Aeromonas hydrophila, an Emerging Causative Agent of Freshwater-Farmed Whiteleg shrimp Litopenaeus vannamei

Abstract
1. Introduction
2. Materials and Methods
2.1. Shrimp and Reagents
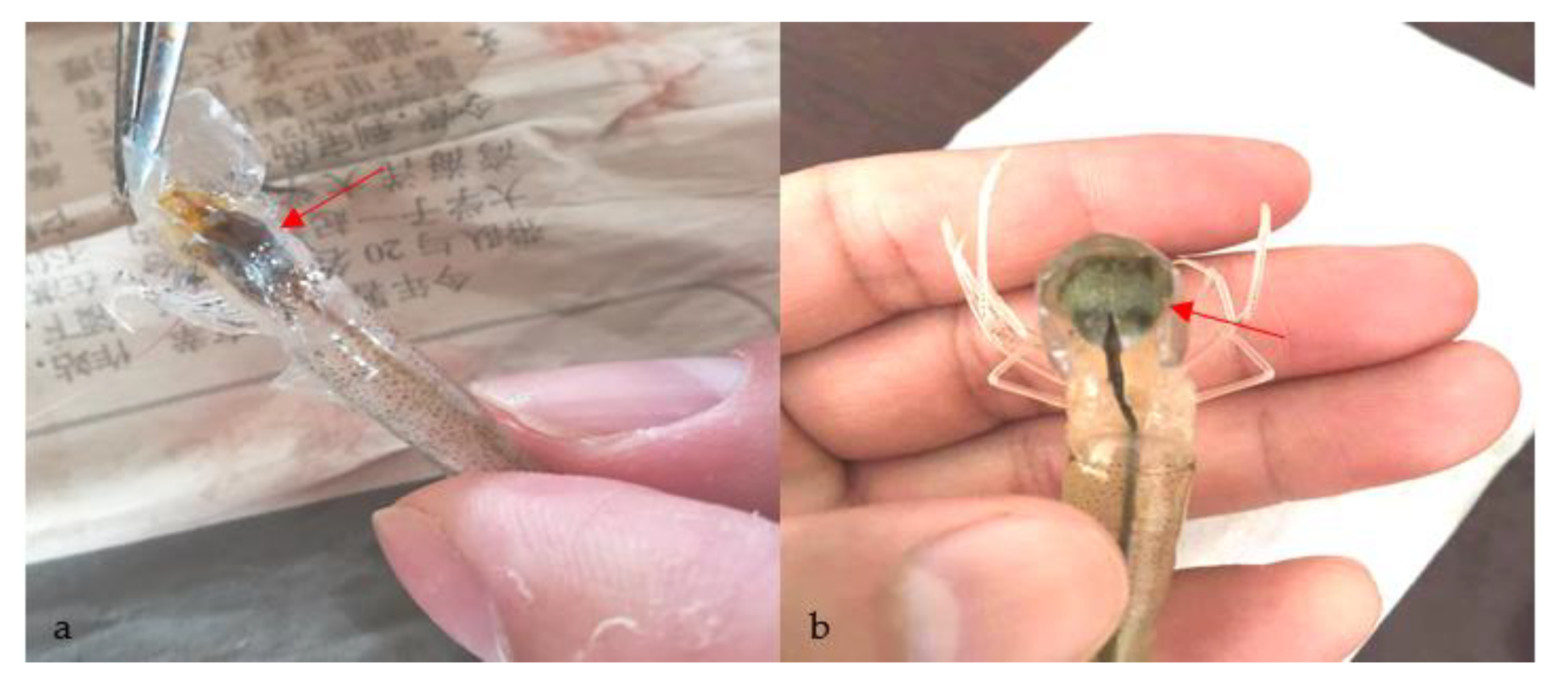
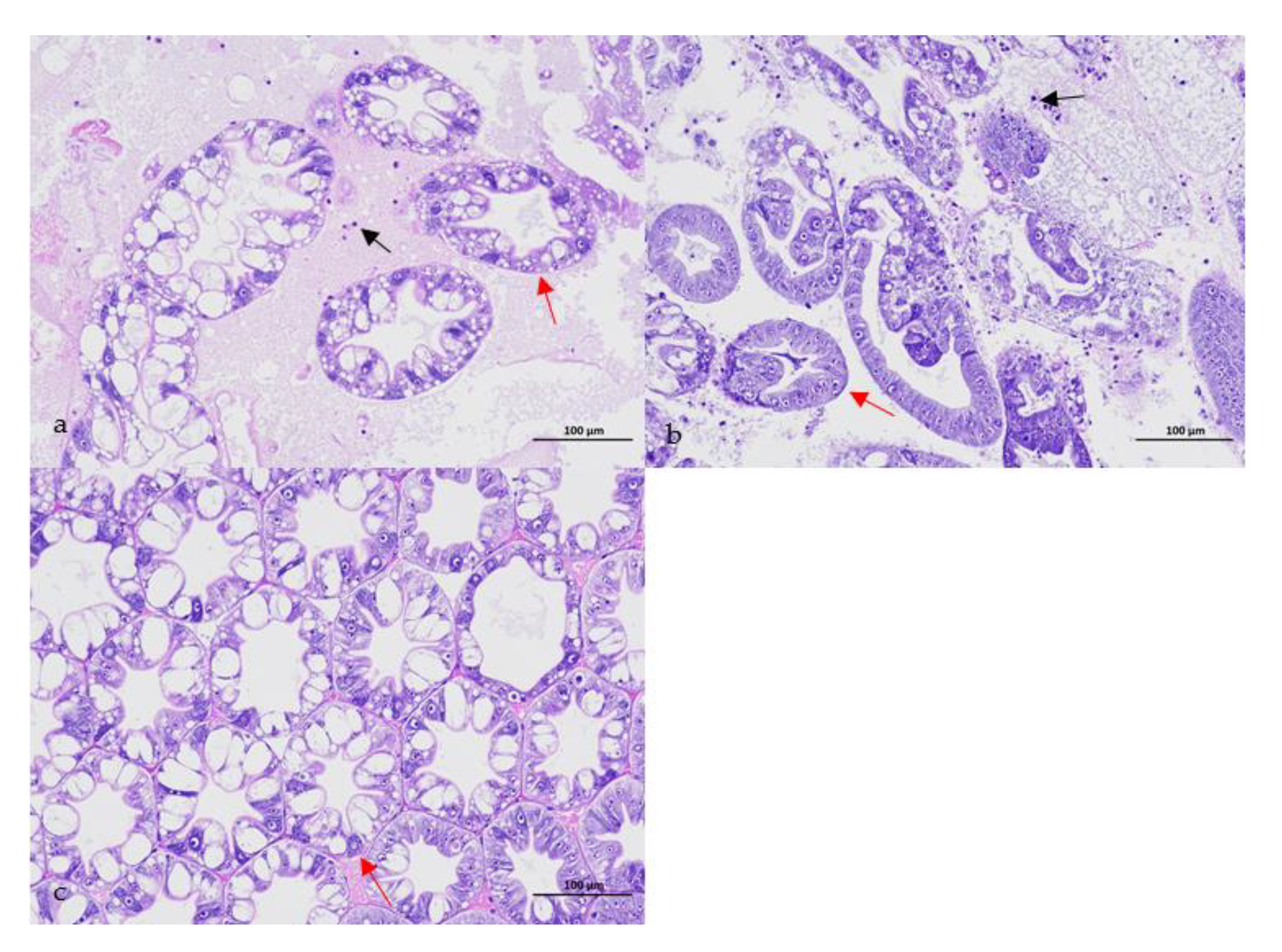
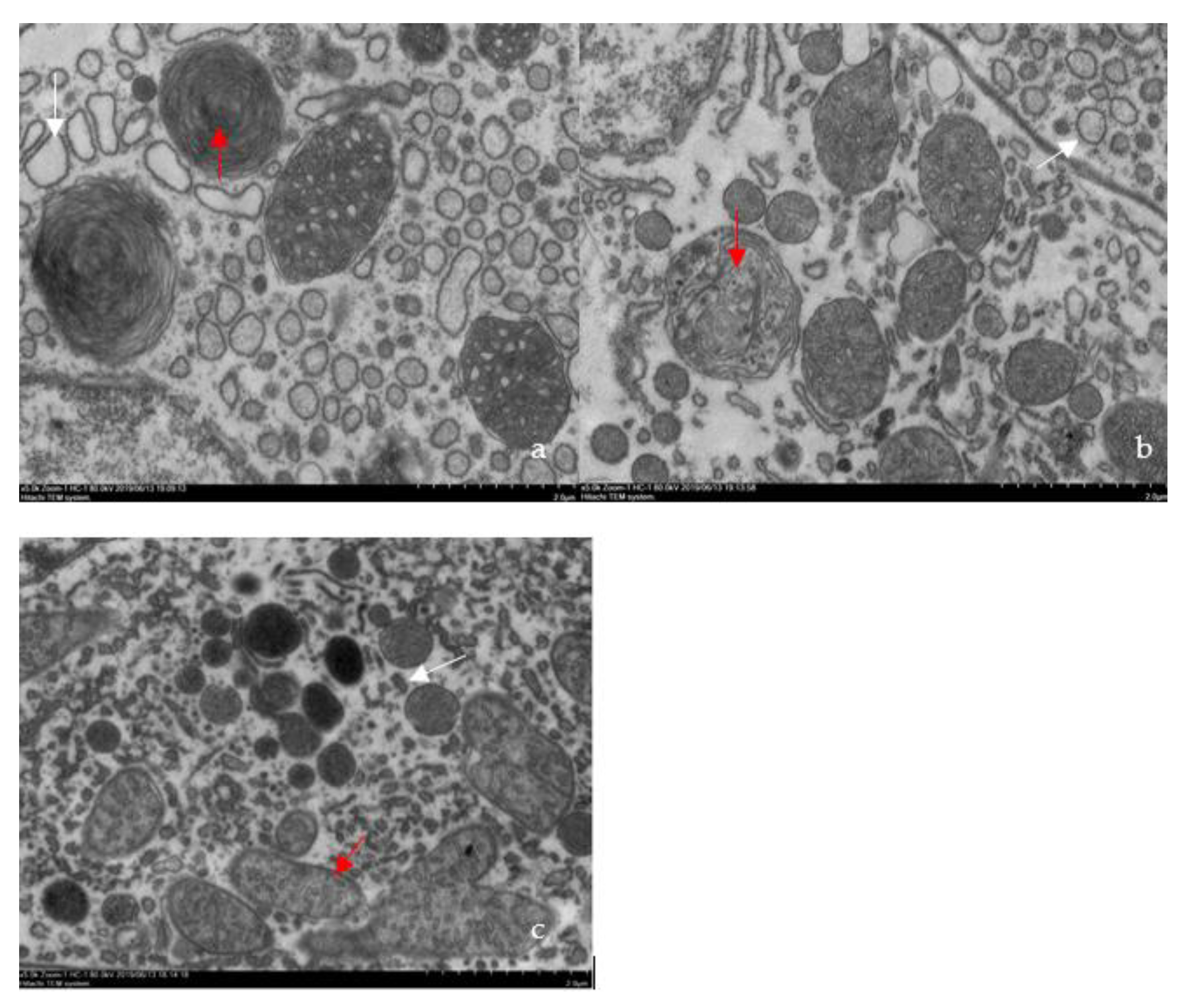
2.2. Confirmation of the Causative Pathogen
