What Happens in the Staphylococcal Nucleoid under Oxidative Stress?

 , , ,
, , , 
Abstract
1. Introduction
2. Staphylococcus aureus Lifestyle and Importance of Oxidative Stress Resistance
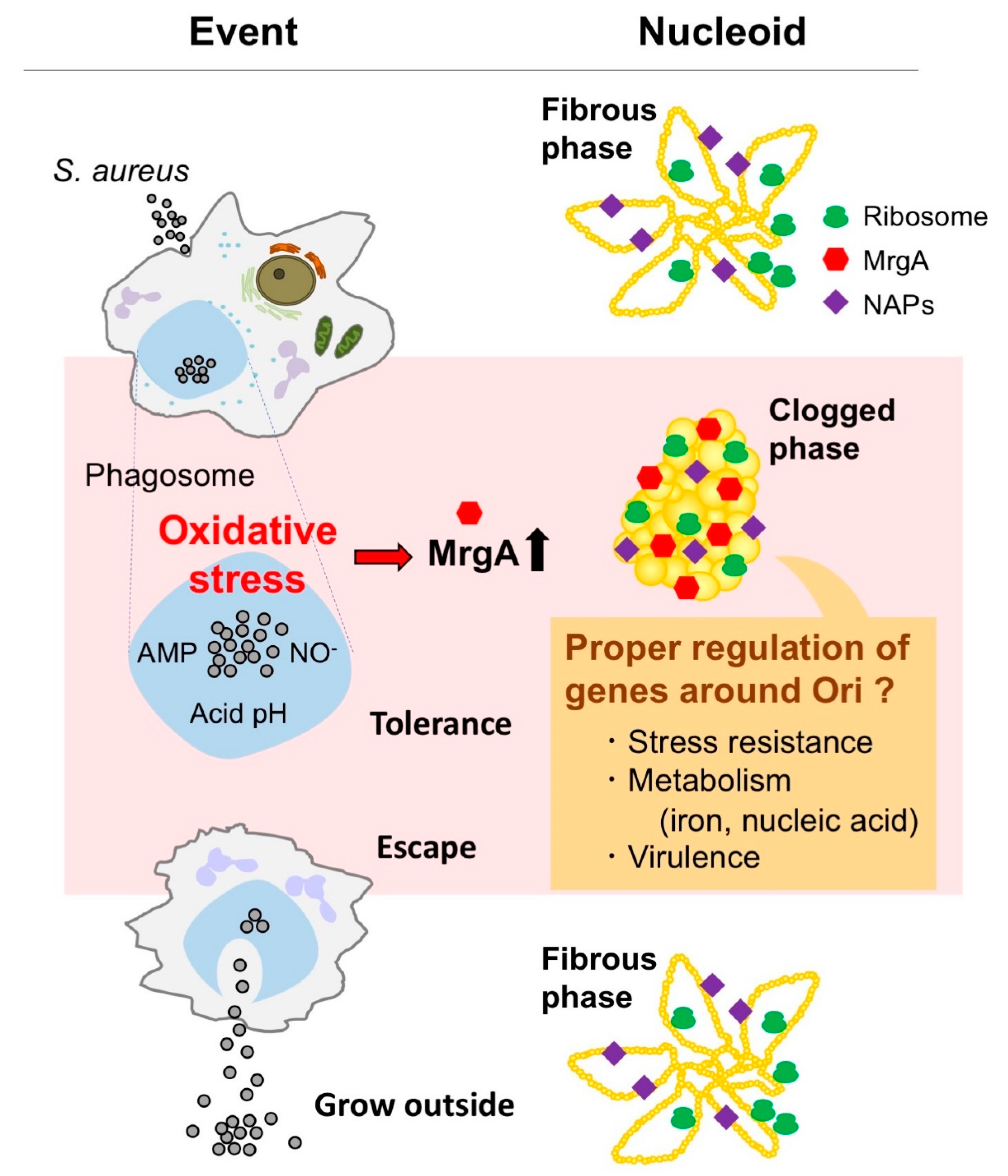
3. Oxidative Stress Induces Nucleoid Clogging
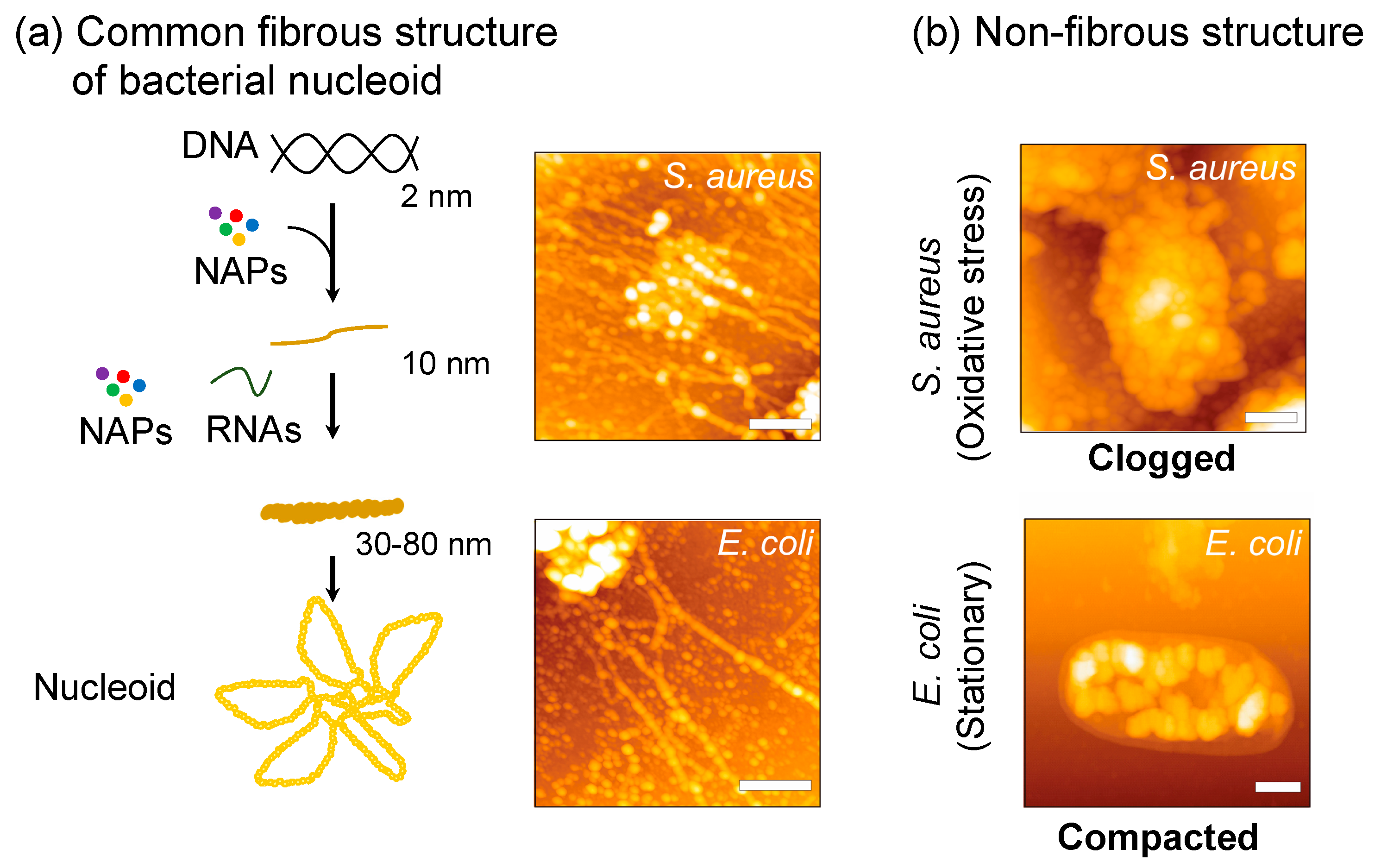
3.1. Characteristics of S. aureus Nucleoid in Comparison with Other Bacteria
