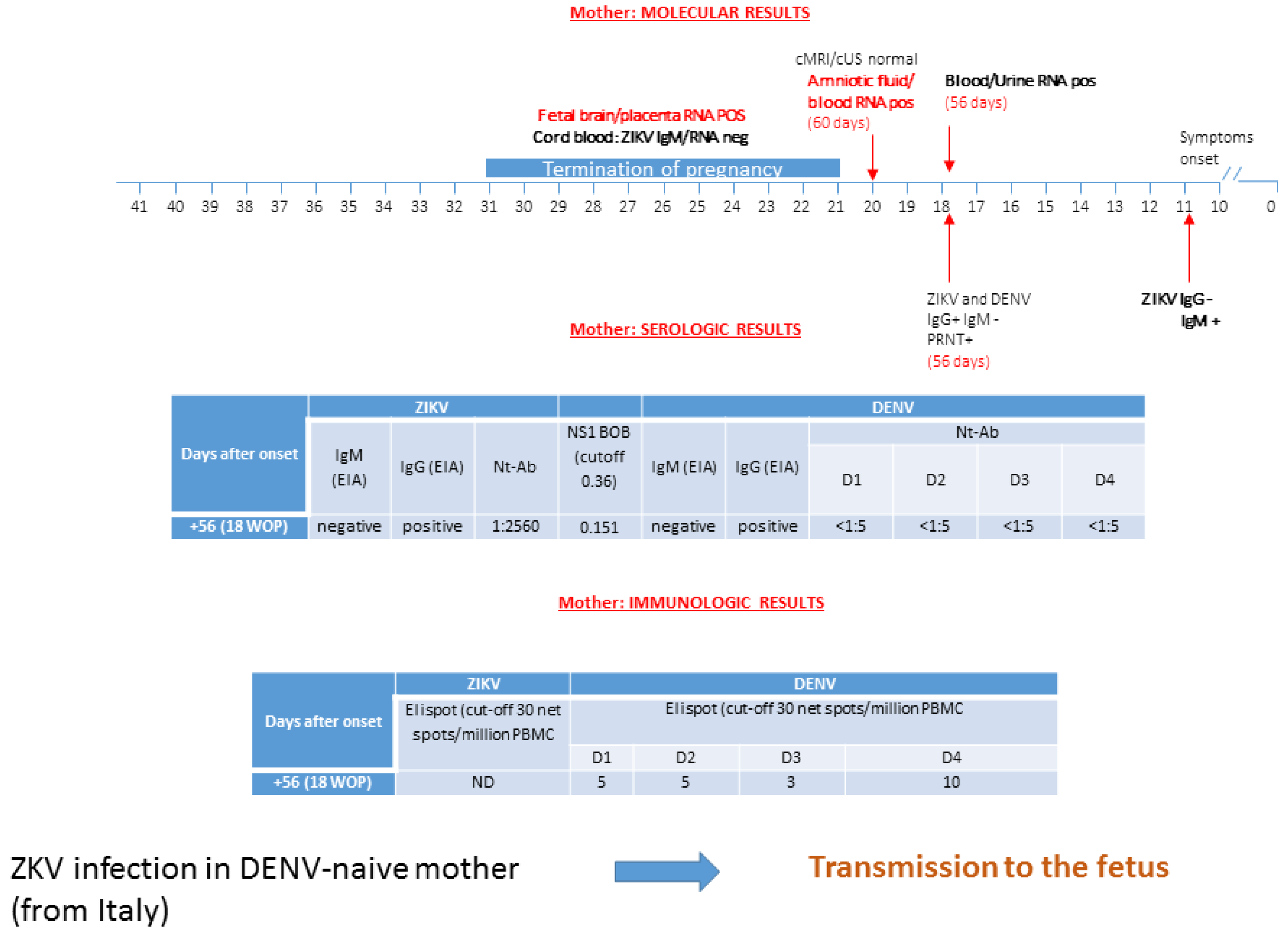
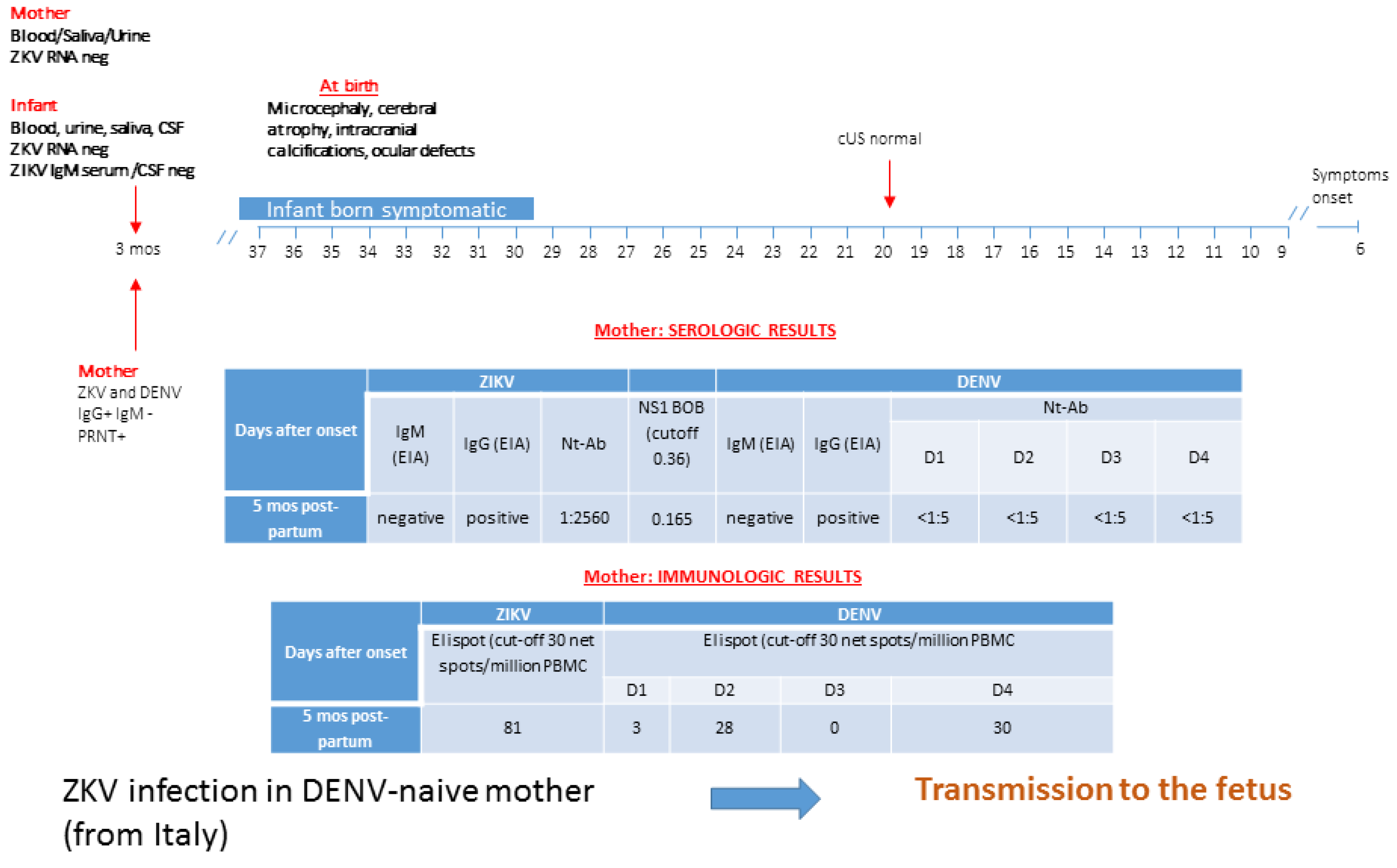
3.4. Patient 4: ZIKV Transmission in DENV-Naive Mother
3.5. Patient 5: ZIKV Transmission in DENV-Naive Mother
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ZIKV | Zika virus |
| DENV | Dengue virus |
| NS1 BOB | blockade-of-binding assay |
| YF | yellow fever |
| cMRI/cUS | cerebral magnetic reasonance imaging/cerebral ultrasonography |
| PRNT | plaque reduction neutralization test |
| Nt-Abs | neutralizing antibodies |
| EDE | envelope dimer epitope |
| ADE | antibody-dependent enhancement |
References
- Hazin, A.N.; Poretti, A.; Di Cavalcanti Souza Cruz, D.; Tenorio, M.; van der Linden, A.; Pena, L.J.; Brito, C.; Gil, L.H.; de Barros Miranda-Filho, D.; Marques, E.T.; et al. Microcephaly Epidemic Research Group. N. Engl. J. Med. 2016, 374, 2193–2195. [Google Scholar] [CrossRef] [PubMed]
- Honein, M.A.; Dawson, A.L.; Petersen, E.E.; Jones, A.M.; Lee, E.H.; Yazdy, M.M.; Ahmad, N.; Macdonald, J.; Evert, N.; Bingham, A.; et al. US Zika Pregnancy Registry Collaboration. JAMA 2017, 317, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Grubaugh, N.D.; Ishtiaq, F.; Setoh, Y.X.; Ko, A.I. Misperceived Risks of Zika-related Microcephaly in India. Trends Microbiol. 2019, 27, 381–383. [Google Scholar] [CrossRef] [PubMed]
- Cauchemez, S.; Besnard, M.; Garel, C.; Fontanet, A.; Mallet, H.P. Could clinical symptoms be a predictor of complications in Zika virus infection?—Authors’ reply. Lancet 2016, 388, 338–339. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.A.; Mier-y-Teran-Romero, L.; Reefhuis, J.; Gilboa, S.M.; Hills, S.L. Zika and the risk of microcephaly. N. Engl. J. Med. 2016, 375, 1–4. [Google Scholar] [CrossRef]
- Marinho, F.; Araújo, V.E.; Porto, D.L.; Ferreira, H.L.; Coelho, M.R.; Lecca, R.C.; Oliveira, H.; Poncioni, I.P.; Maranhão, M.H.; Mendes, Y.M.; et al. Microcephaly in Brazil: Prevalence and characterization of cases from the Information System on Live Births (Sinasc), 2000–2015. Epidemiol. Serv. Saude 2016, 25, 701–712. [Google Scholar] [CrossRef]
- Kleber de Oliveira, W.; Cortez-Escalante, J.; De Oliveira, W.T.; do Carmo, G.M.; Henriques, C.M.; Coelho, G.E.; Araújo de França, G.V. Increase in Reported Prevalence of Microcephaly in Infants Born to Women Living in Areas with Confirmed Zika Virus Transmission During the First Trimester of Pregnancy—Brazil, 2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 242–247. [Google Scholar] [CrossRef]
