Pseudomonas aeruginosa Toxin ExoU as a Therapeutic Target in the Treatment of Bacterial Infections

 , and
, and
Abstract
1. Introduction
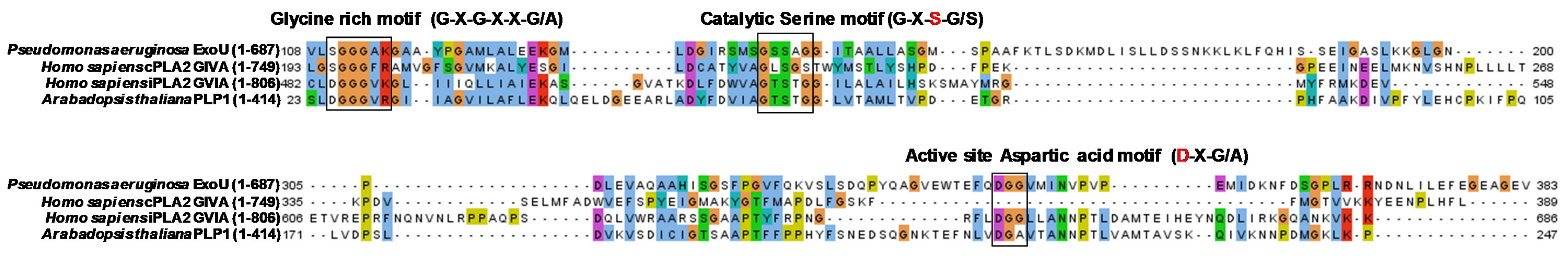
2. Exotoxin U
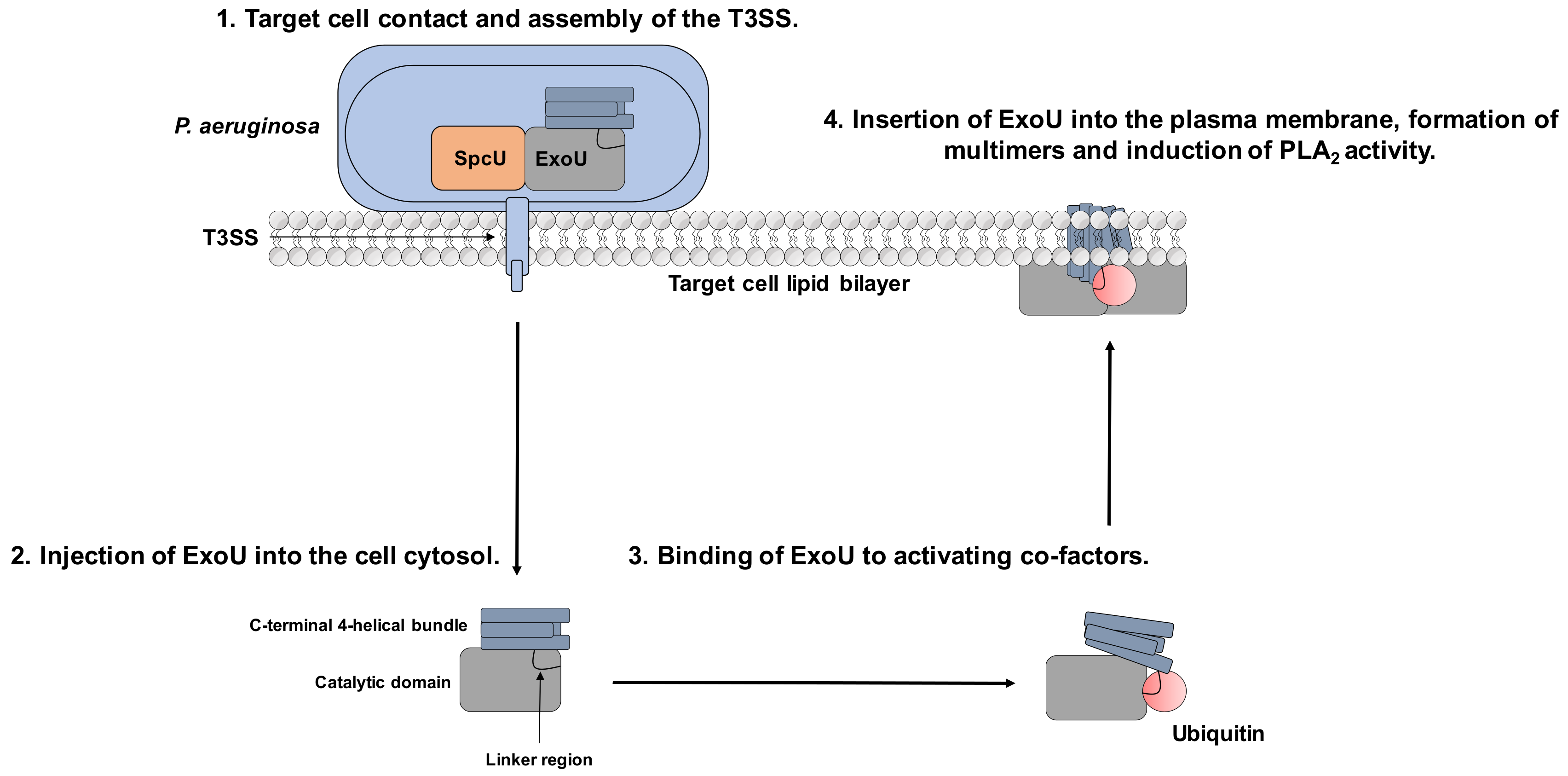
2.1. Secretion of ExoU via the T3SS
2.2. Control of T3SS Gene Expression
2.3. Oligomerisation and Localisation to the Host Cell Wall in the Presence of Phosphatidylinositol 4,5-Bisphosphate (PIP2)
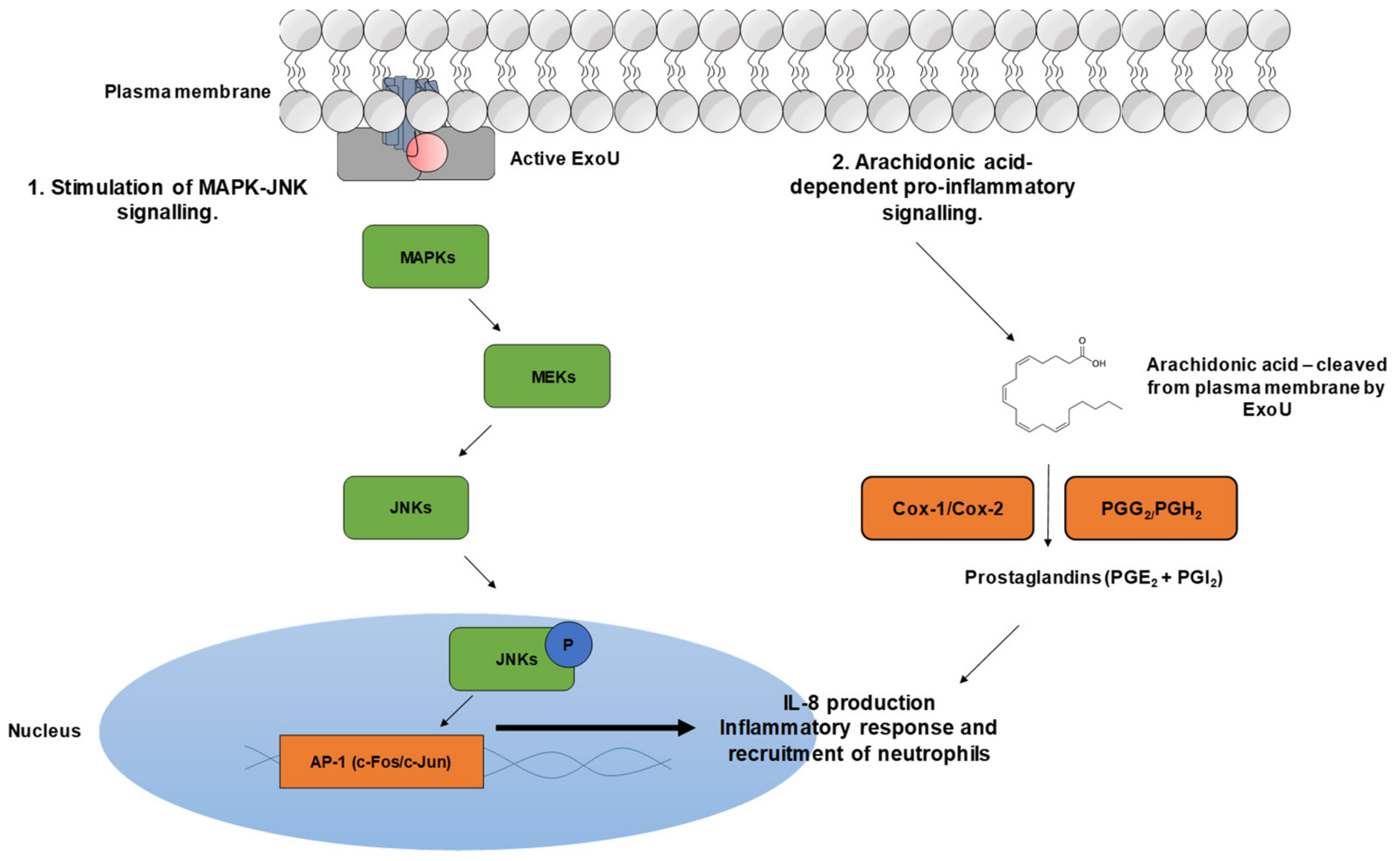
2.4. Interactions with Host Cell Signalling Pathways
3. Pharmacological Targeting of the Bacterial Phospholipase ExoU
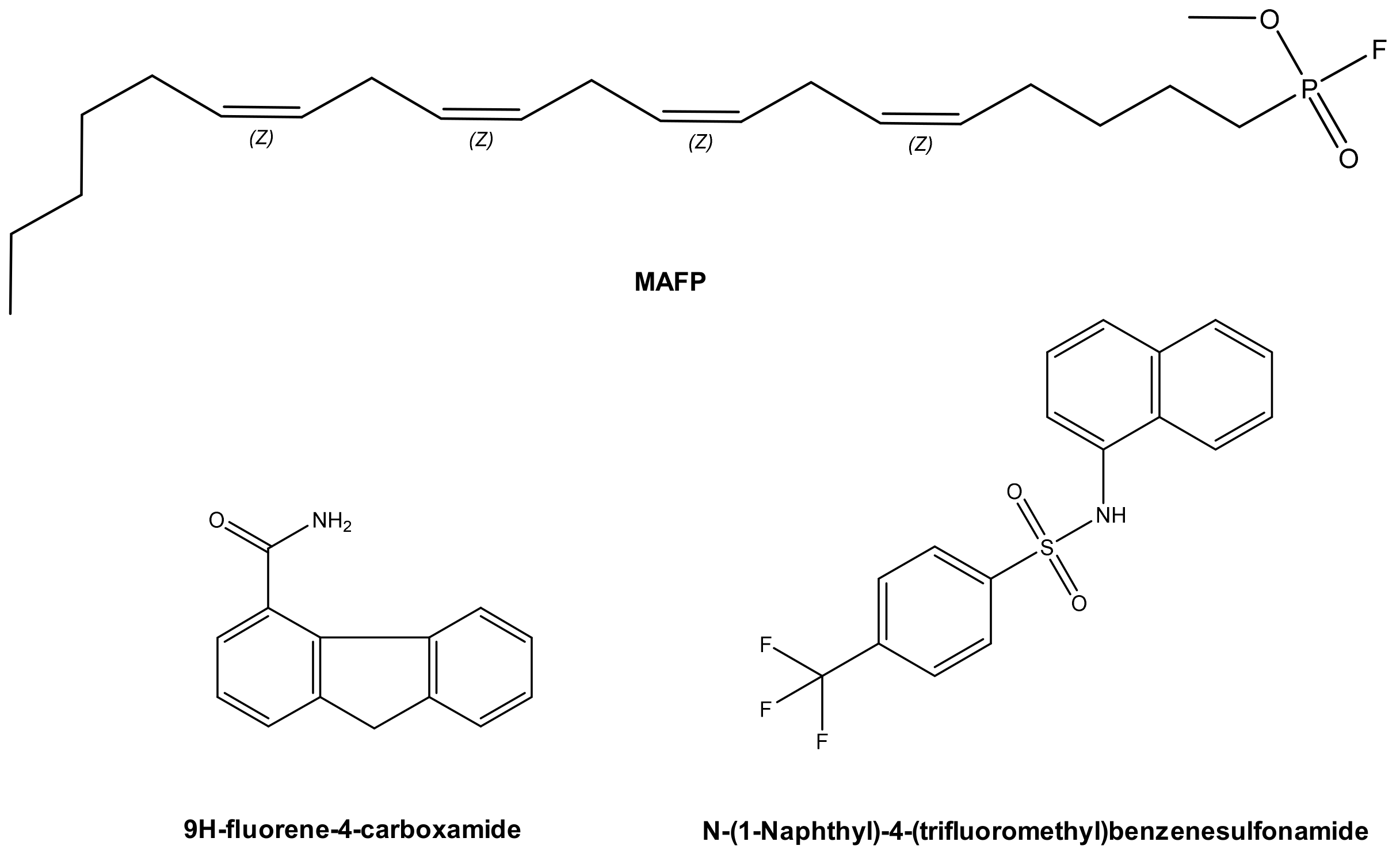
3.1. Allosteric Inhibitors
3.2. Ligand Repurposing Approaches
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Newman, J.W.; Floyd, R.V.; Fothergill, J.L. The contribution of Pseudomonas aeruginosa virulence factors and host factors in the establishment of urinary tract infections. FEMS Microbiol. Lett. 2017, 364, 1–12. [Google Scholar] [CrossRef]
- Stewart, R.M.K.; Wiehlmann, L.; Ashelford, K.E.; Preston, S.J.; Frimmersdorf, E.; Campbell, B.J.; Neal, T.J.; Hall, N.; Tuft, S.; Kaye, S.B.; et al. Genetic characterization indicates that a specific subpopulation of Pseudomonas aeruginosa is associated with keratitis infections. J. Clin. Microbiol. 2011, 49, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Schulert, G.S.; Feltman, H.; Rabin, S.D.; Martin, C.G.; Battle, S.E.; Rello, J.; Hauser, A.R. Secretion of the toxin ExoU is a marker for highly virulent Pseudomonas aeruginosa isolates obtained from patients with hospital-acquired pneumonia. J. Infect. Dis. 2003, 188, 1695–1706. [Google Scholar] [CrossRef] [PubMed]
- Roy-Burman, A.; Savel, R.H.H.; Racine, S.; Swanson, B.L.L.; Revadigar, N.S.S.; Fujimoto, J.; Sawa, T.; Frank, D.W.W.; Wiener-Kronish, J.P.; Roy-Burman, A.; et al. Type III Protein Secretion Is Associated with Death in Lower Respiratory and Systemic Pseudomonas aeruginosa Infections. J. Infect. Dis. 2002, 183, 1767–1774. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.R.; Cobb, E.; Bodi, M.; Mariscal, D.; Valles, J.; Engel, J.N.; Rello, J.; Bodí, M.; Mariscal, D.; Vallés, J.; et al. Type III protein secretion is associated with poor clinical outcomes in patients with ventilator-associated pneumonia caused by Pseudomonas aeruginosa. Crit. Care Med. 2002, 30, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Migiyama, Y.; Yanagihara, K.; Kaku, N.; Harada, Y.; Yamada, K.; Nagaoka, K.; Morinaga, Y.; Akamatsu, N.; Matsuda, J.; Izumikawa, K.; et al. Pseudomonas aeruginosa bacteremia among immunocompetent and immunocompromised patients: Relation to initial antibiotic therapy and survival. Jpn. J. Infect. Dis. 2016, 69, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Bhagirath, A.Y.; Li, Y.; Somayajula, D.; Dadashi, M.; Badr, S.; Duan, K. Cystic fibrosis lung environment and Pseudomonas aeruginosa infection. BMC Pulm. Med. 2016, 16, 1–23. [Google Scholar] [CrossRef]
- Fernández-Barat, L.; Ferrer, M.; De Rosa, F.; Gabarrús, A.; Esperatti, M.; Terraneo, S.; Rinaudo, M.; Li Bassi, G.; Torres, A. Intensive care unit-acquired pneumonia due to Pseudomonas aeruginosa with and without multidrug resistance. J. Infect. 2017, 74, 142–152. [Google Scholar] [CrossRef]
- World Health Organisation. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibitoics. Available online: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf?ua=1 (accessed on 25 February 2017).
- Klockgether, J.; Tümmler, B. Recent advances in understanding Pseudomonas aeruginosa as a pathogen. F1000Research 2017, 6, 1261. [Google Scholar] [CrossRef]
- Finck-Barbançon, V.; Goranson, J.; Zhu, L.; Sawa, T.; Wiener-Kronish, J.P.; Fleiszig, S.M.J.; Wu, C.; Mende-Mueller, L.; Frank, D.W.; Finck-Barbançon, V.; et al. ExoU expression by Pseudomonas aeruginosa correlates with acute cytotoxicity and epithelial injury. Mol. Microbiol. 1997, 25, 547–557. [Google Scholar] [CrossRef]
