Quantitative and Compositional Study of Monospecies Biofilms of Spoilage Microorganisms in the Meat Industry and Their Interaction in the Development of Multispecies Biofilms

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Surfaces to Test
2.2. Bacterial Strains
2.3. Monospecies Biofilms
2.3.1. Inoculum Preparation
2.3.2. Biofilm Formation
2.3.3. Evaluation of Cell Viability and Matrix Composition of the Biofilms by Direct Epifluorescent Microscopy (DEM)
2.4. L. gasicomitatum Preimplantation on Stainless Steel Surfaces and Its Effect on Subsequent Colonization by P. fragi and L. reuteri
2.4.1. Inoculum Preparation
2.4.2. L. gasicomitatum Preimplantation and Subsequent Colonization
2.4.3. Multispecies Biofilm Evaluation by Plate Count
2.5. Statistical Analysis
3. Results and Discussion
3.1. Evaluation of the Formation Capacity of the Monospecies Biofilms
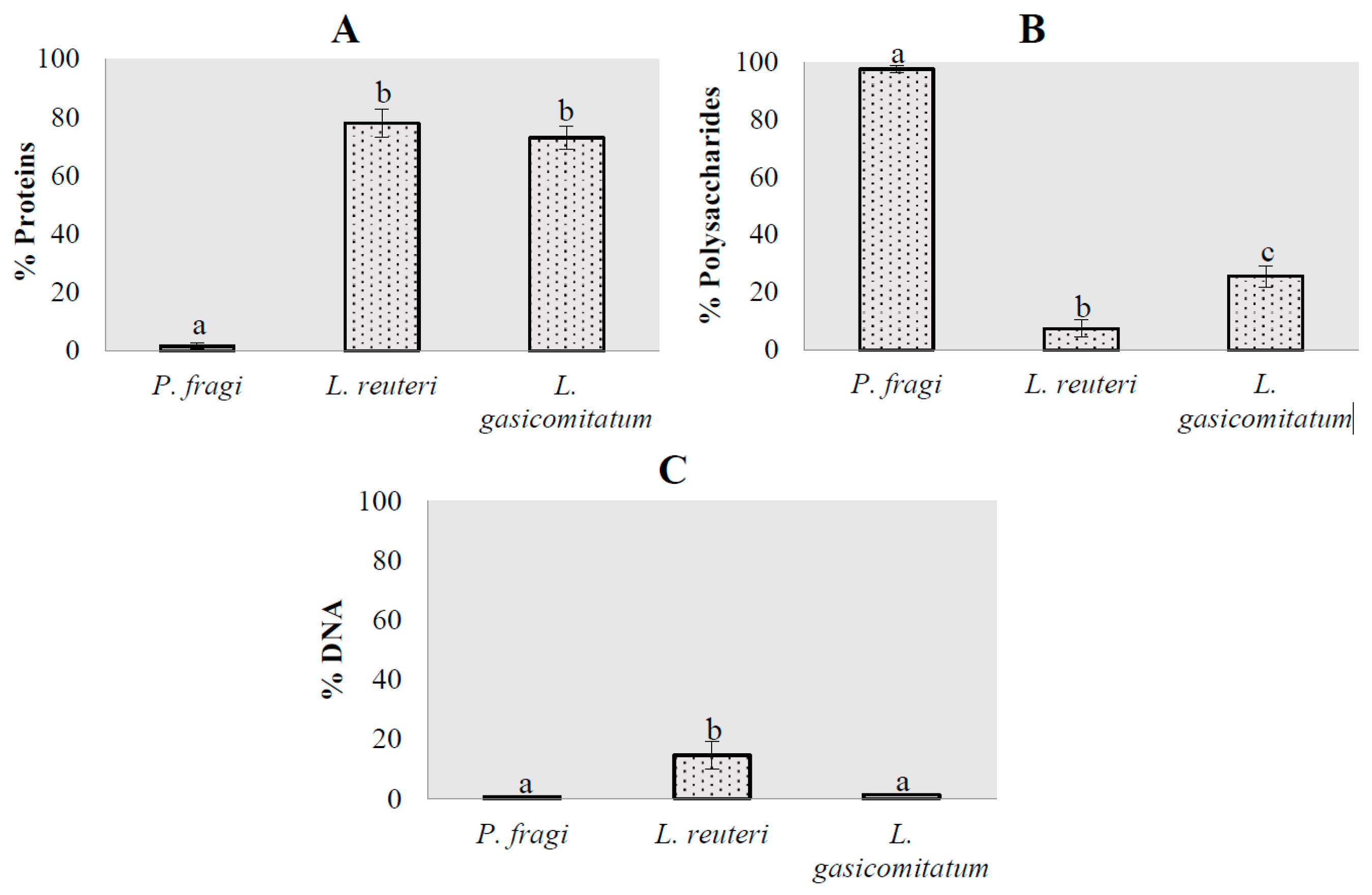
3.2. Matrix Composition in Monospecies Biofilms
3.3. Preimplantation of L. gasicomitatum and Influence on the Growth of P. fragi and L. reuteri
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khelissa, S.O.; Abdallah, M.; Jama, C.; Faille, C.; Chihib, N.E. Bacterial contamination and biofilm formation on abiotic surfaces and strategies to overcome their persistence. J. Mater. Environ. Sci. 2017, 8, 3326–3346. [Google Scholar]
- Desmond, P.; Best, J.P.; Morgenroth, E.; Derlon, N. Linking composition of extracellular polymeric substances (EPS) to the physical structure and hydraulic resistance of membrane biofilms. Water Res. 2018, 132, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Ripolles-Avila, C.; Ríos-Castillo, A.G.; Rodríguez-Jerez, J.J. Development of a peroxide biodetector for a direct detection of biofilms produced by catalase-positive bacteria on food-contact surfaces. CYTA-J. Food 2018, 16, 506–515. [Google Scholar] [CrossRef]
- Winkelstroter, L.K. Microbial Biofilms: The challenge of food industry. Biochem. Mol. Biol. J. 2015, 1, 3–5. [Google Scholar] [CrossRef][Green Version]
- Percival, S.L.; McCarty, S.M.; Lipsky, B. Biofilms and wounds: An overview of the evidence. Adv. Wound Care 2015, 4, 373–381. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Hascoët, A.S.; Ríos-Castillo, A.G.; Rodríguez-Jerez, J.J. Hygienic properties exhibited by single-use wood and plastic packaging on the microbial stability for fish. LWT-Food Sci. Technol. 2019, 113, 108309. [Google Scholar] [CrossRef]
- González-Rivas, F.; Ripolles-Avila, C.; Fontecha-Umaña, F.; Ríos-Castillo, A.G.; Rodríguez-Jerez, J.J. Biofilms in the spotlight: Detection, quantification, and removal methods. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1261–1276. [Google Scholar] [CrossRef]
- Stoodley, P.; Hall-Stoodley, L.; Costerton, B.; DeMeo, P.; Shirtliff, M.; Gawalt, E.; Kathju, S. Biofilms, Biomaterials, and Device-Related Infections; Elsevier Inc.: Amsterdam, Netherlands, 2013. [Google Scholar]
- Flemming, H.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef]