- Cuevas, E.L.; Tong, V.T.; Rozo, N.; Valencia, D.; Pacheco, O.; Gilboa, S.M.; Mercado, M.; Renquist, C.M.; González, M.; Ailes, E.C.; et al. Preliminary Report of Microcephaly Potentially Associated with Zika Virus Infection During Pregnancy—Colombia, January-November 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 1409–1413. [Google Scholar] [CrossRef] [Green Version]
- Meaney-Delman, D.; Hills, S.L.; Williams, C.; Galang, R.R.; Iyengar, P.; Hennenfent, A.K.; Rabe, I.B.; Panella, A.; Oduyebo, T.; Honein, M.A.; et al. Zika Virus Infection Among U.S. Pregnant Travelers—August 2015-February 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 211–214. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, W.K.; de França, G.V.A.; Carmo, E.H.; Duncan, B.B.; de Souza Kuchenbecker, R.; Schmidt, M.I. Infection-related microcephaly after the 2015 and 2016 Zika virus outbreaks in Brazil: A surveillance-based analysis. Lancet 2017, 390, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Hay, J.A.; Nouvellet, P.; Donnelly, C.A.; Riley, S. Potential inconsistencies in Zika surveillance data and our understanding of risk during pregnancy. PLoS Negl. Trop. Dis. 2018, 12, 6991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanstrom, J.A.; Plante, J.A.; Plante, K.S.; Young, E.F.; McGowan, E.; Gallichotte, E.N.; Widman, D.G.; Heise, M.T.; de Silva, A.M.; Baric, R.S. Dengue Virus Envelope Dimer Epitope Monoclonal Antibodies Isolated from Dengue Patients Are Protective against Zika Virus. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardina, S.V.; Bunduc, P.; Tripathi, S.; Duehr, J.; Frere, J.J.; Brown, J.A.; Nachbagauer, R.; Foster, G.A.; Krysztof, D.; Tortorella, D.; et al. Enhancement of Zika virus pathogenesis by preexisting antiflavivirus immunity. Science 2017, 356, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Barraquer, I.; Costa, F.; Nascimento, E.J.M.; Nery, N., Jr.; Castanha, P.M.S.; Sacramento, G.A.; Cruz, J.; Carvalho, M.; De Olivera, D.; Hagan, J.E.; et al. Impact of preexisting dengue immunity on Zika virus emergence in a dengue endemic region. Science 2019, 363, 607–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, A.; Gresh, L.; Ojeda, S.; Katzelnick, L.C.; Sanchez, N.; Mercado, J.C.; Chowell, G.; Lopez, B.; Elizondo, D.; Coloma, J.; et al. Prior dengue virus infection and risk of Zika: A pediatric cohort in Nicaragua. PLoS Med. 2019, 16, 1002726. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, H.S.; Calisher, C.H.; Mathews, J.H. Serum dilution neutralization test for California group virus identification and serology. J. Clin. Microbiol. 1976, 4, 503–510. [Google Scholar] [PubMed]