- Tam, C.; Lewis, S.E.; Li, W.Y.; Lee, E.; Evans, D.J.; Fleiszig, S.M.J. Mutation of the phospholipase catalytic domain of the Pseudomonas aeruginosa cytotoxin ExoU abolishes colonization promoting activity and reduces corneal disease severity. Exp. Eye Res. 2007, 85, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-H.; Kim, S.Y.; Roh, E.Y.; Lee, H.S. Difference of Type 3 secretion system (T3SS) effector gene genotypes (exoU and exoS) and its implication to antibiotics resistances in isolates of Pseudomonas aeruginosa from chronic otitis media. Auris. Nasus. Larynx 2017, 44, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Galle, M.; Carpentier, I.; Beyaert, R. Structure and function of the Type III secretion system of Pseudomonas aeruginosa. Curr. Protein Pept. Sci. 2012, 13, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Monday, S.R.; Schiller, N.L. Alginate synthesis in Pseudomonas aeruginosa: The role of AlgL (alginate lyase) and AlgX. J. Bacteriol. 1996, 178, 625–632. [Google Scholar] [CrossRef]
- Poole, K. Efflux-mediated antimicrobial resistance. Antibiot. Discov. Dev. 2005, 56, 20–51. [Google Scholar] [CrossRef]
- Thibodeaux, B.A.; Caballero, A.R.; Marquart, M.E.; Tommassen, J.; O’Callaghan, R.J. Corneal Virulence of Pseudomonas aeruginosa Elastase B and Alkaline Protease Produced by Pseudomonas putida. Curr. Eye Res. 2007, 32, 373–386. [Google Scholar] [CrossRef]
- Hong, Y.; Ghebrehiwet, B. Effect of Pseudomonas aeruginosa elastase and alkaline protease on serum complement and isolated components C1q and C3. Clin. Immunol. Immunopathol. 1992, 62, 133–138. [Google Scholar] [CrossRef]
- Parmely, M.; Gale, A.; Clabaugh, M.; Horvat, R.; Zhou, W.W. Proteolytic inactivation of cytokines by Pseudomonas aeruginosa. Infect. Immun. 1990, 58, 3009–3014. [Google Scholar]
- Bahl, C.D.; Morisseau, C.; Bomberger, J.M.; Stanton, B.A.; Hammock, B.D.; O’Toole, G.A.; Madden, D.R. Crystal structure of the cystic fibrosis transmembrane conductance regulator inhibitory factor cif reveals novel active-site features of an epoxide hydrolase virulence factor. J. Bacteriol. 2010, 192, 1785–1795. [Google Scholar] [CrossRef]
- Ballok, A.E.; O’Toole, G.A. Pouring salt on a wound: Pseudomonas aeruginosa virulence factors alter Na+ and Cl- flux in the lung. J. Bacteriol. 2013, 195, 4013–4019. [Google Scholar] [CrossRef]
- Flitter, B.A.; Hvorecny, K.L.; Ono, E.; Eddens, T.; Yang, J.; Kwak, D.H.; Bahl, C.D.; Hampton, T.H.; Morisseau, C.; Hammock, B.D.; et al. Pseudomonas aeruginosa sabotages the generation of host proresolving lipid mediators. Proc. Natl. Acad. Sci. USA 2017, 114, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Bomberger, J.M.; Ely, K.H.; Bangia, N.; Ye, S.; Green, K.A.; Green, W.R.; Enelow, R.I.; Stanton, B.A. Pseudomonas aeruginosa Cif protein enhances the ubiquitination and proteasomal degradation of the transporter associated with antigen processing (TAP) and reduces major histocompatibility complex (MHC) class I antigen presentation. J. Biol. Chem. 2014, 289, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Michalska, M.; Wolf, P. Pseudomonas Exotoxin A: Optimized by evolution for effective killing. Front. Microbiol. 2015, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.L.; Coburn, J.; Rucks, E.A.; Olson, J.C. Characterization of Pseudomonas aeruginosa exoenzyme S as a bifunctional enzyme in J774a.1 macrophages. Infect. Immun. 2003, 71, 5296–5305. [Google Scholar] [CrossRef]
- Sun, J.; Barbieri, J.T. Pseudomonas aeruginosa ExoT ADP-ribosylates CT10 regulator of kinase (Crk) proteins. J. Biol. Chem. 2003, 278, 32794–32800. [Google Scholar] [CrossRef]
- Barbieri, J.T.; Sun, J. Pseudomonas aeruginosa ExoS and ExoT. In Reviews of Physiology, Biochemistry and Pharmacology; Springer: Berlin/Heidelberg, Germany, 2004; pp. 79–92. [Google Scholar]
- Galle, M.; Jin, S.; Bogaert, P.; Haegman, M.; Vandenabeele, P.; Beyaert, R. The pseudomonas aeruginosa type III secretion system has an exotoxin S/T/Y independent pathogenic role during acute lung infection. PLoS ONE 2012, 7, e41547. [Google Scholar] [CrossRef]
- Sato, H.; Frank, D.W.; Hillard, C.J.; Feix, J.B.; Pankhaniya, R.R.; Moriyama, K.; Finck-Barbancon, V.; Buchaklian, A.; Lei, M.; Long, R.M.; et al. The mechanism of action of the Pseudomonas aeruginosa-encoded type III cytotoxin, ExoU. EMBO J. 2003, 22, 2959–2969. [Google Scholar] [CrossRef]