- Hobley, L.; Harkins, C.; MacPhee, C.E.; Stanley-Wall, N.R. Giving structure to the biofilm matrix: An overview of individual strategies and emerging common themes. FEMS Microbiol. Rev. 2015, 39, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Borlee, B.R.; Goldman, A.D.; Murakami, K.; Samudrala, R.; Wozniak, D.J.; Parsek, M.R. Pseudomonas aeruginosa uses a cyclic-di-GMP-regulated adhesin to reinforce the biofilm extracellular matrix. Mol. Microbiol. 2010, 75, 827–842. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, L.M.; Cotter, P.D.; Hill, C.; Alvarez-Ordóñez, A. New weapons to fight old enemies: Novel strategies for the (bio)control of bacterial biofilms in the food industry. Front. Microbiol. 2016, 7, 1641. [Google Scholar] [CrossRef]
- Molina-González, D.; Alonso-Calleja, C.; Alonso-Hernando, A.; Capita, R. Effect of sub-lethal concentrations of biocides on the susceptibility to antibiotics of multi-drug resistant Salmonella enterica strains. Food Control 2014, 40, 329–334. [Google Scholar] [CrossRef]
- Rodríguez-Melcón, C.; Capita, R.; Rodríguez-Jerez, J.J.; Martínez-Suárez, J.V.; Alonso-Calleja, C. Effect of low doses of disinfectants on the biofilm-forming ability of Listeria monocytogenes. Foodborne Pathog. Dis. 2019, 16, 262–268. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; García, P. Bacteriophages as weapons against bacterial biofilms in the food industry. Front. Microbiol. 2016, 7, 825. [Google Scholar] [CrossRef]
- Oh, H.S.; Tan, C.H.; Low, J.H.; Rzechowicz, M.; Siddiqui, M.F.; Winters, H.; Kjelleberg, S.; Fane, A.G.; Rice, S.A. Quorum quenching bacteria can be used to inhibit the biofouling of reverse osmosis membranes. Water Res. 2017, 112, 29–37. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Wu, X.; Sun, Y.; Liu, Z. Effect of thyme essential oil against Bacillus cereus planktonic growth and biofilm formation. Appl. Microbiol. Biotechnol. 2018, 102, 10209–10218. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Hascoët, A.S.; Martínez-Suárez, J.V.; Capita, R.; Rodríguez-Jerez, J.J. Evaluation of the microbiological contamination of food processing environments through implementing surface sensors in an iberian pork processing plant: An approach towards the control of Listeria monocytogenes. Food Control 2019, 99, 40–47. [Google Scholar] [CrossRef]
- AECOC. Informe|Congreso AECOC de Productos Cárnicos y Elaborados. Revista AECOC. 2018. Available online: https://www.aecoc.es/libros/congreso-aecoc-de-productos-carnicos-y-elaborados/ (accessed on 21 October 2019).
- Iulietto, M.F.; Sechi, P.; Borgogni, E.; Cenci-Goga, B.T. Meat spoilage: A critical review of a neglected alteration due to ropy slime producing bacteria. Ital. J. Anim. Sci. 2015, 14, 316–326. [Google Scholar] [CrossRef]
- Ercolini, D.; Ferrocino, I.; Nasi, A.; Ndagijimana, M.; Vernocchi, P.; La Storia, A.; Laghi, L.; Mauriello, G.; Guerzoni, M.E.; Villani, F. Monitoring of microbial metabolites and bacterial diversity in beef stored under different packaging londitions. Appl. Environ. Microbiol. 2011, 77, 7372–7381. [Google Scholar] [CrossRef] [PubMed]