- Timiryasova, T.M.; Bonaparte, M.I.; Luo, P.; Zedar, R.; Hu, B.T.; Hildreth, S.W. Optimization and validation of a plaque reduction neutralization test for the detection of neutralizing antibodies to four serotypes of dengue virus used in support of dengue vaccine development. Am. J. Trop. Med. Hyg. 2013, 88, 962–970. [Google Scholar] [CrossRef]
- Scaramozzino, N.; Crance, J.M.; Jouan, A.; DeBriel, D.A.; Stoll, F.; Garin, D. Comparison of flavivirus universal primer pairs and development of a rapid, highly sensitive heminested reverse transcription-PCR assay for detection of flaviviruses targeted to a conserved region of the NS5 gene sequences. J. Clin. Microbiol. 2001, 39, 1922–1927. [Google Scholar] [CrossRef] [Green Version]
- Huhtamo, E.; Hasu, E.; Uzcátegui, N.Y.; Erra, E.; Nikkari, S.; Kantele, A.; Vapalahti, O.; Piiparinen, H. Early diagnosis of dengue in travelers: Comparison of a novel real-time RT-PCR, NS1 antigen detection and serology. J. Clin. Virol. 2010, 47, 49–53. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Balmaseda, A.; Stettler, K.; Medialdea-Carrera, R.; Collado, D.; Jin, X.; Zambrana, J.V.; Jaconi, S.; Cameroni, E.; Saborio, S.; Rovida, F.; et al. Antibody-based assay discriminates Zika virus infection from other flaviviruses. Proc. Natl. Acad. Sci. USA 2017, 114, 8384–8389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calarota, S.A.; Chiesa, A.; Zelini, P.; Comolli, G.; Minoli, L.; Baldanti, F. Detection of Epstein-Barr virus-specific memory CD4+ T cells using a peptide-based cultured enzyme-linked immunospot assay. Immunology 2013, 139, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Calarota, S.A.; Chiesa, A.; Scaramuzzi, L.; Adzasehoun, K.M.; Comolli, G.; Mangione, F.; Esposito, P.; Baldanti, F. Normalizing ELISPOT responses to T-cell counts: A novel approach for quantification of HCMV-specific CD4(+) and CD8(+) T-cell responses in kidney transplant recipients. J. Clin. Virol. 2014, 61, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central memory and effector memory T cell subsets: Function, generation, and maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef] [PubMed]