3.2. Apparent Correlation between Nucleoid Clogging and Oxidative Stress
4. Is Nucleoid Clogging Required or Not for the Oxidative Stress Tolerance?
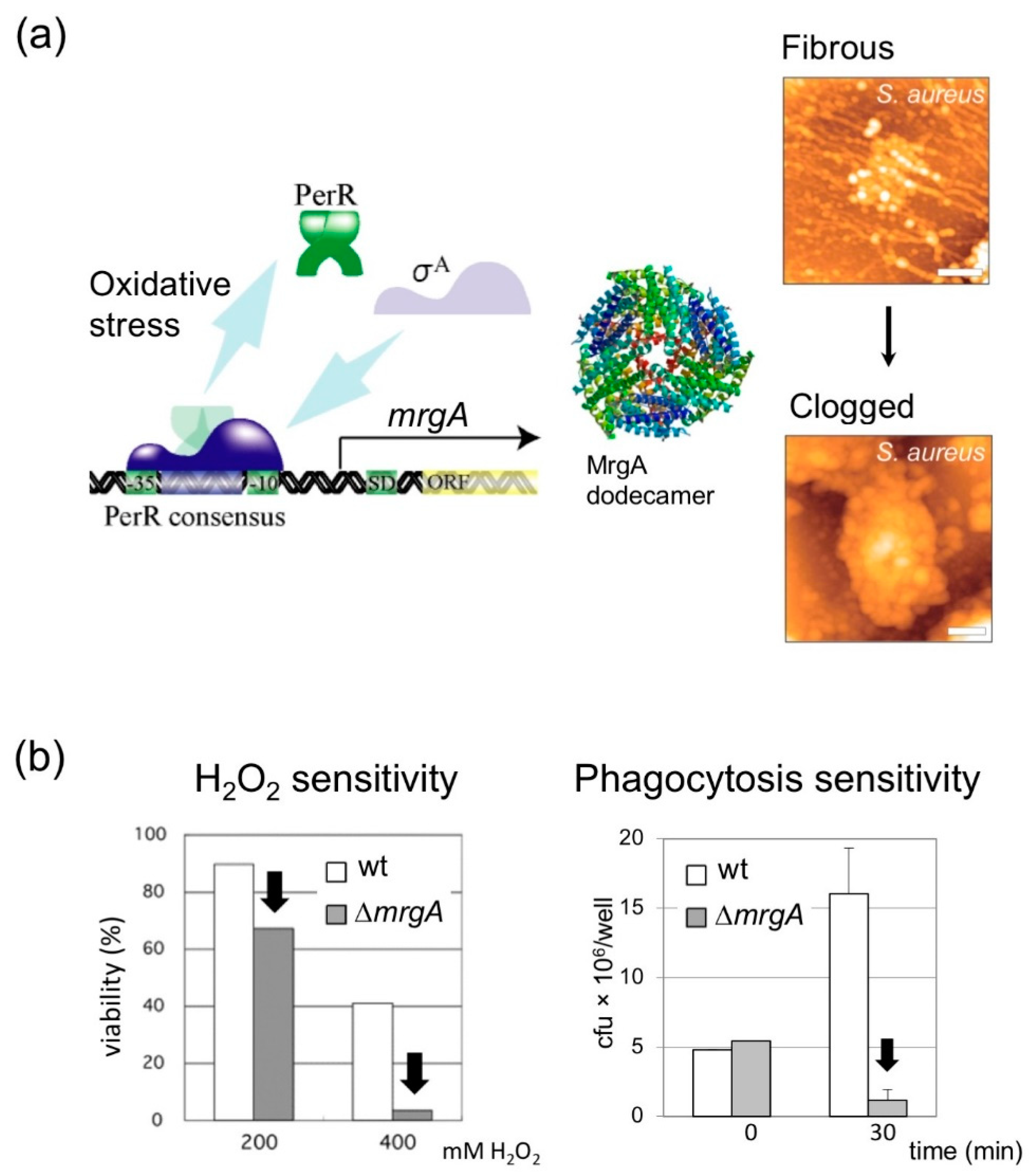
4.1. MrgA Is a Bifucntional Molecule with Ferroxidase Activity That Is Essential for Oxidative Stress Resistance
4.2. DNA Binding Activity of MrgA Is Dispensable for Hydrogen Peroxide Resistance and Survival in Phagosome, but Not for Nucleoid Clogging