- Anderson, D.M.; Schmalzer, K.M.; Sato, H.; Casey, M.; Terhune, S.S.; Haas, A.L.; Feix, J.B.; Frank, D.W. Ubiquitin and ubiquitin-modified proteins activate the Pseudomonas aeruginosa T3SS cytotoxin, ExoU. Mol. Microbiol. 2011, 82, 1454–1467. [Google Scholar] [CrossRef]
- Miao, E.A.; Andersen-Nissen, E.; Warren, S.E.; Aderem, A. TLR5 and Ipaf: Dual sensors of bacterial flagellin in the innate immune system. Semin. Immunopathol. 2007, 29, 275–288. [Google Scholar] [CrossRef]
- Spencer, J.; Murphy, L.M.; Conners, R.; Sessions, R.B.; Gamblin, S.J. Crystal structure of the lasA virulence factor from Pseudomonas aeruginosa: Substrate specificity and mechanism of M23 metallopeptidases. J. Mol. Biol. 2010, 396, 908–923. [Google Scholar] [CrossRef]
- Kessler, E.; Safrin, M.; Gustin, J.K.; Ohman, D.E. Elastase and the LasA protease of Pseudomonas aeruginosa are secreted with their propeptides. J. Biol. Chem. 1998, 273, 30225–30231. [Google Scholar] [CrossRef] [PubMed]
- Wargo, M.J.; Ho, T.C.; Gross, M.J.; Whittaker, L.A.; Hogan, D.A. GbdR regulates Pseudomonas aeruginosa plcH and pchP transcription in response to choline catabolites. Infect. Immun. 2009, 77, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Ostroff, R.M.; Vasil, A.I.; Vasil, M.L. Molecular comparison of a nonhemolytic and a hemolytic phospholipase C from Pseudomonas aeruginosa. J. Bacteriol. 1990, 172, 5915–5923. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.; Brown, H.A. Biochemical characterization of a pseudomonas aeruginosa phospholipase d. Biochemistry 2015, 54, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Waterfield, N.R.; Yang, J.; Yang, G.; Jin, Q. A Pseudomonas aeruginosa type VI secretion phospholipase D effector targets both prokaryotic and eukaryotic cells. Cell Host Microbe 2014, 15, 600–610. [Google Scholar] [CrossRef]
- Traidej, M.; Marquart, M.E.; Caballero, A.R.; Thibodeaux, B.A.; O’Callaghan, R.J. Identification of the active site residues of Pseudomonas aeruginosa protease IV: Importance of enzyme activity in autoprocessing and activation. J. Biol. Chem. 2003, 278, 2549–2553. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, S.K.; So, Y.I.; Park, H.Y.; Li, X.H. Protease IV, a quorum sensing-dependent protease of Pseudomonas aeruginosa modulates insect innate immunity. Mol. Microbiol. 2014, 94, 1298–1314. [Google Scholar] [CrossRef]
- Lau, G.W.; Ran, H.; Kong, F.; Hassett, D.J.; Mavrodi, D. Pseudomonas aeruginosa pyocyanin is critical for lung infection in mice. Infect. Immun. 2004, 72, 4275–4278. [Google Scholar] [CrossRef]
- Davey, M.E.; Caiazza, N.C.; O’Toole, G.A. Rhamnolipid surfactant production affects biofilm architecture in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2003, 185, 1027–1036. [Google Scholar] [CrossRef]
- Jiang, F.; Wang, X.; Wang, B.; Chen, L.; Zhao, Z.; Waterfield, N.R.; Yang, G.; Jin, Q. The Pseudomonas aeruginosa Type VI Secretion PGAP1-like Effector Induces Host Autophagy by Activating Endoplasmic Reticulum Stress. Cell Rep. 2016, 16, 1502–1509. [Google Scholar] [CrossRef]
- Borkar, D.S.; Fleiszig, S.M.J.; Leong, C.; Lalitha, P.; Srinivasan, M.; Ghanekar, A.A.; Tam, C.; Li, W.Y.; Zegans, M.E.; McLeod, S.D.; et al. Association between cytotoxic and invasive pseudomonas aeruginosa and clinical outcomes in bacterial keratitis. JAMA Ophthalmol. 2013, 131, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Goehring, U.M.; Schmidt, G.; Pederson, K.J.; Aktories, K.; Barbieri, J.T. The N-terminal domain of Pseudomonas aeruginosa exoenzyme S is a GTPase- activating protein for Rho GTPases. J. Biol. Chem. 1999, 274, 36369–36372. [Google Scholar] [CrossRef]
- Vareechon, C.; Zmina, S.E.; Karmakar, M.; Pearlman, E.; Rietsch, A. Pseudomonas aeruginosa Effector ExoS Inhibits ROS Production in Human Neutrophils. Cell Host Microbe 2017, 21, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, M.R.; Jia, J.; Zeng, L.; Ha, U.; Chow, M.; Jin, S. Pseudomonas aeruginosa mediated apoptosis requires the ADP-ribosylating activity of ExoS. Microbiology 2000, 146, 2531–2541. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Frank, D.W. ExoU is a potent intracellular phospholipase. Mol. Microbiol. 2004, 53, 1279–1290. [Google Scholar] [CrossRef]