2.3. Identification of the Causative Pathogen
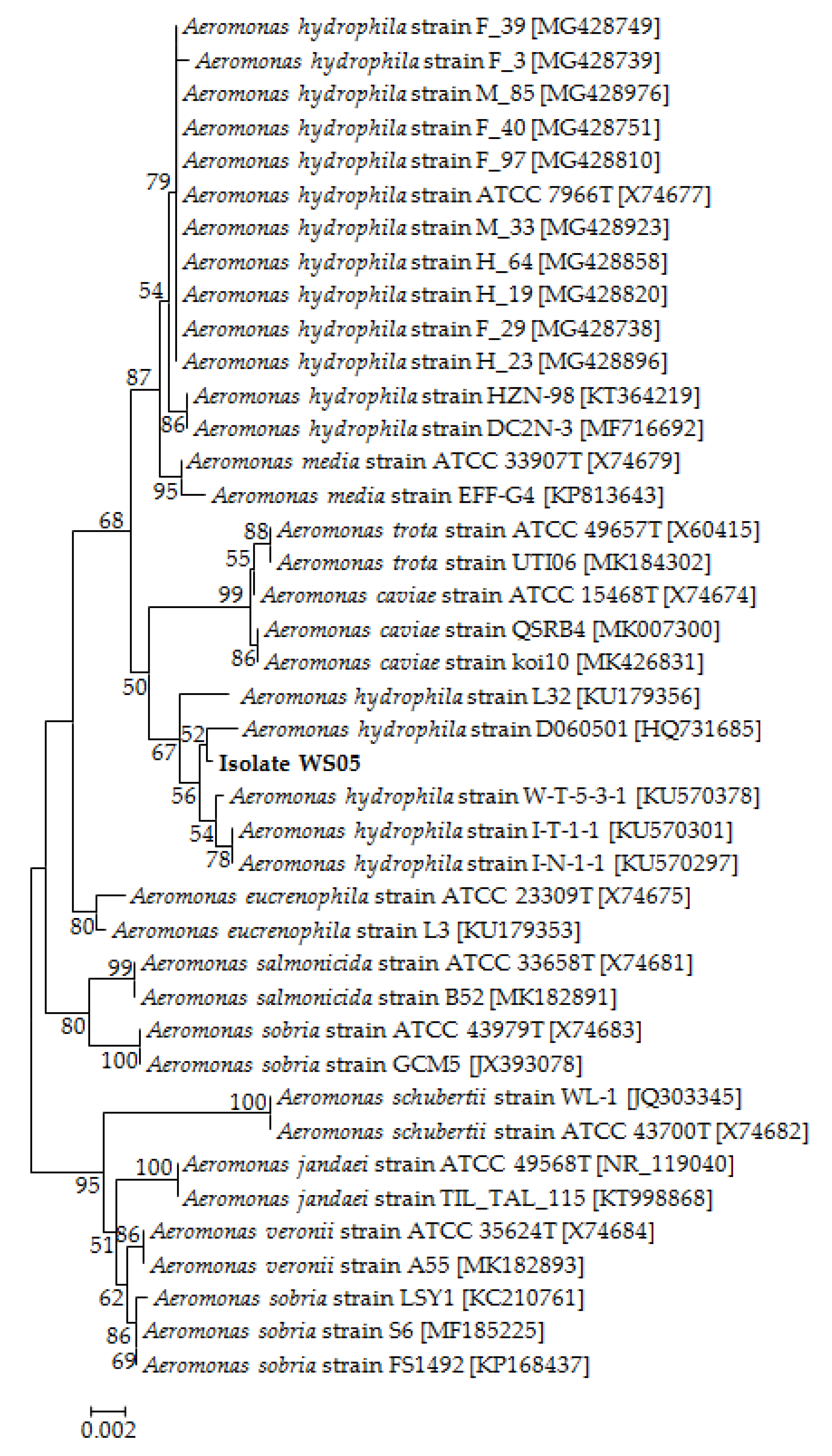
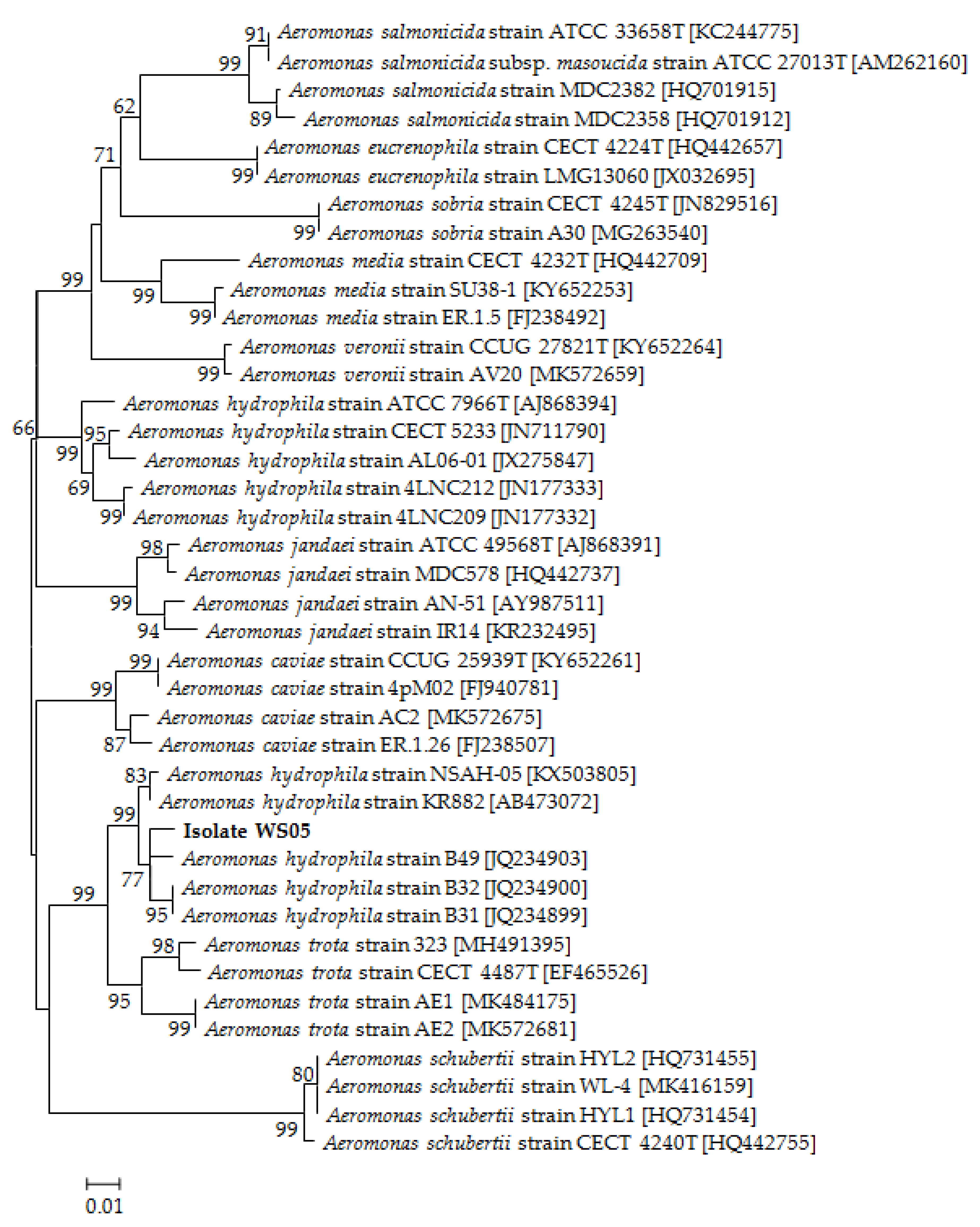
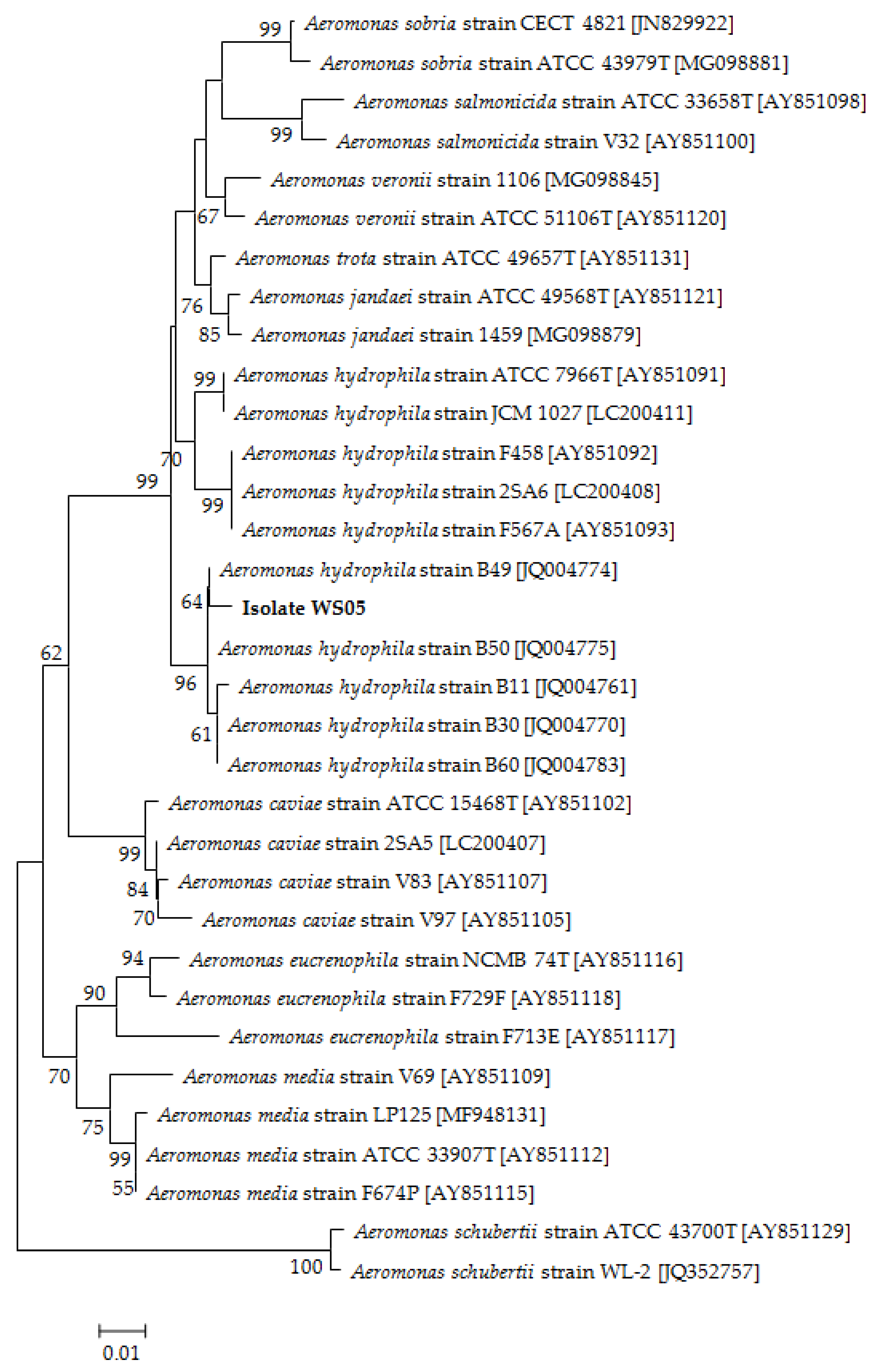
2.3.1. Molecular Identification
2.3.2. Phenotypic Identification
2.4. Bacterial Virulence Experiment
2.5. Antibiotics Susceptibility Assay