5. Any Physiological Relevance in Nucleoid Clogging?
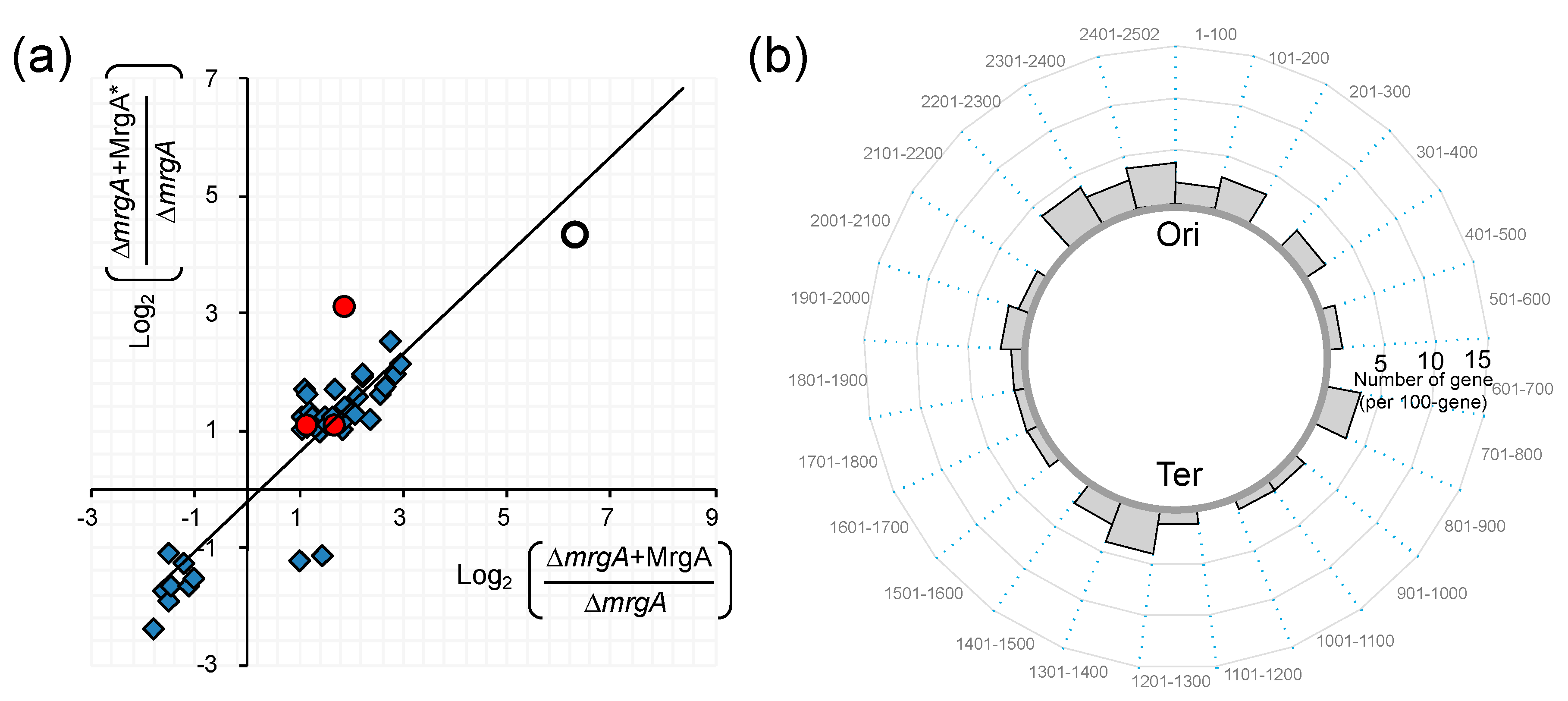
5.1. Characteristics of csNAPs in the Clogged and Relaxed Nucleoid
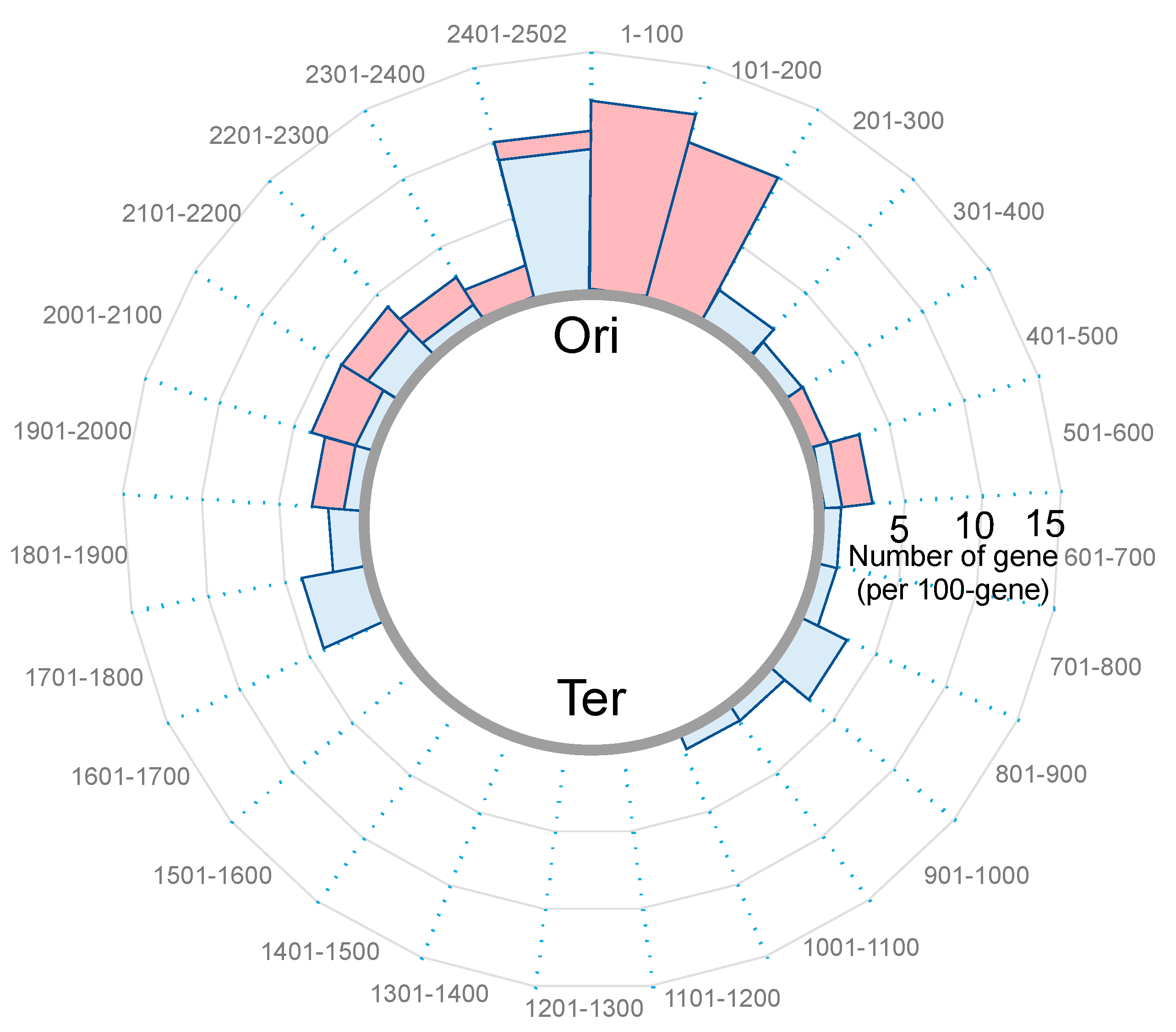
5.2. Effect of Nucleoid Clogging on Transcriptome Profile
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yoshimura, S.H.; Kim, J.; Takeyasu, K. On-substrate lysis treatment combined with scanning probe microscopy revealed chromosome structures in eukaryotes and prokaryotes. J. Electron. Microsc. (Tokyo) 2003, 52, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Ohniwa, R.L.; Morikawa, K.; Wada, C.; Ohta, T.; Takeyasu, K. Nucleoid architecture and dynamics in bacteria. In Bacterial DNA, DNA Polymerase and DNA Helicase; Knudsen, W.D., Bruns, S.S., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2010. [Google Scholar]
- Takeyasu, K.; Kim, J.; Ohniwa, R.L.; Kobori, T.; Inose, Y.; Morikawa, K.; Ohta, T.; Ishihama, A.; Yoshimura, S.H. Genome architecture studied by nanoscale imaging: Analyses among bacterial phyla and their implication to eukaryotic genome folding. Cytogenet. Genome Res. 2004, 107, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Ohniwa, R.L.; Maruyama, H.; Morikawa, K.; Takeyasu, K. Atomic Force Microscopy Imaging and Analysis of Prokaryotic Genome Organization. Methods Mol. Biol. 2018, 1837, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Lamers, R.P.; Muthukrishnan, G.; Castoe, T.A.; Tafur, S.; Cole, A.M.; Parkinson, C.L. Phylogenetic relationships among Staphylococcus species and refinement of cluster groups based on multilocus data. BMC Evol. Biol. 2012, 12, 171. [Google Scholar] [CrossRef]
- Watkins, R.R.; Holubar, M.; David, M.Z. Antimicrobial resistance in methicillin-resistant Staphylococcus aureus to newer antimicrobial agents. Antimicrob. Agents Chemother. 2019. [Google Scholar] [CrossRef]
- Morikawa, K.; Ohniwa, R.L.; Ohta, T.; Tanaka, Y.; Takeyasu, K.; Msadek, T. Adaptation beyond the Stress Response: Cell Structure Dynamics and Population Heterogeneity in Staphylococcus aureus. Microbes Environ. 2010, 25, 75–82. [Google Scholar] [CrossRef]
- Chaibenjawong, P.; Foster, S.J. Desiccation tolerance in Staphylococcus aureus. Arch. Microbiol. 2011, 193, 125–135. [Google Scholar] [CrossRef]
- Maudsdotter, L.; Imai, S.; Ohniwa, R.L.; Saito, S.; Morikawa, K. Staphylococcus aureus dry stress survivors have a heritable fitness advantage in subsequent dry exposure. Microbes Infect. 2015, 17, 456–461. [Google Scholar] [CrossRef]
- Tsai, M.; Ohniwa, R.L.; Kato, Y.; Takeshita, S.L.; Ohta, T.; Saito, S.; Hayashi, H.; Morikawa, K. Staphylococcus aureus requires cardiolipin for survival under conditions of high salinity. BMC Microbiol. 2011, 11, 13. [Google Scholar] [CrossRef]
- Thammavongsa, V.; Kim, H.K.; Missiakas, D.; Schneewind, O. Staphylococcal manipulation of host immune responses. Nat. Rev. Microbiol. 2015, 13, 529–543. [Google Scholar] [CrossRef]
- Foster, T.J. Immune evasion by staphylococci. Nat. Rev. Microbiol. 2005, 3, 948–958. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, F.R.; Allen, L.A.; Apicella, M.; Nauseef, W.M. NADPH oxidase activation and assembly during phagocytosis. J. Immunol. 1999, 163, 6732–6740. [Google Scholar] [PubMed]
- McCord, J.M.; Fridovich, I. Superoxide dismutase: The first twenty years (1968–1988). Free Radic. Biol. Med. 1988, 5, 363–369. [Google Scholar] [CrossRef]
- Tainer, J.A.; Getzoff, E.D.; Richardson, J.S.; Richardson, D.C. Structure and mechanism of copper, zinc superoxide dismutase. Nature 1983, 306, 284–287. [Google Scholar] [CrossRef]
- Luo, Y.; Han, Z.; Chin, S.M.; Linn, S. Three chemically distinct types of oxidants formed by iron-mediated Fenton reactions in the presence of DNA. Proc. Natl. Acad. Sci. USA 1994, 91, 12438–12442. [Google Scholar] [CrossRef]
- Henle, E.S.; Linn, S. Formation, prevention, and repair of DNA damage by iron/hydrogen peroxide. J. Biol. Chem. 1997, 272, 19095–19098. [Google Scholar] [CrossRef]
- Voyich, J.M.; Braughton, K.R.; Sturdevant, D.E.; Whitney, A.R.; Saïd-Salim, B.; Porcella, S.F.; Long, R.D.; Dorward, D.W.; Gardner, D.J.; Kreiswirth, B.N.; et al. Insights into mechanisms used by Staphylococcus aureus to avoid destruction by human neutrophils. J. Immunol. 2005, 175, 3907–3919. [Google Scholar] [CrossRef]
- Kubica, M.G.K.; Koziel, J.; Zarebski, M.; Richter, W.; Gajkowska, B.; Golda, A.; Maciag-Gudowska, A.; Brix, K.; Shaw, L.; Foster, T.; et al. A potential new pathway for Staphylococcus aureus dissemination: The silent survival of S. aureus phagocytosed by human monocyte-derived macrophages. PLoS ONE 2008, 3, e1409. [Google Scholar] [CrossRef]