- Stellato, G.; Utter, D.R.; Voorhis, A.; De Angelis, M.; Murat Eren, A.; Ercolini, D. A few Pseudomonas oligotypes dominate in the meat and dairy processing environment. Front. Microbiol. 2017, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Stanborough, T.; Fegan, N.; Powell, S.M.; Singh, T.; Tamplin, M.; Chandry, P.S. Genomic and metabolic characterization of spoilage-associated Pseudomonas species. Int. J. Food Microbiol. 2018, 268, 61–72. [Google Scholar] [CrossRef]
- Casaburi, A.; Piombino, P.; Nychas, G.J.; Villani, F.; Ercolini, D. Bacterial populations and the volatilome associated to meat spoilage. Food Microbiol. 2015, 45, 83–102. [Google Scholar] [CrossRef]
- Castellano, P.; Pérez Ibarreche, M.; Blanco Massani, M.; Fontana, C.; Vignolo, G. Strategies for pathogen biocontrol using lactic acid bacteria and their metabolites: A focus on meat ecosystems and industrial environments. Microorganisms 2017, 5, 38. [Google Scholar] [CrossRef]
- Wang, L.; Fang, M.; Hu, Y.; Yang, Y.; Yang, M.; Chen, Y. Characterization of the most abundant Lactobacillus species in chicken gastrointestinal tract and potential use as probiotics for genetic engineering. Acta Biochim. Biophys. Sin. 2014, 46, 612–619. [Google Scholar] [CrossRef]
- Hascoët, A.-S.; Ripolles-Avila, C.; Guerrero-Navarro, A.E.; Rodríguez-Jerez, J.J. Microbial ecology evaluation of an iberian pig processing plant through implementing SCH sensors and the influence of the resident microbiota on Listeria monocytogenes. Appl. Sci. 2019, 9, 4611. [Google Scholar] [CrossRef]
- Nieminen, T.T.; Koskinen, K.; Laine, P.; Hultman, J.; Säde, E.; Paulin, L.; Paloranta, A.; Johansson, P.; Björkroth, J.; Auvinen, P. Comparison of microbial communities in marinated and unmarinated broiler meat by metagenomics. Int. J. Food Microbiol. 2012, 157, 142–149. [Google Scholar] [CrossRef]
- Nieminen, T.T.; Välitalo, H.; Säde, E.; Paloranta, A.; Koskinen, K.; Björkroth, J. The effect of marination on lactic acid bacteria communities in raw broiler fillet strips. Front. Microbiol. 2012, 3, 376. [Google Scholar] [CrossRef]
- Pothakos, V.; Devlieghere, F.; Villani, F.; Björkroth, J.; Ercolini, D. Lactic acid bacteria and their controversial role in fresh meat spoilage. Meat Sci. 2015, 109, 66–74. [Google Scholar] [CrossRef]
- UNE-EN 13697. Chemical Disinfectants and Antiseptics—Quantitative Non-porous Surface Test for the Evaluation of Bactericidal and/or Fungicidal Activity of Chemical Disinfectants Used in Food, Industrial, Domestic and Institutional Areas—Test Method and Requirements without Mechanical Action (Phase 2, Step 2); AENOR: Madrid, Spain, 2015. [Google Scholar]
- Ripolles-Avila, C.; Hascoët, A.S.; Guerrero-Navarro, A.E.; Rodríguez-Jerez, J.J. Establishment of incubation conditions to optimize the in vitro formation of mature Listeria monocytogenes biofilms on food-contact surfaces. Food Control 2018, 92, 240–248. [Google Scholar] [CrossRef]