- Phillips, R.M.; Six, D.A.; Dennis, E.A.; Ghosh, P. In vivo phospholipase activity of the Pseudomonas aeruginosa cytotoxin ExoU and protection of mammalian cells with phospholipase A2 inhibitors. J. Biol. Chem. 2003, 278, 41326–41332. [Google Scholar] [CrossRef] [PubMed]
- Pankhaniya, R.R.; Tamura, M.; Allmond, L.R.; Moriyama, K.; Ajayi, T.; Wiener-Kronish, J.P.; Sawa, T. Pseudomonas aeruginosa causes acute lung injury via the catalytic activity of the patatin-like phospholipase domain of ExoU. Crit. Care Med. 2004, 32, 2293–2299. [Google Scholar] [CrossRef]
- El Solh, A.A.; Akinnusi, M.E.; Wiener-Kronish, J.P.; Lynch, S.V.; Pineda, L.A.; Szarpa, K. Persistent infection with Pseudomonas aeruginosa in ventilator-associated pneumonia. Am. J. Respir. Crit. Care Med. 2008, 178, 513–519. [Google Scholar] [CrossRef]
- Diaz, M.H.; Shaver, C.M.; King, J.D.; Musunuri, S.; Kazzaz, J.A.; Hauser, A.R. Pseudomonas aeruginosa induces localized immunosuppression during pneumonia. Infect. Immun. 2008, 76, 4414–4421. [Google Scholar] [CrossRef]
- Engel, J.; Balachandran, P. Role of Pseudomonas aeruginosa type III effectors in disease. Curr. Opin. Microbiol. 2009, 12, 61–66. [Google Scholar] [CrossRef]
- Rabin, S.D.P.P.; Hauser, A.R. Functional regions of the Pseudomonas aeruginosa cytotoxin ExoU. Infect. Immun. 2005, 73, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Apodaca, G.; Bomsel, M.; Lindstedt, R.; Engel, J.; Frank, D.; Mostov, K.E.; Wiener- Kronish, J. Characterization of Pseudomonas aeruginosa-induced MDCK cell injury: Glycosylation-defective host cells are resistant to bacterial killing. Infect. Immun. 1995, 63, 1541–1551. [Google Scholar] [PubMed]
- Wiener-Kronish, J.P.; Sakuma, T.; Kudoh, I.; Pittet, J.F.; Frank, D.; Dobbs, L.; Vasil, M.L.; Matthay, M.A. Alveolar epithelial injury and pleural empyema in acute P. aeruginosa pneumonia in anesthetized rabbits. J. Appl. Physiol. 1993, 74, 1661–1669. [Google Scholar] [CrossRef] [PubMed]
- Pittet, J.F.; Wiener-Kronish, J.P.; McElroy, M.C.; Folkesson, H.G.; Matthay, M.A. Stimulation of lung epithelial liquid clearance by endogenous release of catecholamines in septic shock in anesthetized rats. J. Clin. Invest. 1994, 94, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Jung Kang, P.; Hauser, A.R.; Apodaca, G.; Fleiszig, S.M.J.; Wiener-Kronish, J.; Mostov, K.; Engel, J.N. Identification of Pseudomonas aeruginosa genes required for epithelial cell injury. Mol. Microbiol. 1997, 24, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Feix, J.B.; Frank, D.W. Identification of superoxide dismutase as a cofactor for the pseudomonas type III toxin, ExoU. Biochemistry 2006, 45, 10368–10375. [Google Scholar] [CrossRef] [PubMed]
- Stirling, F.R.; Cuzick, A.; Kelly, S.M.; Oxley, D.; Evans, T.J. Eukaryotic localization, activation and ubiquitinylation of a bacterial type III secreted toxin. Cell. Microbiol. 2006, 8, 1294–1309. [Google Scholar] [CrossRef]
- Anderson, D.M.; Sato, H.; Dirck, A.T.; Feix, J.B.; Frank, D.W. Ubiquitin activates patatin-like phospholipases from multiple bacterial species. J. Bacteriol. 2015, 197, 529–541. [Google Scholar] [CrossRef]
- Finck-Barbancon, V.; Frank, D.W. Multiple domains are required for the toxic activity of Pseudomonas aeruginosa ExoU. J. Bacteriol. 2001, 183, 4330–4344. [Google Scholar] [CrossRef]
- Parsot, C.; Hamiaux, C.; Page, A.L. The various and varying roles of specific chaperones in type III secretion systems. Curr. Opin. Microbiol. 2003, 6, 7–14. [Google Scholar] [CrossRef]
- Akeda, Y.; Galán, J.E. Chaperone release and unfolding of substrates in type III secretion. Nature 2005, 437, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Halavaty, A.S.; Borek, D.; Tyson, G.H.; Veesenmeyer, J.L.; Shuvalova, L.; Minasov, G.; Otwinowski, Z.; Hauser, A.R.; Anderson, W.F. Structure of the Type III Secretion Effector Protein ExoU in Complex with Its Chaperone SpcU. PLoS ONE 2012, 7, e49388. [Google Scholar] [CrossRef] [PubMed]