- Horn, J.; Stelzner, K.; Rudel, T.; Fraunholz, M. Inside job: Staphylococcus aureus host-pathogen interactions. Int. J. Med. Microbiol. 2018, 308, 607–624. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide dismutases. Annu. Rev. Biochem. 1975, 44, 147–159. [Google Scholar] [CrossRef]
- Valderas, M.W.; Hart, M.E. Identification and characterization of a second superoxide dismutase gene (sodM) from Staphylococcus aureus. J. Bacteriol. 2001, 183, 3399–3407. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.E. Mechanisms of reaction of hemeproteins with oxygen and hydrogen peroxide in the oxidation of organic substrates. Pharm. Ther. 1980, 10, 171–189. [Google Scholar] [CrossRef]
- Ushijima, Y.; Ohniwa, R.L.; Maruyama, A.; Saito, S.; Tanaka, Y.; Morikawa, K. Nucleoid compaction by MrgA(Asp56Ala/Glu60Ala) does not contribute to staphylococcal cell survival against oxidative stress and phagocytic killing by macrophages. Fems. Microbiol. Lett. 2014, 360, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Robinow, C.; Kellenberger, E. The bacterial nucleoid revisited. Microbiol. Rev. 1994, 58, 211–232. [Google Scholar] [PubMed]
- Kavenoff, R.; Bowen, B.C. Electron microscopy of membrane-free folded chromosomes from Escherichia coli. Chromosoma 1976, 59, 89–101. [Google Scholar] [CrossRef]
- Kavenoff, R.; Ryder, O.A. Electron microscopy of membrane-associated folded chromosomes of Escherichia coli. Chromosoma 1976, 55, 13–25. [Google Scholar] [CrossRef]
- Sloof, P.; Maagdelijn, A.; Boswinkel, E. Folding of prokaryotic DNA. Isolation and characterization of nucleoids from Bacillus licheniformis. J. Mol. Biol. 1983, 163, 277–297. [Google Scholar] [CrossRef]
- Ohniwa, R.L.; Muchaku, H.; Saito, S.; Wada, C.; Morikawa, K. Atomic force microscopy analysis of the role of major DNA-binding proteins in organization of the nucleoid in Escherichia coli. PLoS ONE 2013, 8, e72954. [Google Scholar] [CrossRef]
- Beintema, J.J.; Kleineidam, R.G. The ribonuclease A superfamily: General discussion. Cell Mol. Life Sci. 1998, 54, 825–832. [Google Scholar] [CrossRef]
- Ohniwa, R.L.; Morikawa, K.; Takeshita, S.L.; Kim, J.; Ohta, T.; Wada, C.; Takeyasu, K. Transcription-coupled nucleoid architecture in bacteria. Genes. Cells 2007, 12, 1141–1152. [Google Scholar] [CrossRef]
- Ishihama, A. Prokaryotic genome regulation: A revolutionary paradigm. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2012, 88, 485–508. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rueda, E.; Collado-Vides, J. The repertoire of DNA-binding transcriptional regulators in Escherichia coli K-12. Nucleic Acids Res. 2000, 28, 1838–1847. [Google Scholar] [CrossRef] [PubMed]
- Azam, T.A.; Ishihama, A. Twelve species of the nucleoid-associated protein from Escherichia coli Sequence recognition specificity and DNA binding affinity. J. Biol. Chem. 1999, 274, 33105–33113. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Luisi, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol. 2011, 9, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Cech, G.M.; Szalewska-Palasz, A.; Kubiak, K.; Malabirade, A.; Grange, W.; Arluison, V.; Wegrzyn, G. The Escherichia Coli. Hfq Protein: An Unattended DNA-Transactions Regulator. Front. Mol. Biosci. 2016, 3, 36. [Google Scholar] [CrossRef] [PubMed]
- Malabirade, A.; Partouche, D.; El Hamoui, O.; Turbant, F.; Geinguenaud, F.; Recouvreux, P.; Bizien, T.; Busi, F.; Wien, F.; Arluison, V. Revised role for Hfq bacterial regulator on DNA topology. Sci. Rep. 2018, 8, 16792. [Google Scholar] [CrossRef]
- Ohniwa, R.L.; Ushijima, Y.; Saito, S.; Morikawa, K. Proteomic analyses of nucleoid-associated proteins in Escherichia coli, Pseudomonas aeruginosa, Bacillus subtilis, and Staphylococcus aureus. PLoS ONE 2011, 6, e19172. [Google Scholar] [CrossRef]
- Huntzinger, E.; Boisset, S.; Saveanu, C.; Benito, Y.; Geissmann, T.; Namane, A.; Lina, G.; Etienne, J.; Ehresmann, B.; Ehresmann, C.; et al. Staphylococcus aureus RNAIII and the endoribonuclease III coordinately regulate spa gene expression. EMBO J. 2005, 24, 824–835. [Google Scholar] [CrossRef]
- Bohn, C.; Rigoulay, C.; Bouloc, P. No detectable effect of RNA-binding protein Hfq absence in Staphylococcus aureus. BMC Microbiol. 2007, 7, 10. [Google Scholar] [CrossRef]
- Ushijima, Y.; Ohniwa, R.L.; Morikawa, K. Identification of nucleoid associated proteins (NAPs) under oxidative stress in Staphylococcus aureus. BMC Microbiol. 2017, 17, 207. [Google Scholar] [CrossRef]