- Fuster-Valls, N.; Hernández-Herrero, M.; Marín-de-Mateo, M.; Rodríguez-Jerez, J.J. Effect of different environmental conditions on the bacteria survival on stainless steel surfaces. Food Control 2008, 19, 308–314. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Cervantes-Huaman, B.H.; Hascoët, A.S.; Yuste, J.; Rodríguez-Jerez, J.J. Quantification of mature Listeria monocytogenes biofilm cells formed by an in vitro model: A comparison of different methods. Int. J. Food Microbiol. 2019, 289, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.A. Bacterial biofilms in food processing environments: A review of recent developments in chemical and biological control. Int. J. Food Sci. Technol. 2016, 51, 1731–1743. [Google Scholar] [CrossRef]
- Garrett, T.R.; Bhakoo, M.; Zhang, Z. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Achinas, S.; Charalampogiannis, N.; Euverink, G.J.W. A brief recap of microbial adhesion and biofilms. Appl. Sci. 2019, 9, 2801. [Google Scholar] [CrossRef]
- Martinez-Martinez, L.; Pascual, A.; Perea, E.J. Kinetics of adherence of mucoid and non-mucoid Pseudomonas aeruginosa to plastic catheters. J. Med. Microbiol. 1991, 34, 7–12. [Google Scholar] [CrossRef]
- Asally, M.; Kittisopikul, M.; Rué, P.; Du, Y.; Hu, Z.; Çağatay, T.; Robinson, A.B.; Lu, H.; Garcia-Ojalvo, J.; Süel, G.M. Localized cell death focuses mechanical forces during 3D patterning in a biofilm. Proc. Natl. Acad. Sci. USA 2012, 109, 18891–18896. [Google Scholar] [CrossRef]
- Hartmann, R.; Singh, P.K.; Pearce, P.; Mok, R.; Song, B.; Díaz-Pascual, F.; Dunkel, J.; Drescher, K. Emergence of three-dimensional order and structure in growing biofilms. Nat. Phys. 2019, 15, 251–256. [Google Scholar] [CrossRef]
- Chambless, J.D.; Hunt, S.M.; Stewart, P.S.; Chambless, J.D.; Hunt, S.M.; Stewart, P.S. A three-dimensional computer model of four hypothetical mechanisms protecting biofilms from antimicrobials. Appl. Environ. Microbiol. 2013, 72, 2005–2013. [Google Scholar] [CrossRef]
- Chmielewski, R.A.N.; Frank, J.F. Biofilm formation and control in food processing facilities. Compr. Rev. Food Sci. Food Saf. 2003, 2, 22–32. [Google Scholar] [CrossRef]
- Guilbaud, M.; Piveteau, P.; Desvaux, M.; Brisse, S.; Briandet, R. Exploring the diversity of Listeria monocytogenes biofilm architecture by high-throughput confocal laser scanning microscopy and the predominance of the honeycomb-like morphotype. Appl. Environ. Microbiol. 2015, 81, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
- Marsh, E.J.; Luo, H.; Wang, H. A three-tiered approach to differentiate Listeria monocytogenes biofilm-forming abilities. FEMS Microbiol. Lett. 2003, 228, 203–210. [Google Scholar] [CrossRef]
- Dos Reis-Teixeira, F.B.; Alves, V.F.; de Martinis, E.C.P. Growth, viability and architecture of biofilms of Listeria monocytogenes formed on abiotic surfaces. Braz. J. Microbiol. 2017, 48, 587–591. [Google Scholar] [CrossRef]