- Rydel, T.J.; Williams, J.M.; Krieger, E.; Moshiri, F.; Stallings, W.C.; Brown, S.M.; Pershing, J.C.; Purcell, J.P.; Alibhai, M.F. The crystal structure, mutagenesis, and activity studies reveal that patatin is a lipid acyl hydrolase with a Ser-Asp catalytic dyad. Biochemistry 2003, 42, 6696–6708. [Google Scholar] [CrossRef] [PubMed]
- Hirschberg, H.J.H.B.; Simons, J.W.F.A.; Dekker, N.; Egmond, M.R. Cloning, expression, purification and characterization of patatin, a novel phospholipase A. Eur. J. Biochem. 2001, 268, 5037–5044. [Google Scholar] [CrossRef]
- Dessen, A.; Tang, J.; Schmidt, H.; Stahl, M.; Clark, J.D.; Seehra, J.; Somers, W.S. Crystal structure of human cytosolic phospholipase A2 reveals a novel topology and catalytic mechanism. Cell 1999, 97, 349–360. [Google Scholar] [CrossRef]
- Gendrin, C.; Contreras-Martel, C.; Bouillot, S.S.; Elsen, S.; Lemaire, D.; Skoufias, D.A.; Huber, P.; Attree, I.; Dessen, A.A. Structural basis of cytotoxicity mediated by the type III secretion toxin ExoU from Pseudomonas aeruginosa. PLoS Pathog. 2012, 8, e1002637. [Google Scholar] [CrossRef]
- Geissler, B.; Tungekar, R.; Satchell, K.J.F. Identification of a conserved membrane localization domain within numerous large bacterial protein toxins. Proc. Natl. Acad. Sci. USA 2010, 107, 5581–5586. [Google Scholar] [CrossRef]
- Geissler, B.; Ahrens, S.; Satchell, K.J.F. Plasma membrane association of three classes of bacterial toxins is mediated by a basic-hydrophobic motif. Cell. Microbiol. 2012, 14, 286–298. [Google Scholar] [CrossRef]
- Yahr, T.L.; Barbieri, J.T.; Frank, D.W. Genetic relationship between the 53- and 49-kilodalton forms of exoenzyme S from Pseudomonas aeruginosa. J. Bacteriol. 1996, 178, 1412–1419. [Google Scholar] [CrossRef]
- Pastor, A.; Chabert, J.; Louwagie, M.; Garin, J.; Attree, I. PscF is a major component of the Pseudomonas aeruginosa type III secretion needle. FEMS Microbiol. Lett. 2005, 253, 95–101. [Google Scholar] [CrossRef]
- Mueller, C.A.; Broz, P.; Cornelis, G.R. The type III secretion system tip complex and translocon. Mol. Microbiol. 2008, 68, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Sarker, M.R.; Neyt, C.; Stainier, I.; Cornelis, G.R. The Yersinia Yop virulon: Lcr V is required to extrusion of the translocators YopB and YopD. J. Bacteriol. 1998, 180, 1207–1214. [Google Scholar] [PubMed]
- Halder, P.K.; Roy, C.; Datta, S. Structural and functional characterization of type three secretion system ATPase PscN and its regulator PscL from Pseudomonas aeruginosa. Proteins Struct. Funct. Bioinform. 2019, 87, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Yahr, T.L.; Goranson, J.; Frank, D.W. Exoenzyme of S of Pseudomonas aeruginosa is secreted by a type III pathway. Mol. Microbiol. 1996, 22, 991–1003. [Google Scholar] [CrossRef]
- Galán, J.E.; Lara-Tejero, M.; Marlovits, T.C.; Wagner, S. Bacterial Type III Secretion Systems: Specialized Nanomachines for Protein Delivery into Target Cells. Annu. Rev. Microbiol. 2014, 68, 415–438. [Google Scholar] [CrossRef]
- Kim, J.; Ahn, K.; Min, S.; Jia, J.; Ha, U.; Wu, D.; Jin, S. Factors triggering type III secretion in Pseudomonas aeruginosa. Microbiology 2005, 151, 3575–3587. [Google Scholar] [CrossRef]
- Williams McMackin, E.A.; Djapgne, L.; Corley, J.M.; Yahr, T.L. Fitting Pieces into the Puzzle of Pseudomonas aeruginosa Type III Secretion System Gene Expression. J. Bacteriol. 2019, 201, e00209-19. [Google Scholar] [CrossRef]
- Zheng, Z.; Chen, G.; Joshi, S.; Brutinel, E.D.; Yahr, T.L.; Chen, L. Biochemical characterization of a regulatory cascade controlling transcription of the Pseudomonas aeruginosa type III secretion system. J. Biol. Chem. 2007, 62, 631–640. [Google Scholar]
- Thibault, J.; Faudry, E.; Ebel, C.; Attree, I.; Elsen, S. Anti-activator ExsD forms a 1:1 complex with ExsA to inhibit transcription of type III secretion operons. J. Biol. Chem. 2009, 284, 15762–15770. [Google Scholar] [CrossRef]