- Morikawa, K.; Ohniwa, R.L.; Kim, J.; Maruyama, A.; Ohta, T.; Takeyasu, K. Bacterial nucleoid dynamics: Oxidative stress response in Staphylococcus aureus. Genes Cells 2006, 11, 409–423. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, Y.; Yoshida, O.; Villanueva, M.J.; Ohniwa, R.L.; Morikawa, K. Nucleoid clumping is dispensable for the Dps-dependent hydrogen peroxide resistance in Staphylococcus aureus. Microbiology 2016. [Google Scholar] [CrossRef] [PubMed]
- Azam, T.A.; Iwata, A.; Nishimura, A.; Ueda, S.; Ishihama, A. Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid. J. Bacteriol. 1999, 181, 6361–6370. [Google Scholar]
- Kim, J.; Yoshimura, S.H.; Hizume, K.; Ohniwa, R.L.; Ishihama, A.; Takeyasu, K. Fundamental structural units of the Escherichia coli nucleoid revealed by atomic force microscopy. Nucleic Acids Res. 2004, 32, 1982–1992. [Google Scholar] [CrossRef] [PubMed]
- Janissen, R.; Arens, M.M.A.; Vtyurina, N.N.; Rivai, Z.; Sunday, N.D.; Eslami-Mossallam, B.; Gritsenko, A.A.; Laan, L.; de Ridder, D.; Artsimovitch, I.; et al. Global DNA Compaction in Stationary-Phase Bacteria Does Not Affect Transcription. Cell 2018, 174, 1188–1199. [Google Scholar] [CrossRef]
- Ohniwa, R.L.; Morikawa, K.; Kim, J.; Ohta, T.; Ishihama, A.; Wada, C.; Takeyasu, K. Dynamic state of DNA topology is essential for genome condensation in bacteria. EMBO J. 2006, 25, 5591–5602. [Google Scholar] [CrossRef]
- Haikarainen, T.; Papageorgiou, A.C. Dps-like proteins: Structural and functional insights into a versatile protein family. Cell Mol. Life Sci. 2010, 67, 341–351. [Google Scholar] [CrossRef]
- Imlay, J.A. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Poole, L.B.; Hantgan, R.R.; Kamio, Y. An iron-binding protein, Dpr, from Streptococcus mutans prevents iron-dependent hydroxyl radical formation in vitro. J. Bacteriol. 2002, 184, 2931–2939. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Fukui, K.; Koujin, N.; Ohya, H.; Kimura, K.; Kamio, Y. Regulation of the intracellular free iron pool by Dpr provides oxygen tolerance to Streptococcus mutans. J. Bacteriol. 2004, 186, 5997–6002. [Google Scholar] [CrossRef]
- Talukder, A.; Ishihama, A. Growth phase dependent changes in the structure and protein composition of nucleoid in Escherichia coli. Sci. China Life Sci. 2015, 58, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Cui, L.; Oguchi, A.; Aoki, K.; Nagai, Y.; et al. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Nobre, L.S.; Saraiva, L.M. Effect of combined oxidative and nitrosative stresses on Staphylococcus aureus transcriptome. Appl. Microbiol. Biotechnol. 2013, 97, 2563–2573. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Fujii, S.; Yamano, S.; Cho, A.K.; Kamisuki, S.; Nakai, Y.; Sugawara, F.; Froines, J.R.; Kumagai, Y. An approach to evaluate two-electron reduction of 9,10-phenanthraquinone and redox activity of the hydroquinone associated with oxidative stress. Free Radic. Biol. Med. 2007, 43, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, A.; Kumagai, Y.; Morikawa, K.; Taguchi, K.; Hayashi, H.; Ohta, T. Oxidative-stress-inducible qorA encodes an NADPH-dependent quinone oxidoreductase catalysing a one-electron reduction in Staphylococcus aureus. Microbiology 2003, 149, 389–398. [Google Scholar] [CrossRef]
- Gaupp, R.; Ledala, N.; Somerville, G.A. Staphylococcal response to oxidative stress. Front. Cell Infect. Microbiol. 2012, 2, 33. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxidative Stress | |||
| ID | Gene | Annotation | emPAI |
| sau:SA1414 | rpsT | 30S ribosomal protein S20 | 3.45 |
| sau:SAS033 | rpmF | 50S ribosomal protein L32 | 3.34 |
| sau:SA0092 | hypothetical protein | 3.23 | |
| sau:SA2032 | rplR | 50S ribosomal protein L18 | 2.72 |
| sau:SA1663 | UPF0342 protein SA1663 | 1.82 | |
| sau:SA0093 | hypothetical protein | 1.55 | |
| sau:SA1504 | infC | Translation initiation factor IF-3 | 1 |
| sau:SA2043 | rpsS | 30S ribosomal protein S19 | 0.89 |
| sau:SA1074 | fabG | 3-oxoacyl-[acyl-carrier-protein] reductase | 0.72 |
| sau:SA1279 | gpsB | Cell cycle protein gpsB | 0.69 |
| sau:SA2022 | rplQ | 50S ribosomal protein L17 | 0.65 |
| sau:SA2062 | sarV | HTH-type transcriptional regulator sarV | 0.64 |
| sau:SA1404 | rpsU | 30S ribosomal protein S21 | 0.62 |
| sau:SA0133 | dra | Deoxyribose-phosphate aldolase | 0.59 |
| sau:SA0162 | aldA | Putative aldehyde dehydrogenase aldA | 0.58 |
| sau:SA0957 | UPF0637 protein SA0957 | 0.56 | |
| sau:SA1053 | rpoZ | DNA-directed RNA polymerase subunit omega | 0.51 |
| sau:SA0204 | acpD | FMN-dependent NADH-azoreductase | 0.49 |
| sau:SA0232 | lctE | L-lactate dehydrogenase 1 | 0.47 |
| sau:SA0307 | nanE | Putative N-acetylmannosamine-6-phosphate 2-epimerase | 0.43 |
| sau:SA1305 | hu | DNA-binding protein HU | 0.42 |