- Yan, J.; Bassler, B.L. Surviving as a community: Antibiotic tolerance and persistence in bacterial biofilms. Cell Host Microbe. 2019, 26, 15–21. [Google Scholar] [CrossRef]
- Colagiorgi, A.; Di Ciccio, P.; Zanardi, E.; Ghidini, S.; Ianieri, A. A look inside the Listeria monocytogenes biofilms extracellular matrix. Microorganisms 2016, 4, 22. [Google Scholar] [CrossRef]
- Combrouse, T.; Sadovskaya, I.; Faille, C.; Kol, O.; Guérardel, Y.; Midelet-Bourdin, G. Quantification of the extracellular matrix of the Listeria monocytogenes biofilms of different phylogenic lineages with optimization of culture conditions. J. Appl. Microbiol. 2013, 114, 1120–1131. [Google Scholar] [CrossRef]
- Colagiorgi, A.; Bruini, I.; Di Ciccio, P.A.; Zanardi, E.; Ghidini, S.; Ianieri, A. Listeria monocytogenes biofilms in the wonderland of food industry. Pathogens. 2017, 6, 41. [Google Scholar] [CrossRef]
- Nouha, K.; Kumar, R.S.; Balasubramanian, S.; Tyagi, R.D. Critical review of EPS production, synthesis and composition for sludge flocculation. J. Environ. Sci. 2017, 66, 225–245. [Google Scholar] [CrossRef]
- Peterson, C.P.; Sauer, C.; Chatfield, C.H. The extracellular polymeric substances of Legionella pneumophila biofilms contain amyloid structures. Curr. Microbiol. 2018, 75, 736–744. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H.; Xing, T.; Wu, N.; Xu, X.; Zhou, G. Removal of Salmonella biofilm formed under meat processing environment by surfactant in combination with bio-enzyme. LWT-Food Sci. Technol. 2016, 66, 298–304. [Google Scholar] [CrossRef]
- Zetzmann, M.; Okshevsky, M.; Endres, J.; Sedlag, A.; Caccia, N.; Auchter, M.; Waidmann, M.S.; Desvaux, M.; Meyer, R.L.; Riedel, C.U. DNase-sensitive and -resistant modes of biofilm formation by Listeria monocytogenes. Front. Microbiol. 2015, 6, 1428. [Google Scholar] [CrossRef] [PubMed]
- Bridier, A.; Sanchez-Vizuete, P.; Guilbaud, M.; Piard, J.C.; Naïtali, M.; Briandet, R. Biofilm-associated persistence of food-borne pathogens. Food Microbiol. 2015, 45, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Hultman, J.; Rahkila, R.; Ali, J.; Rousu, J.; Björkroth, K.J. Meat processing plant microbiome and contamination patterns of cold-tolerant bacteria causing food safety and spoilage risks in the manufacture of vacuum-packaged cooked sausages. Appl. Environ. Microbiol. 2015, 81, 7088–7097. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Frausto, J.J.; Cepeda-Marquez, L.G.; Salgado, L.M.; Iturriaga, M.H.; Arvizu-Medrano, S.M. Detection and genotyping of Leuconostoc spp. in a sausage processing plant. J. Food Prot. 2015, 78, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.K.; Rainey, P.B.; Haagensen, J.A.J.; Molin, S. Evolution of species interactions in a biofilm community. Nature. 2007, 445, 533–536. [Google Scholar] [CrossRef]
- Burmølle, M.; Ren, D.; Bjarnsholt, T.; Sørensen, S.J. Interactions in multispecies biofilms: Do they actually matter? Trends Microbiol. 2014, 22, 84–91. [Google Scholar] [CrossRef]