- Papageorghiou, A.T.; Thilaganathan, B.; Bilardo, C.M.; Ngu, A.; Malinger, G.; Herrera, M.; Salomon, L.J.; Riley, L.E.; Copel, J.A. ISUOG Interim Guidance on ultrasound for Zika virus infection in pregnancy: Information for healthcare professionals. Ultrasound Obstet. Gynecol. 2016, 47, 530–532. [Google Scholar] [CrossRef] [Green Version]
- International Society of Ultrasound in Obstetrics & Gynecology Education Committee. Sonographic examination of the fetal central nervous system: Guidelines for performing the ‘basic examination’ and the ‘fetal neurosonogram’. Ultrasound Obstet. Gynecol. 2007, 29, 109–116. [Google Scholar] [CrossRef]
- Schaub, B.; Vouga, M.; Najioullah, F.; Gueneret, M.; Monthieux, A.; Harte, C.; Muller, F.; Jolivet, E.; Adenet, C.; Dreux, S.; et al. Analysis of blood from Zika virus-infected fetuses: A prospective case series. Lancet Infect. Dis. 2017, 17, 520–527. [Google Scholar] [CrossRef] [Green Version]
- Pereira, J.P., Jr.; Maykin, M.M.; Vasconcelos, Z.; Avvad-Portari, E.; Zin, A.A.; Tsui, I.; Brasil, P.; Nielsen-Saines, K.; Moreira, M.E.; Gaw, S.L. The Role of Amniocentesis in the Diagnosis of Congenital Zika Syndrome. Clin. Infect. Dis 2019. [Google Scholar] [CrossRef]
- Oduyebo, T.; Igbinosa, I.; Petersen, E.E.; Polen, K.N.; Pillai, S.K.; Ailes, E.C.; Villanueva, J.M.; Newsome, K.; Fischer, M.; Gupta, P.M.; et al. Update: Interim Guidance for Health Care Providers Caring for Pregnant Women with Possible Zika Virus Exposure—United States, July 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Overview of Zika Virus Testing: Guidance for U.S. Laboratories Testing for Zika Virus Infection. Available online: https://www.cdc.gov/zika/laboratories/lab-guidance.html (accessed on 24th July 2017).
- Paz-Bailey, G.; Rosenberg, E.S.; Doyle, K.; Munoz-Jordan, J.; Santiago, G.A.; Klein, L.; Perez-Padilla, J.; Medina, F.A.; Waterman, S.H.; Gubern, C.G.; et al. Persistence of Zika Virus in Body Fluids—Final Report. N. Engl. J. Med. 2017, 379, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- De Góes Cavalcanti, L.P.; Tauil, P.L.; Alencar, C.H.; Oliveira, W.; Teixeira, M.M.; Heukelbach, J. Zika virus infection, associated microcephaly, and low yellow fever vaccination coverage in Brazil: Is there any causal link? J. Infect. Dev. Ctries. 