| sau:SA0365 | ahpF | Alkyl hydroperoxide reductase subunit F | 0.42 |
| sau:SA1922 | rpmE2 | 50S ribosomal protein L31 type B | 0.41 |
| sau:SA0366 | ahpC | Alkyl hydroperoxide reductase subunit C | 0.4 |
| sau:SA0367 | nfrA | NADPH-dependent oxidoreductase | 0.4 |
| sau:SA1081 | rpsP | 30S ribosomal protein S16 | 0.39 |
| sau:SA1471 | rpmA | 50S ribosomal protein L27 | 0.39 |
| sau:SA1116 | rpsO | 30S ribosomal protein S15 | 0.38 |
| sau:SA2036 | rplX | 50S ribosomal protein L24 | 0.34 |
| sau:SA0468 | hprT | Hypoxanthine-guanine phosphoribosyltransferase | 0.34 |
| sau:SA0478 | pdxT | Glutamine amidotransferase subunit pdxT | 0.32 |
| sau:SA0488 | syc | Cysteinyl-tRNA synthetase | 0.32 |
| sau:SA0573 | sarA | Transcriptional regulator sarA | 0.26 |
| sau:SA2029 | rplO | 50S ribosomal protein L15 | 0.25 |
| sau:SA2423 | clfB | Clumping factor B | 0.25 |
| sau:SA1901 | fabZ | (3R)-hydroxymyristoyl-[acyl-carrier-protein] dehydratase | 0.24 |
| sau:SA0512 | ilvE | Probable branched-chain-amino-acid aminotransferase | 0.24 |
| sau:SA0520 | sdrD | Serine-aspartate repeat-containing protein D | 0.24 |
| sau:SA0480 | ctsR | Transcriptional regulator ctsR | 0.22 |
| sau:SA1172 | guaC | GMP reductase | 0.22 |
| sau:SA0537 | thiD | Phosphomethylpyrimidine kinase | 0.22 |
| sau:SA0544 | UPF0447 protein MW0542; heme peroxidase | 0.22 | |
| sau:SA1583 | rot | HTH-type transcriptional regulator rot | 0.2 |
| sau:SA0772 | Y772 | UPF0337 protein SA0772 | 0.2 |
| sau:SA0818 | rocD | Ornithine aminotransferase 2 | 0.2 |
| sau:SA0977 | isdA | Iron-regulated surface determinant protein A | 0.2 |
| sau:SA0942 | def | Peptide deformylase | 0.19 |
| sau:SA1032 | sepF | Cell division protein sepF | 0.18 |
| sau:SA1468 | ruvA | Holliday junction ATP-dependent DNA helicase ruvA | 0.17 |
| sau:SA2046 | rplD | 50S ribosomal protein L4 | 0.17 |
| Log Phase | |||
| ID | Gene | Annotation | emPAI |
| sau:SA0944 | phdB | Pyruvate dehydrogenase E1 component subunit beta | 2.39 |
| sau:SA1414 | rpsT | 30S ribosomal protein S20 | 2.06 |
| sau:SA2033 | rplF | 50S ribosomal protein L6 | 1.91 |
| sau:SA0723 | clpP | ATP-dependent Clp protease proteolytic subunit | 1.67 |
| sau:SA0504 | rpsG | 30S ribosomal protein S7 | 1.2 |
| sau:SA1382 | sodA | Superoxide dismutase [Mn/Fe] 1 | 1.18 |
| sau:SA0729 | tpi | Triosephosphate isomerase | 1.18 |
| sau:SA1663 | UPF0342 protein SA1663 | 1.17 | |
| sau:SA0366 | ahpC | Alkyl hydroperoxide reductase subunit C | 0.95 |
| sau:SA0456 | spoVG | Putative septation protein spoVG | 0.83 |
| sau:SA2036 | rplX | 50S ribosomal protein L24 | 0.81 |
| sau:SA1073 | fabD | Malonyl CoA-acyl carrier protein transacylase | 0.7 |
| sau:SA1930 | rpoE | Probable DNA-directed RNA polymerase subunit delta | 0.66 |
| sau:SA1113 | rbfA | Ribosome-binding factor A | 0.66 |
| sau:SA2312 | ddh | D-lactate dehydrogenase | 0.63 |
| sau:SA1404 | rpsU | 30S ribosomal protein S21 | 0.62 |
| sau:SA0856 | spxA | Regulatory protein spx | 0.56 |
| sau:SA2029 | rplO | 50S ribosomal protein L15 | 0.56 |
| sau:SA1901 | fabZ | (3R)-hydroxymyristoyl-[acyl-carrier-protein] dehydratase | 0.53 |
| sau:SA0719 | trxB | Thioredoxin reductase | 0.53 |
| sau:SA2039 | rpmC | 50S ribosomal protein L29 | 0.51 |
| sau:SA2026 | infA | Translation initiation factor IF-1 | 0.49 |
| sau:SA0245 | ispD | 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase 2 | 0.49 |
| sau:SA0918 | purC | Phosphoribosylaminoimidazole-succinocarboxamide synthase | 0.49 |
| sau:SA0941 | UPF0356 protein SA0941 | 0.46 | |
| sau:SA0354 | rpsR | 30S ribosomal protein S18 | 0.43 |
| sau:SA1653 | traP | Signal transduction protein TRAP | 0.43 |
| sau:SA1305 | hu | DNA-binding protein HU | 0.42 |
| sau:SA1359 | EF-P | Elongation factor P | 0.41 |
| sau:SA0942 | pdf1 | Peptide deformylase | 0.41 |
| sau:SAS074 | UPF0457 protein SA1975 | 0.4 | |
| sau:SA1081 | rpsP | 30S ribosomal protein S16 | 0.39 |
| sau:SA2043 | rpsS | 30S ribosomal protein S19 | 0.38 |
| sau:SA2399 | Fructose-bisphosphate aldolase class 1 | 0.38 | |
| sau:SA0707 | Uncharacterized protein SAB0704 | 0.37 | |
| sau:SA0128 | sodM | Superoxide dismutase [Mn/Fe] 2 | 0.36 |
| sau:SA1717 | gatC | tRNA(Asn/Gln) amidotransferase subunit C | 0.36 |
| sau:SA0352 | rpsF | 30S ribosomal protein S6 | 0.34 |
| sau:SA0855 | trpS | Tryptophanyl-tRNA synthetase | 0.34 |
| sau:SA0437 | UPF0133 protein SAB0428 | 0.34 | |
| sau:SA2427 | arcB | Ornithine carbamoyltransferase catabolic | 0.33 |
| sau:SA2127 | rpi | Ribose-5-phosphate isomerase A | 0.32 |
| sau:SA1074 | fabG | 3-oxoacyl-[acyl-carrier-protein] reductase | 0.31 |