2.6. Synergistic Effect of Florfenicol–Herb Extracts Assay
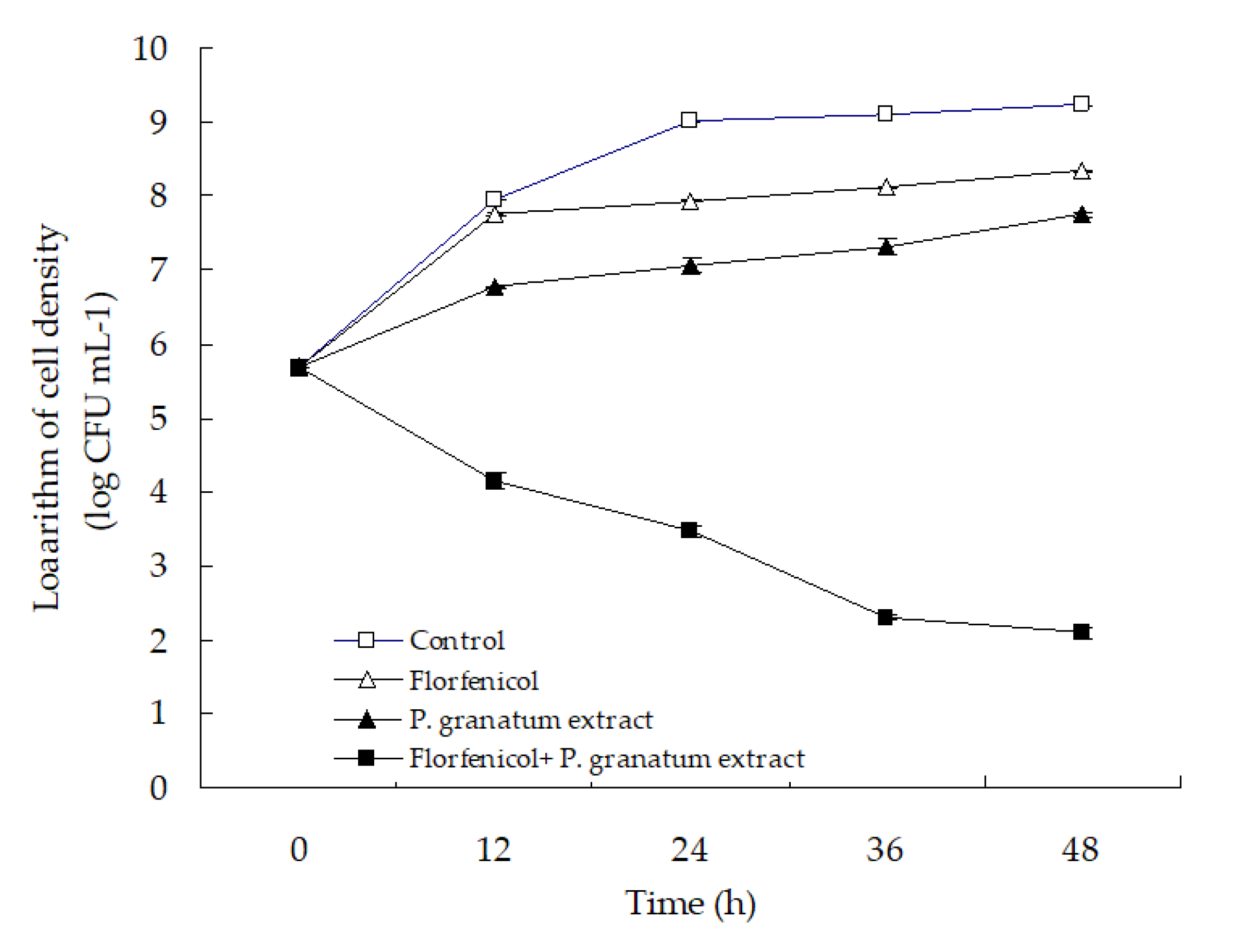
2.7. The in Vitro Activity of Florfenicol-Herb Extracts Assay
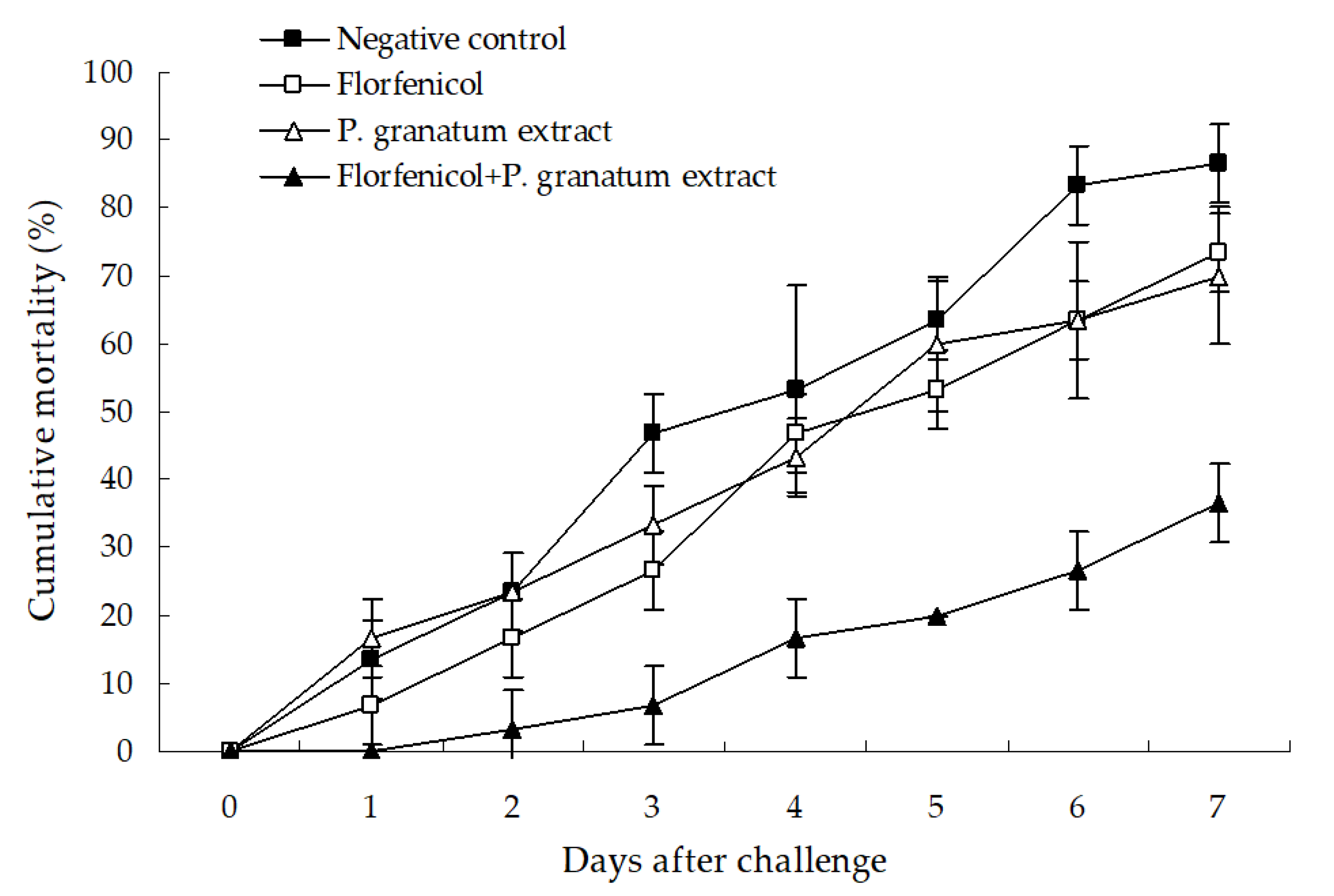
2.8. Protective Effect of Florfenicol-Herb Extracts Assay
2.9. Statistical Analysis
2.10. Ethics Statements
3. Results
3.1. Confirmation of the Causative Pathogen
3.2. Identification of the Pathogenic Isolate
3.3. Antibiotic Susceptibility of the Pathogenic Isolate
3.4. Synergistic Effect of Florfenicol–Herb Extracts
3.5. The in Vitro Activity of Florfenicol-P. granatum Extract
3.6. Protective Effect of Florfenicol-P. granatum Extract
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benzie, J.A.H. Use and exchange of genetic resources of penaeid shrimps for food and aquaculture. Rev. Aquacult. 2009, 1, 232–250. [Google Scholar] [CrossRef]
- Zhou, J. Development path and technological change of shrimp aquaculture in China. Chin. Agric. Sci. Bull. 2016, 32, 22–29. [Google Scholar]
- Ding, S.; Wang, F.; Dong, S.; Li, Y. Comparison of the respiratory metabolism of juvenile Litopenaeus vannamei cultured in seawater and freshwater. J. Ocean. Univ. China (Ocean. Coast. Sea Res.) 2014, 13, 331–337. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Rural Affairs of China. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2018; p. 24.