2016, 10, 563–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salje, H.; Cummings, D.A.T.; Rodriguez-Barraquer, I.; Katzelnick, L.C.; Lessler, J.; Klungthong, C.; Thaisomboonsuk, B.; Nisalak, A.; Weg, A.; Ellison, D.; et al. Reconstruction of antibody dynamics and infection histories to evaluate dengue risk. Nature 2018, 557, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Percivalle, E.; Pacenti, M.; Rovida, F.; Zavattoni, M.; Del Bravo, P.; Cattelan, A.M.; Palù, G.; Baldanti, F. Virus and Antibody Dynamics in Travelers with Acute Zika Virus Infection. Clin. Infect. Dis. 2018, 66, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Chanama, S.; Anantapreecha, S.; A-nuegoonpipat, A.; Sa-gnasang, A.; Kurane, I.; Sawanpanyalert, P. Analysis of specific IgM responses in secondary dengue virus infections: Levels and positive rates in comparison with primary infections. J. Clin. Virol. 2004, 31, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Stettler, K.; Beltramello, M.; Espinosa, D.A.; Graham, V.; Cassotta, A.; Bianchi, S.; Vanzetta, F.; Minola, A.; Jaconi, S.; Mele, F.; et al. Specificity, cross-reactivity, and function of antibodies elicited by Zika virus infection. Science 2016, 353, 823–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassaniti, I.; Percivalle, E.; Adzasehoun, K.M.G.; Comolli, G.; Baldanti, F. Dengue Virus-Specific Humoral and T Cellular Immune Response in Italian Residents and Travelers Returning from Endemic Areas. Vector Borne Zoonotic Dis. 2019. [Google Scholar] [CrossRef]
- Grifoni, A.; Pham, J.; Sidney, J.; O’Rourke, P.H.; Paul, S.; Peters, B.; Martini, S.R.; de Silva, A.D.; Ricciardi, M.J.; Magnani, D.M.; et al. Prior Dengue Virus Exposure Shapes T Cell Immunity to Zika Virus in Humans. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Suy, A.; Sulleiro, E.; Rodó, C.; Vázquez, É.; Bocanegra, C.; Molina, I.; Esperalba, J.; Sánchez-Seco, M.P.; Boix, H.; Pumarola, T.; et al. Prolonged Zika Virus Viremia during Pregnancy. N. Engl. J. Med. 2016, 375, 2611–2613. [Google Scholar] [CrossRef]
- Tabata, T.; Petitt, M.; Puerta-Guardo, H.; Michlmayr, D.; Harris, E.; Pereira, L. Zika Virus Replicates in Proliferating Cells in Explants from First-Trimester Human Placentas, Potential Sites for Dissemination of Infection. J. Infect. Dis. 2018, 217, 1202–1213. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.J.; Frosch, M.P. Zika and histopathology in first trimester infections. Lancet 2016, 388, 847–849. [Google Scholar] [CrossRef] [Green Version]