| sau:SA0160 | Heme-degrading monooxygenase isdI | 0.31 | |
| sau:SA2037 | rplN | 50S ribosomal protein L14 | 0.3 |
| sau:SA2089 | sarR | HTH-type transcriptional regulator sarR | 0.29 |
| sau:SA2022 | rplQ | 50S ribosomal protein L17 | 0.29 |
| sau:SA0473 | folB | Dihydroneopterin aldolase | 0.29 |
| sau:SA0108 | sarH1 | HTH-type transcriptional regulator sarS | 0.27 |
| sau:SA0573 | sarA | Transcriptional regulator sarA | 0.26 |
| Stationary Phase | |||
| ID | Gene | Annotation | emPAI |
| sau:SA0992 | trxA | Thioredoxin | 4.91 |
| sau:SA0295 | 30 kDa neutral phosphatase (Fragment) | 3.92 | |
| sau:SA0873 | UPF0477 protein SA0873 | 2.57 | |
| sau:SA1178 | UPF0154 protein SSP1415 | 2.14 | |
| sau:SA1305 | hu | DNA-binding protein HU | 1.85 |
| sau:SA1663 | UPF0342 protein SA1663 | 1.82 | |
| sau:SA2043 | rpsS | 30S ribosomal protein S19 | 1.6 |
| sau:SA1067 | rpmB | 50S ribosomal protein L28 | 1.55 |
| sau:SA0456 | spoVG | Putative septation protein spoVG | 1.47 |
| sau:SA1909 | atpF | ATP synthase subunit b | 1.46 |
| sau:SA1709 | ftn | Ferritin | 1.45 |
| sau:SA0160 | isdI | Heme-degrading monooxygenase isdI | 1.24 |
| sau:SA2062 | sarV | HTH-type transcriptional regulator sarV | 1.1 |
| sau:SA0760 | Glycine cleavage system H protein | 1.09 | |
| sau:SA0108 | sarH1 | HTH-type transcriptional regulator sarH1 | 1.04 |
| sau:SAS078 | rpmJ | 50S ribosomal protein L36 | 1.01 |
| sau:SA1904 | atpC | ATP synthase epsilon chain | 1.01 |
| sau:SA0032 | bleO | Bleomycin resistance protein | 0.99 |
| sau:SA0494 | nusG | Transcription antitermination protein nusG | 0.98 |
| sau:SA0478 | pdxT | Glutamine amidotransferase subunit pdxT | 0.97 |
| sau:SA2038 | rpsQ | 30S ribosomal protein S17 | 0.94 |
| sau:SA0245 | ispD | 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase 2 | 0.94 |
| sau:SA1901 | fabZ | (3R)-hydroxymyristoyl-[acyl-carrier-protein] dehydratase | 0.9 |
| sau:SA1256 | msrB | Peptide methionine sulfoxide reductase msrB | 0.89 |
| sau:SA0128 | sodM | Superoxide dismutase [Mn/Fe] 2 | 0.85 |
| sau:SA1019 | Uncharacterized N-acetyltransferase SA1019 | 0.85 | |
| sau:SA0437 | UPF0133 protein SAB0428 | 0.81 | |
| sau:SA1074 | fabG | 3-oxoacyl-[acyl-carrier-protein] reductase | 0.72 |
| sau:SA2431 | isaB | Immunodominant staphylococcal antigen B | 0.72 |
| sau:SA1041 | pyrR | Bifunctional protein pyrR | 0.7 |
| sau:SA2089 | sarR | HTH-type transcriptional regulator sarR | 0.65 |
| sau:SAP018 | arsC | Protein arsC | 0.59 |
| sau:SA2040 | rplP | 50S ribosomal protein L16 | 0.54 |
| sau:SA2266 | Uncharacterized oxidoreductase SAR2567 | 0.54 | |
| sau:SA1529 | UPF0173 metal-dependent hydrolase SA1529 | 0.52 | |
| sau:SA1146 | bsaA | Glutathione peroxidase homolog bsaA | 0.47 |
| sau:SA1076 | rnc | Ribonuclease 3 | 0.46 |
| sau:SA0774 | Probable ABC transporter ATP-binding protein | 0.46 | |
| sau:SA0941 | UPF0356 protein SA0941 | 0.46 | |
| sau:SA1461 | apt | Adenine phosphoribosyltransferase | 0.44 |
| sau:SA2392 | panB | 3-methyl-2-oxobutanoate hydroxymethyltransferase | 0.44 |
| sau:SA1206 | femA | Aminoacyltransferase femA | 0.44 |
| sau:SA0354 | rpsR | 30S ribosomal protein S18 | 0.43 |
| sau:SA0934 | ptsH | Phosphocarrier protein HPr | 0.42 |
| sau:SA1032 | sepF | Cell division protein sepF | 0.4 |
| sau:SA0470 | hsp33 | 33 kDa chaperonin | 0.4 |
| sau:SA1471 | rpmA | 50S ribosomal protein L27 | 0.39 |
| sau:SA1081 | rpsP | 30S ribosomal protein S16 | 0.39 |
| sau:SA0704 | UPF0230 protein | 0.39 | |
| sau:SA0826 | spsB | Signal peptidase IB | 0.39 |
| Log-Difference WT(+PQ) /DmrgA(+PQ) | Gene Name | N315 SA Number | Annotation/Similarity |
|---|---|---|---|
| 4.04 | mrgA(dps) | SA1941 | MrgA, Dps family protein |
| 2.07 | lrgB | SA0253 | antiholin-like protein LrgB |
| 1.86 | / | SA2133 | hypothetical protein |
| 1.65 | oppB | SA0853 | oligopeptide ABC transporter permease |
| 1.59 | arcB | SA2427 | ornithine carbamoyltransferase |
| 1.54 | sen | SA1643 | enterotoxin SeN (in pathogenicity island, SaPIn3) |
| 1.53 | / | SA2470 | histidinol dehydrogenase |
| 1.51 | / | SA2417 | nisin susceptibility-associated two-component sensor histidine kinase |
| 1.50 | / | SA2264 | hypothetical protein |
| 1.49 | hisZ | SA2472 | ATP phosphoribosyltransferase regulatory subunit/ His-tRNA synthase |
| 1.48 | / | SA2454 | acetyltransferase |
| 1.35 | / | SA2429 | ArgR family transcriptional regulator |
| 1.31 | / | SA0667 | 7-cyano-7-deazaguanine synthase |
| 1.30 | pyrF | SA1047 | orotidine 5′-phosphate decarboxylase |
| 1.30 | / | SA0846 | oligopeptide transport system permease OppC |
| 1.29 | / | SA1760 | holin-like protein (in phage phiN315) |
| 1.28 | / | SA1807 | mobile element associated protiein (in phage phiN315) |