- Aguirre-Guzmán, G.; Ruíz, H.M.; Ascencio, R.F. A review of extracellular virulence product of Vibrio species important in diseases of cultivated shrimp. Aquac. Res. 2004, 35, 1395–1404. [Google Scholar] [CrossRef]
- Sudheesh, P.S.; Xu, H. Pathogenicity of Vibrio parahaemolyticus in tiger prawn Penaeus monodon Fabricius: Possible role of extracellular proteases. Aquaculture 2001, 196, 37–46. [Google Scholar] [CrossRef]
- Zhou, J.; Fang, W.; Yang, X.; Zhou, S.; Hu, L.; Li, X.; Qi, X.; Su, H.; Xie, L. A nonluminescent and highly virulent Vibrio harveyi strain is associated with “bacterial white tail disease” of Litopenaeus vannamei shrimp. PLoS ONE 2012, 7, e29961. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; An, J.; Zheng, W.; He, S. Vibrio cholerae pathogen from the freshwater-cultured whiteleg shrimp Penaeus vannamei and control with Bdellovibrio bacteriovorus. J. Invertebr. Pathol. 2015, 130, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; He, S.; Lu, L.; Yang, X.; Chen, B. Identification of a Proteus penneri isolate as the causal agent for the red body disease of the cultured white shrimp Penaeus vannamei and its control with Bdellovibrio bacteriovorus. Anton. Leeuwen. 2014, 105, 423–430. [Google Scholar] [CrossRef]
- Cao, H.; An, J.; He, S.; Lu, L.; Yang, X.; Zheng, W. Aeromonas schubertii: A potential pathogen for the freshwater cultured whiteleg shrimp Penaeus vannamei. Isr. J. Aquacult. Bamid. 2015, 67, 1–6. [Google Scholar]
- Cao, H.; Chen, S.; Lu, L.; An, J. Shewanella algae: An emerging pathogen of black spot disease in freshwater-cultured whiteleg shrimp (Penaeus vannamei). Isr. J. Aquacult. Bamid. 2018, 70, 1–7. [Google Scholar]
- Holmstrom, K.; Graslund, S.; Wahlstrom, A.; Poungshompoo, S.; Bengtsson, B.E.; Kautsky, N. Antibiotics use in shrimp farming and implications for environmental impacts and human health. Int. J. Food Sci. Tech. 2003, 38, 255–266. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Balasundaram, C. Modern trends in Aeromonas hydrophila disease management with fish. Rev. Fish. Sci. 2005, 13, 281–320. [Google Scholar] [CrossRef]
- Bhuvaneswari, R.; Balasundaram, C. Traditional Indian herbal extracts used in vitro against growth of the pathogenic bacteria—Aeromonas hydrophila. Isr. J. Aquacult. Bamid. 2006, 58, 89–96. [Google Scholar]
- Haniffa, M.A.; Kavitha, K. Antibacterial activity of medicinal herbs against the fish pathogen Aeromonas hydrophila. J. Agric. Tech. 2012, 8, 205–211. [Google Scholar]
- Al Laham, S.A.; Al Fadel, F.M. Antibacterial activity of various plants extracts against antibiotic-resistant Aeromonas hydrophila. Jundishapur. J. Microbiol. 2014, 7, e11370. [Google Scholar] [CrossRef]
- Türker, H.; Yildirim, A.B.; Karakaş, F.P.; Köylüoğlu, H. Antibacterial activities of extracts from some Turkish endemic plants on common fish pathogens. Turk. J. Biol. 2009, 33, 73–78. [Google Scholar]
- Nya, E.J.; Austin, B. Use of garlic, Allium sativum, to control Aeromonas hydrophila infection in rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish. Dis. 2009, 32, 963–970. [Google Scholar] [CrossRef]
- Pakravan, S.; Hajimoradloo, A.; Ghorbani, R. Effect of dietary willow herb, Epilobium hirsutum extract on growth performance, body composition, haematological parameters and Aeromonas hydrophila challenge on common carp, Cyprinus carpio. Aquac. Res. 2012, 43, 861–869. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M. The use of American ginseng (Panax quinquefolium) in practical diets for Nile tilapia (Oreochromis niloticus): Growth performance and challenge with Aeromonas hydrophila. J. Appl. Aquacult. 2012, 24, 366–376. [Google Scholar] [CrossRef]
- Wu, C.C.; Liu, C.H.; Chang, Y.P.; Hsieh, S.L. Effects of hot-water extract of Toona sinensis on immune response and resistance to Aeromonas hydrophila in Oreochromis mossambicus. Fish Shellfish. Immun. 2010, 29, 258–263. [Google Scholar] [CrossRef]
- Cheesman, M.J.; Ilanko, A.; Blonk, B.; Cock, I.E. Developing new antimicrobial therapies: Are synergistic combinations of plant extracts/compounds with conventional antibiotics the solution? Pharmacogn. Rev. 2017, 11, 57–72. [Google Scholar] [PubMed]
- Dong, J.; Ruan, J.; Xu, N.; Yang, Y.; Ai, X. In vitro synergistic effects of fisetin and norfloxacin against aquatic isolates of Serratia marcescens. FEMS Microbiol. Lett. 2016, 363, 1–5. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jedlicková, Z.; Mottl, O.; Sery, V. Antibacterial properties of the Vietnamese cajeput oil and ocimum oil in combination with antibacterial agents. J. Hyg. Epidemiol. Microbiol. Immunol. 1992, 36, 303–309. [Google Scholar] [PubMed]
- Jayaraman, P.; Sakharkar, M.K.; Lim, C.S.; Tang, T.H.; Sakharkar, K.R. Activity and interactions of antibiotic and phytochemical combinations against Pseudomonas aeruginosa in vitro. Int. J. Biol. Sci. 2010, 6, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghe, C.V.L.; Ahmed, S.B.N.; Kariyawasam, M.G.I.U. The isolation and identification of Vibrio species in marine shrimps of Sri Lanka. J. Food Agric. 2008, 1, 36–44. [Google Scholar] [CrossRef]