- Sabin, A.B. Research on dengue during World War II. Am. J. Trop. Med. Hyg. 1952, 1, 30–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyamvada, L.; Quicke, K.M.; Hudson, W.H.; Onlamoon, N.; Sewatanon, J.; Edupuganti, S.; Pattanapanyasat, K.; Chokephaibulkit, K.; Mulligan, M.J.; Wilson, P.C.; et al. Human antibody responses after dengue virus infection are highly cross-reactive to Zika virus. Proc. Natl. Acad. Sci. USA 2106, 113, 7852–7857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat. Immunol. 2016, 9, 1102–1108. [Google Scholar] [CrossRef]
- Halstead, S.B. Neutralization and antibody-dependent enhancement of dengue viruses. Adv. Virus Res. 2003, 60, 421–467. [Google Scholar]
- Gibbons, R.V.; Kalanarooj, S.; Jarman, R.G.; Nisalak, A.; Vaughn, D.W.; Endy, T.P.; Mammen, M.P., Jr.; Srikiatkhachorn, A. Analysis of repeat hospital admissions for dengue to estimate the frequency of third or fourth dengue infections resulting in admissions and dengue hemorrhagic fever, and serotype sequences. Am. J. Trop. Med. Hyg. 2007, 77, 910–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olkowski, S.; Forshey, B.M.; Morrison, A.C.; Rocha, C.; Vilcarromero, S.; Halsey, E.S.; Kochel, T.J.; Scott, T.W.; Stoddard, S.T. Reduced risk of disease during postsecondary dengue virus infections. J. Infect. Dis. 2013, 208, 1026–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauchemez, S.; Besnard, M.; Bompard, P.; Dub, T.; Guillemette-Artur, P.; Eyrolle-Guignot, D.; Salje, H.; Van Kerkhove, M.D.; Abadie, V.; et al. Association between Zika virus and microcephaly in French Polynesia, 2013–2015: A retrospective study. Lancet 2016, 387, 2125–2132. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Patient #1 | Patient #2 | Patient #3 | Patient #4 | Patient #5 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| days after symptoms onset | 5 | 16 | 31 | 59 | 74 | 165 | 198 | 107 | 154 | 34 | 95 | 11 | 56 | 60 | 3 months post-partum |
| ZIKV RNA in blood | Pos | Pos | Pos | Pos | Pos | Pos | Neg | Pos | Pos | Pos | Pos | Neg | |||
| ZIKV RNA in urine | Pos | Pos | Neg | Pos | Neg | Neg | Pos | Neg | |||||||
| ZIKV RNA in saliva | Pos | Neg | Neg | Neg | Pos | Neg | Neg | ||||||||
| ZIKV RNA in AF | Neg | Neg | Pos | ||||||||||||
| ZIKV IgM EIA | Neg | Neg | Neg | Neg | Neg | Neg | Pos | Neg | Neg | ||||||
| ZIKV IgG EIA | Pos | Pos | Pos | Pos | Pos | Pos | Neg | Pos | Pos | ||||||
| ZIKV NT Abs | 1:640 | 1:1280 | 1:2560 | 1:160 | 1:160 | 1:2560 | 1:2560 | ||||||||
| DENV IgM EIA | Neg | Neg | Neg | Neg | Neg | Neg | Neg | ||||||||
| DENV IgG EIA | Pos | Pos | Pos | Pos | Pos | Pos | Pos | ||||||||
| DENV NT Abs | |||||||||||||||