| 1.26 | / | SA0804 | Na+/H+ antiporter family protein |
| 1.22 | lrgA | SA0252 | murein hydrolase regulator LrgA |
| 1.22 | yent1 | SA1645 | enterotoxin Yent1 (in pathogenicity island, SaPIn3) |
| 1.21 | / | SA2469 | histidinol-phosphate aminotransferase |
| 1.20 | / | SA0582 | monovalent cation/H+ antiporter subunit E |
| 1.15 | purC | SA0918 | phosphoribosylaminoimidazole-succinocarboxamide synthase |
| 1.11 | / | SA2189 | Ferrochelatase family / cobalamin biosynthesis CbiX/ transcriptional regulator NirR |
| 1.09 | ureE | SA2085 | urease accessory protein UreE |
| 1.09 | / | SA1768 | phage tail protein (in phage phiN315) |
| 1.03 | / | SA1636 | hypothetical protein |
| 1.02 | / | SA1675 | amino acid ABC transporter permease/substrate-binding protein |
| 1.02 | nrdD | SA2410 | anaerobic ribonucleoside triphosphate reductase |
| 1.02 | ssp | SA0744 | secretory extracellular matrix and plasma binding protein |
| 1.01 | / | SA0324 | mepB family protein |
| 1.00 | clfB | SA2423 | clumping factor B |
| Log-Difference WT(+PQ) /DmrgA(+PQ) | Gene Name | N315 SA Number | Annotation/Similarity |
|---|---|---|---|
| −3.03 | spa | SA0107 | immunoglobulin G binding protein A |
| −2.41 | / | SA0080 | membrane protein similar to sulfite exporter TauE/SafE family protein |
| −2.35 | / | SA0100 | Na/Pi cotransporter family protein |
| −2.31 | sirC | SA0109 | iron compound ABC transporter permease SirC |
| −1.92 | / | SA0090 | hypothetical protein |
| −1.90 | sarH1 | SA0108 | staphylococcal accessory regulator H1 |
| −1.87 | lacC | SA1995 | tagatose-6-phosphate kinase |
| −1.84 | / | SA0085 | hypothetical protein |
| −1.81 | / | SA0061 | (in Staphylococcus Cassette Chromosome, SCC) |
| −1.81 | / | SA0077 | serine/threonine protein kinase (in Staphylococcus Cassette Chromosome, SCC) |
| −1.78 | lctP | SA0106 | L-lactate permease |
| −1.73 | / | SA2092 | AraC family transcriptional regulator |
| −1.68 | / | SA0102 | myosin-cross-reactive MHC class-II like protein |
| −1.67 | lpl8 | SA0404 | lipoprotein encoded in pathogenicity island (in pathogenicity island, SaPIn2) |
| −1.63 | / | SA0124 | capsular polysaccharide biosynthesis glycosyltransferase TuaA |
| −1.61 | / | SA0120 | SbnI, siderophore biosynthesis protein |
| −1.54 | / | SA2230 | fmtA-like protein/ beta lactamase |
| −1.50 | / | SA0085 | tRNA-dihydrouridine synthase |
| −1.45 | / | SA0099 | transmembrane efflux pump protein |
| −1.43 | vraA | SA0533 | long chain fatty acid-CoA ligase vraA |
| −1.42 | / | SA2303 | ABC transporter permease protein |
| −1.42 | / | SA0097 | AraC/XylS family transcriptional regulator |
| −1.38 | sarY | SA2091 | staphylococcal accessory regulator Y |
| −1.21 | / | SA0105 | hypothetical protein |
| −1.20 | / | SA1826 | pathogenicity island protein (in pathogenicity island, SaPIn1) |
| −1.18 | / | SA2274 | hypothetical protein |
| −1.17 | / | SA2302 | ABC transporter ATP-binding protein |
| −1.12 | / | SA0087 | tfoX N-terminal domain protein |
| −1.08 | / | SA0078 | hypothetical protein |
| −1.07 | / | SA0037 | MaoC domain-containing protein (in Staphylococcus Cassette Chromosome, SCC) |
| −1.06 | / | SA0536 | hypothetical protein |
| −1.05 | / | SA2154 | hypothetical protein |
| −1.05 | / | SA0088 | hypothetical protein |
| −1.05 | lacA | SA1997 | galactose-6-phosphate isomerase subunit LacA |
| −1.04 | sarV | SA2062 | staphylococcal accessory regulator V |
| −1.04 | hisG | SA2471 | ATP phosphoribosyltransferase catalytic subunit |
| −1.03 | / | SAS028 | hypothetical protein |
| −1.00 | sodM | SA0128 | superoxide dismutase |
| −1.00 | fmhA | SA2199 | fmhA protein (FemAB like protein,) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morikawa, K.; Ushijima, Y.; Ohniwa, R.L.; Miyakoshi, M.; Takeyasu, K. What Happens in the Staphylococcal Nucleoid under Oxidative Stress? Microorganisms 2019, 7, 631. https://doi.org/10.3390/microorganisms7120631
Morikawa K, Ushijima Y, Ohniwa RL, Miyakoshi M, Takeyasu K. What Happens in the Staphylococcal Nucleoid under Oxidative Stress? Microorganisms. 2019; 7(12):631. https://doi.org/10.3390/microorganisms7120631
Chicago/Turabian StyleMorikawa, Kazuya, Yuri Ushijima, Ryosuke L. Ohniwa, Masatoshi Miyakoshi, and Kunio Takeyasu. 2019. "What Happens in the Staphylococcal Nucleoid under Oxidative Stress?" Microorganisms 7, no. 12: 631. https://doi.org/10.3390/microorganisms7120631
APA StyleMorikawa, K., Ushijima, Y., Ohniwa, R. L., Miyakoshi, M., & Takeyasu, K. (2019). What Happens in the Staphylococcal Nucleoid under Oxidative Stress? Microorganisms, 7(12), 631. https://doi.org/10.3390/microorganisms7120631