- Urbanowski, M.L.; Lykken, G.L.; Yahr, T.L. A secreted regulatory protein couples transcription to the secretory activity of the Pseudomonas aeruginosa type III secretion system. Proc. Natl. Acad. Sci. USA 2005, 102, 9930–9935. [Google Scholar] [CrossRef]
- Lykken, G.L.; Chen, G.Z.; Brutinel, E.D.; Chen, L.; Yahr, T.L. Characterization of ExsC and ExsD self-association and heterocomplex formation. J. Bacteriol. 2006, 188, 6832–6840. [Google Scholar] [CrossRef]
- Tessmer, M.H.; Anderson, D.M.; Buchaklian, A.; Frank, D.W.; Feix, J.B. Cooperative substrate-cofactor interactions and membrane localization of the bacterial phospholipase A 2 (PLA 2 ) enzyme, ExoU. J. Biol. Chem. 2017, 292, 3411–3419. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.H.; Halavaty, A.S.; Kim, H.; Geissler, B.; Agard, M.; Satchell, K.J.; Cho, W.; Anderson, W.F.; Hauser, A.R. A novel phosphatidylinositol 4,5-bisphosphate binding domain mediates plasma membrane localization of ExoU and other patatin-like phospholipases. J. Biol. Chem. 2015, 290, 2919–2937. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.H.; Hauser, A.R. Phosphatidylinositol 4,5-bisphosphate is a novel coactivator of the Pseudomonas aeruginosa cytotoxin ExoU. Infect. Immun. 2013, 81, 2873–2881. [Google Scholar] [CrossRef]
- Wenk, M.R.; Lucast, L.; Di Paolo, G.; Romanelli, A.J.; Suchy, S.F.; Nussbaum, R.L.; Cline, G.W.; Shulman, G.I.; McMurray, W.; De Camilli, P. Phosphoinositide profiling in complex lipid mixtures using electrospray ionization mass spectrometry. Nat. Biotechnol. 2003, 21, 813–817. [Google Scholar] [CrossRef]
- Ferrell, J.E.; Huests, W.H. Phosphoinositide metabolism and the morphology of human erythrocytes. J. Cell Biol. 1984, 98, 1992–1998. [Google Scholar] [CrossRef]
- Raucher, D.; Stauffer, T.; Chen, W.; Shen, K.; Guo, S.; York, J.D.; Sheetz, M.P.; Meyer, T. Phosphatidylinositol 4,5-bisphosphate functions as a second messenger that regulates cytoskeleton-plasma membrane adhesion. Cell 2000, 100, 221–228. [Google Scholar] [CrossRef]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef]
- Saarikangas, J.; Zhao, H.; Lappalainen, P. Regulation of the actin cytoskeleton-plasma membrane interplay by phosphoinositides. Physiol. Rev. 2010, 90, 259–289. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Frank, D.W. Intoxication of host cells by the T3SS phospholipase ExoU: PI(4,5)P2-associated, cytoskeletal collapse and late phase membrane blebbing. PLoS ONE 2014, 9, e103127. [Google Scholar] [CrossRef]
- Zhang, A.; Veesenmeyer, J.L.; Hauser, A.R. Phosphatidylinositol 4,5-Bisphosphate-Dependent Oligomerization of the Pseudomonas aeruginosa Cytotoxin ExoU. Infect. Immun. 2018, 86, 2873–2881. [Google Scholar] [CrossRef] [PubMed]
- Rabin, S.D.P.; Veesenmeyer, J.L.; Bieging, K.T.; Hauser, A.R. A C-terminal domain targets the Pseudomonas aeruginosa cytotoxin ExoU to the plasma membrane of host cells. Infect. Immun. 2006, 74, 2552–2561. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Keshet, Y.; Seger, R. The MAP kinase signaling cascades: A system of hundreds of components regulates a diverse array of physiological functions. Methods Mol. Biol. 2010, 661, 3–38. [Google Scholar] [PubMed]
- Cuzick, A.; Stirling, F.R.; Lindsay, S.L.; Evans, T.J. The type III pseudomonal exotoxin U activates the c-Jun NH 2 -terminal kinase pathway and increases human epithelial interleukin-8 production. Infect. Immun. 2006, 74, 4104–4113. [Google Scholar] [CrossRef]
- McMorran, B.; Town, L.; Costelloe, E.; Palmer, J.; Engel, J.; Hume, D.; Wainwright, B. Effector ExoU from the type III secretion system is an important modulator of gene expression in lung epithelial cells in response to Pseudomonas aeruginosa infection. Infect. Immun. 2003, 71, 6035–6044. [Google Scholar] [CrossRef]
- Saliba, A.M.; Nascimento, D.O.; Silva, M.C.A.; Assis, M.C.; Gayer, C.R.M.; Raymond, B.; Coelho, M.G.P.; Marques, E.A.; Touqui, L.; Albano, R.M.; et al. Eicosanoid-mediated proinflammatory activity of pseudomonas aeruginosa ExoU. Cell. Microbiol. 2005, 7, 1811–1822. [Google Scholar] [CrossRef]