- Ren, D.; Madsen, J.S.; de la Cruz-Perera, C.I.; Bergmark, L.; Sørensen, S.J.; Burmølle, M. High-throughput screening of multispecies biofilm formation and quantitative PCR-based assessment of individual species proportions, useful for exploring interspecific bacterial interactions. Microb. Ecol. 2014, 68, 146–154. [Google Scholar] [CrossRef]
- Iñiguez-Moreno, M.; Gutiérrez-Lomelí, M.; Avila-Novoa, M.G. Kinetics of biofilm formation by pathogenic and spoilage microorganisms under conditions that mimic the poultry, meat, and egg processing industries. Int. J. Food Microbiol. 2019, 303, 32–41. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Microorganism | Total Count (Log CFU/cm2) | % Survival * |
|---|---|---|
| P. fragi | 4.82 ± 0.12 a | 0.03 ± 0.02 a |
| L. reuteri | 7.10 ± 0.05 b | 10.35 ± 5.85 a |
| L. gasicomitatum | 7.05 ± 0.26 b | 0.92 ± 0.87 a |
| Bacteria | Incubation Period as Multispecies Biofilm (Hours) | ||
|---|---|---|---|
| 24 | 48 | 72 | |
| P. fragi | 3.69 ± 0.66 aA | 4.73 ± 0.33 aA | 4.46 ± 0.29 aA |
| L. gasicomitatum | 5.98 ± 0.22 bA | 6.10 ± 0.18 bA | 5.76 ± 0.22 bA |
| Control L. gasicomitatum | 5.73 ± 0.16 bA | 6.17 ± 0.15 bA | 5.76 ± 0.20 bA |
| Bacteria | Incubation Period as Multispecies Biofilm (Hours) | ||
|---|---|---|---|
| 24 | 48 | 72 | |
| L. reuteri | 5.42 ± 0.73 aA | 6.21 ± 0.17 aA | 4.63 ± 0.64 aA |
| L. gasicomitatum | 5.00 ± 0.67 aA | 6.32 ± 0.04 aA | 5.97 ± 0.13 bA |
| Control L. gasicomitatum | 6.72 ± 0.03 aA | 6.36 ± 0.09 aB | 6.03 ± 0.12 bB |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ripolles-Avila, C.; García-Hernández, N.; Cervantes-Huamán, B.H.; Mazaheri, T.; Rodríguez-Jerez, J.J. Quantitative and Compositional Study of Monospecies Biofilms of Spoilage Microorganisms in the Meat Industry and Their Interaction in the Development of Multispecies Biofilms. Microorganisms 2019, 7, 655. https://doi.org/10.3390/microorganisms7120655
Ripolles-Avila C, García-Hernández N, Cervantes-Huamán BH, Mazaheri T, Rodríguez-Jerez JJ. Quantitative and Compositional Study of Monospecies Biofilms of Spoilage Microorganisms in the Meat Industry and Their Interaction in the Development of Multispecies Biofilms. Microorganisms. 2019; 7(12):655. https://doi.org/10.3390/microorganisms7120655
Chicago/Turabian StyleRipolles-Avila, Carolina, Nerea García-Hernández, Brayan H. Cervantes-Huamán, Tina Mazaheri, and José Juan Rodríguez-Jerez. 2019. "Quantitative and Compositional Study of Monospecies Biofilms of Spoilage Microorganisms in the Meat Industry and Their Interaction in the Development of Multispecies Biofilms" Microorganisms 7, no. 12: 655. https://doi.org/10.3390/microorganisms7120655
APA StyleRipolles-Avila, C., García-Hernández, N., Cervantes-Huamán, B. H., Mazaheri, T., & Rodríguez-Jerez, J. J. (2019). Quantitative and Compositional Study of Monospecies Biofilms of Spoilage Microorganisms in the Meat Industry and Their Interaction in the Development of Multispecies Biofilms. Microorganisms, 7(12), 655. https://doi.org/10.3390/microorganisms7120655