| DENV1 | 1:2560 | 1:40,000 | 1:640,000 | 1:5000 | 1:320 | <5 | <5 | ||||||||
| DENV2 | 1:160,000 | 1:640,000 | 1:640,000 | 1:320 | 1:320 | <5 | <5 | ||||||||
| DENV 3 | 1:640,000 | 1:640,000 | 1:640,000 | 1:640 | 1:320 | <5 | <5 | ||||||||
| DENV 4 | 1:640 | 1:2560 | 1:640,000 | 1:80 | 1:80 | <5 | <5 | ||||||||
| BOB assay * | 0.179 | 0.197 | 0.156 | 0.151 | 0.165 | ||||||||||
| ZIKV ELISPOT ** | 61 | 81 | |||||||||||||
| DENV ELISPOT ** | |||||||||||||||
| DENV1 | 153 | 5 | 3 | ||||||||||||
| DENV2 | 163 | 5 | 28 | ||||||||||||
| DENV 3 | 198 | 3 | 0 | ||||||||||||
| DENV 4 | 118 | 10 | 30 | ||||||||||||
| outcome | no transmission | no transmission | no transmission | transmission | transmission | ||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zavattoni, M.; Rovida, F.; Percivalle, E.; Cassaniti, I.; Sarasini, A.; Arossa, A.; Tassis, B.; Bollani, L.; Lombardi, G.; Orcesi, S.; et al. Zika Virus Infection in Pregnancy: Advanced Diagnostic Approaches in Dengue-Naive and Dengue-Experienced Pregnant Women and Possible Implication for Cross-Reactivity and Cross-Protection. Microorganisms 2020, 8, 56. https://doi.org/10.3390/microorganisms8010056
Zavattoni M, Rovida F, Percivalle E, Cassaniti I, Sarasini A, Arossa A, Tassis B, Bollani L, Lombardi G, Orcesi S, et al. Zika Virus Infection in Pregnancy: Advanced Diagnostic Approaches in Dengue-Naive and Dengue-Experienced Pregnant Women and Possible Implication for Cross-Reactivity and Cross-Protection. Microorganisms. 2020; 8(1):56. https://doi.org/10.3390/microorganisms8010056
Chicago/Turabian StyleZavattoni, Maurizio, Francesca Rovida, Elena Percivalle, Irene Cassaniti, Antonella Sarasini, Alessia Arossa, Beatrice Tassis, Lina Bollani, Giuseppina Lombardi, Simona Orcesi, and et al. 2020. "Zika Virus Infection in Pregnancy: Advanced Diagnostic Approaches in Dengue-Naive and Dengue-Experienced Pregnant Women and Possible Implication for Cross-Reactivity and Cross-Protection" Microorganisms 8, no. 1: 56. https://doi.org/10.3390/microorganisms8010056
APA StyleZavattoni, M., Rovida, F., Percivalle, E., Cassaniti, I., Sarasini, A., Arossa, A., Tassis, B., Bollani, L., Lombardi, G., Orcesi, S., & Baldanti, F. (2020). Zika Virus Infection in Pregnancy: Advanced Diagnostic Approaches in Dengue-Naive and Dengue-Experienced Pregnant Women and Possible Implication for Cross-Reactivity and Cross-Protection. Microorganisms, 8(1), 56. https://doi.org/10.3390/microorganisms8010056