- Lee, V.T.; Pukatzki, S.; Sato, H.; Kikawada, E.; Kazimirova, A.A.; Huang, J.; Li, X.; Arm, J.P.; Frank, D.W.; Lory, S. Pseudolipasin A is a specific inhibitor for phospholipase A2 activity of Pseudomonas aeruginosa cytotoxin ExoU. Infect. Immun. 2007, 75, 1089–1098. [Google Scholar] [CrossRef]
- Nikolaou, A.; Kokotou, M.G.; Vasilakaki, S.; Kokotos, G. Small-molecule inhibitors as potential therapeutics and as tools to understand the role of phospholipases A 2. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2019, 1864, 941–956. [Google Scholar] [CrossRef]
- Bao, J.; Marathe, B.; Govorkova, E.A.; Zheng, J.J. Drug Repurposing Identifies Inhibitors of Oseltamivir-Resistant Influenza Viruses. Angew. Chemie-Int. Ed. 2016, 55, 3438–3441. [Google Scholar] [CrossRef]
- Kim, D.; Baek, J.; Song, J.; Byeon, H.; Min, H.; Min, K.H. Identification of arylsulfonamides as ExoU inhibitors. Bioorg. Med. Chem. Lett. 2014, 24, 3823–3825. [Google Scholar] [CrossRef]
- Leroux, A.E.; Schulze, J.O.; Biondi, R.M. AGC kinases, mechanisms of regulation and innovative drug development. Semin. Cancer Biol. 2018, 48, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tessmer, M.H.; Anderson, D.M.; Pickrum, A.M.; Riegert, M.O.; Moretti, R.; Meiler, J.; Feix, J.B.; Frank, D.W. Identification of a ubiquitin-binding interface using Rosetta and DEER. Proc. Natl. Acad. Sci. USA 2018, 115, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.A.; Thomas, P.G.; Lindsley, C.W. Targeting phospholipase D in cancer, infection and neurodegenerative disorders. Nat. Rev. Drug Discov. 2017, 16, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, C.W.; Zhao, Z.; Leister, W.H.; Robinson, R.G.; Barnett, S.F.; Defeo-Jones, D.; Jones, R.E.; Hartman, G.D.; Huff, J.R.; Huber, H.E.; et al. Allosteric Akt (PKB) inhibitors: Discovery and SAR of isozyme selective inhibitors. Bioorganic Med. Chem. Lett. 2005, 15, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Barnett, S.F.; Defeo-Jones, D.; Fu, S.; Hancock, P.J.; Haskell, K.M.; Jones, R.E.; Kahana, J.A.; Kral, A.M.; Leander, K.; Lee, L.L.; et al. Identification and characterization of pleckstrin-homology-domain-dependent and isoenzyme-specific Akt inhibitors. Biochem. J. 2005, 385, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Schein, C.H. Repurposing approved drugs on the pathway to novel therapies. Med. Res. Rev. 2019. [Google Scholar] [CrossRef]
- Barker, W.; Nemeth, A.; Brackett, S.; Basak, A.; Chandler, C.E.; Jania, L.; Zuercher, W.; Melander, R.J.; Koller, B.; Ernst, R.K.; et al. Repurposing Eukaryotic Kinase Inhibitors as Colistin Adjuvants in Gram-negative Bacteria. ACS Infect. Dis. 2019, 5, 1764–1771. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virulence Factor | Category | Activity | Function | Ref. |
|---|---|---|---|---|
| Alginate | Extracellular polysaccharide | Biofilm formation | Contributes to biofilm formation and reduces susceptibility to antibiotics. | [15,16] |
| Alkaline protease (ArpA) | Exoenzyme | Zinc-dependent metalloprotease | Degrades host immune complements C1q, C2, and C3 and cytokines interferon (IFN)-γ and tumour necrosis factor (TNF)-α. | [17,18,19] |
| Cystic fibrosis transmembrane conductance regulator (CFTR) inhibitory factor (Cif) | Phenazine | Epoxide hydrolase | Promotes sustained inflammation by hydrolysing the paracrine signal 14,15-epoxyeicosatrienoic acid that stimulates neutrophils to produce the pro-resolving lipid mediator 15-epi lipoxin A4. Cif increases the ubiquitination and lysosomal degradation of some ATP-binding cassette transporters (ABC) including CFTR, P-glycoprotein, and TAP1. | [20,21,22,23] |
| ExoA | Exoenzyme | Catalytic ADP-ribosylation of elongation factor 2 | Inhibits protein synthesis and induces apoptosis in the host cell. | [24] |
| ExoS | Exoenzyme | Bifunctional toxin with Rho GTPase-activating protein (RhoGAP) activity and ADP-ribosyltransferase (ADPRT) activity | It blocks the reactive oxygen species burst in neutrophils by ADP-ribosylation of Ras, thereby preventing the activation of phosphoinositide-3-kinase (PI3K), which is required to stimulate the phagocytic NADPH-oxidase. | [25,26] |
| ExoT | Exoenzyme | Bifunctional toxin with RhoGAP activity and ADPRT activity | It impairs the production of reactive oxygen species burst in neutrophils and promotes the apoptosis of host cells by transforming host protein Crk by ADP-ribosylation into a cytotoxin and by activation of the intrinsic mitochondrial apoptotic pathway. | [27,28] |
| ExoU | Exoenzyme | Phospholipase A2 | It becomes activated by interaction with ubiquitin or ubiquitinylated proteins in the cytosol of the host cell before localising to the cell membrane to catalyse fatty acids from a broad range of phospholipids and lysophospholipids. | [29,30] |
| ExoY | Exoenzyme | Secreted adenyl cyclase | Increases concentration of intracellular cAMP in host cells through disruption of the actin cytoskeleton and increased endothelial permeability. | [28] |
| Flagella | Organelle | Motility and adherence to surfaces | Elicits strong NFκB-mediated inflammatory response via signalling through toll-like receptor (TLR) 5 and a caspase-1-mediated response through Nod-like receptor, Ipaf. Provides bacterium with swimming motility in liquid. | [31] |
| LasA | Exoenzyme | Metallopeptidase, also known as staphylolysin | LasA acts with restricted specificity, predominantly at glycine-glycine peptide bonds, but also increases the elastinolytic activity of LasB. | [32] |
| LasB | Exoenzyme | Zinc-metalloprotease | Causes elastin degradation. | [17,33] |
| PlcH | Exoenzyme | Haemolytic phospholipase C | Releases phosphate esters from sphingomyelin and phosphatidylcholine. | [34] |
| PlcN | Exoenzyme | Non-haemolytic phospholipase C | Releases phosphate esters from phosphatidylserine and phosphatidylcholine. | [35] |
| PldA and PldB | Exoenzyme | Phospholipase D | Facilitates intracellular invasion of host eukaryotic cells by activation of the PI3K/ Akt pathway. | [36,37] |
| PrpL | Exoenzyme | Class IV protease, lysine endoproteinase | Inactivates a range of host defences including fibrinogen, plasminogen, immunoglobulin G, and complement proteins C1q and C3. | [38,39] |
| Pyocyanin | Phenazine | Redox-active zwitterion | Inhibits host cell respiration, ciliary function, and epidermal growth; disrupts calcium homeostasis and induces apoptosis in neutrophils. | [40] |
| Rhamnolipids | Surfactant | Biosurfactants | Participates in the maintenance of uninhabited channels surrounding biofilm communities, which serve to provide nutrients and oxygen to the colonies of bacteria. Biofilms can form on implants and on dead or living tissue. They are inherently difficult to eradicate with antibiotics due to the inability of antibiotic molecules to penetrate the extracellular matrix. | [41] |
| TplE | Exoenzyme | Phospholipase A1 | Disrupts the endoplasmic reticulum and thereby promotes autophagy by the activation of the unfolded protein response. | [42] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foulkes, D.M.; McLean, K.; Haneef, A.S.; Fernig, D.G.; Winstanley, C.; Berry, N.; Kaye, S.B. Pseudomonas aeruginosa Toxin ExoU as a Therapeutic Target in the Treatment of Bacterial Infections. Microorganisms 2019, 7, 707. https://doi.org/10.3390/microorganisms7120707
Foulkes DM, McLean K, Haneef AS, Fernig DG, Winstanley C, Berry N, Kaye SB. Pseudomonas aeruginosa Toxin ExoU as a Therapeutic Target in the Treatment of Bacterial Infections. Microorganisms. 2019; 7(12):707. https://doi.org/10.3390/microorganisms7120707
Chicago/Turabian StyleFoulkes, Daniel M., Keri McLean, Atikah S. Haneef, David G. Fernig, Craig Winstanley, Neil Berry, and Stephen B. Kaye. 2019. "Pseudomonas aeruginosa Toxin ExoU as a Therapeutic Target in the Treatment of Bacterial Infections" Microorganisms 7, no. 12: 707. https://doi.org/10.3390/microorganisms7120707
APA StyleFoulkes, D. M., McLean, K., Haneef, A. S., Fernig, D. G., Winstanley, C., Berry, N., & Kaye, S. B. (2019). Pseudomonas aeruginosa Toxin ExoU as a Therapeutic Target in the Treatment of Bacterial Infections. Microorganisms, 7(12), 707. https://doi.org/10.3390/microorganisms7120707