- Moss, S.M.; Pruder, G.D.; Leber, K.M.; Wyban, J.A. The relative enhancement of Penaeus vannamei growth by selected fractions of shrimp pond water. Aquaculture 1992, 101, 229–239. [Google Scholar] [CrossRef]
- Moss, S.M.; Divakaran, S.; Kim, B.G. Stimulating effects of pond water on digestive enzyme activity in the Pacific white shrimp, Litopenaeus vannamei (Boone). Aquac. Res. 2001, 32, 125–131. [Google Scholar] [CrossRef]
- Network of Aquaculture Centers in Asia-Pacific. Shrimp Health Management Extension Manual; MPEDA House: Cochin, India, 2003; p. 23. [Google Scholar]
- Biswas, G.; Korenaga, H.; Nagamine, R.; Kono, T.; Shimokawa, H.; Itami, T.; Sakai, M. Immune stimulant effects of a nucleotide-rich baker’s yeast extract in the Kuruma shrimp, Marsupenaeus japonicus. Aquaculture 2012, 366, 40–45. [Google Scholar] [CrossRef]
- Luo, G.; Tan, H.; Zhu, X. The effects of several water-treatment units in a recirculating aquaculture system. J. Dalian Fish. Univ. 2008, 23, 68–72. [Google Scholar]
- Ekanem, A.P.; Eyo, V.O.; Sampson, A.F. Parasites of landed fish from Great Kwa River, Calabar, Cross River State, Nigeria. Int. J. Fish. Aquacult. 2011, 3, 225–230. [Google Scholar]
- Pan, X.; Shen, J.; Li, J.; He, B.; Yin, W.; Hao, G.; Xu, Y.; Yao, J. Identification and biological characteristics of the pathogen causing Macrobrachium nipponense soft-shell syndrome. Microbiol. China 2009, 36, 1571–1576. [Google Scholar]
- Cao, H.; Yang, Y.; Chen, S.; Ai, X. Providencia alcalifaciens: A causal agent of red leg disease in freshwater-cultured whiteleg shrimp Penaeus vannamei. Isr. J. Aquacult. Bamid. 2018, 70, 1–7. [Google Scholar]
- Orozova, P.; Sirakov, I.; Petkov, I.; Crumlish, M.; Austin, B. Recovery of Aeromonas hydrophila associated with bacteraemia in captive snakes. FEMS Microbiol. Lett. 2012, 334, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Petti, C.A.; Polage, C.R.; Schreckenberger, P. The role of 16S rRNA gene sequencing in identification of microorganisms misidentified by conventional methods. J. Clin. Microbiol. 2005, 43, 6123–6125. [Google Scholar] [CrossRef]
- Cao, H.; He, S.; Lu, L.; Hou, L. Characterization and phylogenetic analysis of the bitrichous pathogenic Aeromonas hydrophila isolated from diseased Siberian sturgeon. Isr. J. Aquacult. Bamid. 2010, 62, 181–189. [Google Scholar]
- Yáňez, M.A.; Catalán, V.; Apráiz, D.; Figueras, M.J.; Martínez-Murcia, A.J. Phylogenetic analysis of members of the genus Aeromonas based on gyrB gene sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 875–883. [Google Scholar] [CrossRef]
- Küpfer, M.; Kuhnert, P.; Korczak, B.M.; Peduzzi, R.; Demarta, A. Genetic relationships of Aeromonas strains inferred from 16S rRNA, gyrB and rpoB gene sequences. Int. J. Syst. Evol. Microbiol. 2006, 56, 2743–2751. [Google Scholar] [CrossRef]
- Abeyta, C.; Kaysner, C.A.; Wekell, M.M.; Sullivan, J.J.; Stelma, G.N. Recovery of Aeromonas hydrophila from oysters implicated in an outbreak of foodborne illness. J. Food Protect. 2019, 49, 643–646. [Google Scholar] [CrossRef]
- Long, S.; Han, S.; Niu, Z.; Liang, J.; Hu, D.; Huang, J.; Li, H.; Liu, Q.; Su, J. Isolation and identification of pathogenic Aeromonas in Clarias fuscus and analysis of the correlation between its pathogenicity and virulence genorypes. J. Fish. China 2016, 40, 308–317. [Google Scholar]
- Ye, W.; Guo, C.; Cao, H.; Yang, X. Isolation, identification, pathogenicity and in vitro antibacterial drugs of pathogenic Aeromonas hydrophila from Micropterus salmoides with hemorrhagic disease. Freshw. Fish. 2018, 5, 54–60. [Google Scholar]
- Teng, T.; Liang, L.; Chen, K.; Xi, B.; Xie, J.; Xu, P. Isolation, identification and phenotypic and molecular characterization of pathogenic Vibrio vulnificus isolated from Litopenaeus vannamei. PLoS ONE 2017, 12, e0186135. [Google Scholar] [CrossRef] [PubMed]
- Ogbuagu, D.H.; Iwuchukwu, E.I. Evaluation of the toxicity of three hair shampoos on the catfish (Clarias gariepinus) fingerlings. Appl. Ecol. Environ. Sci. 2014, 2, 86–89. [Google Scholar]
- Jones, R.N.; Ballow, C.H.; Biedenbach, D.J. Multi-laboratory assessment of the linezolid spectrum of activity using the Kirby-Bauer disk diffusion method: Report of the Zyvox (R) antimicrobial potency study (ZAPS) in the United States. Diagn. Microbiol. Infect. Dis. 2001, 40, 59–66. [Google Scholar] [CrossRef]
- Chen, Z.; Li, X.; Wu, X.; Wang, W.; Wang, W.D. Synergistic activity of econazole-nitrate and chelerythrine against clinical isolates of Candida albicans. Iran. J. Pharm. Res. 2014, 13, 567–573. [Google Scholar] [PubMed]
- Liu, Q.; Li, J.; Wang, Q.; Dai, F. Pharmacodynamic Test Technical Specification for Aquaculture Antimicrobial Agent—Part1: Antibiotics Susceptibility Test of Macro-Broth Dilution Method; China Standards Press: Beijing, China, 2011; pp. 1–6. [Google Scholar]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Prophylactic effect of Andrographis paniculata extracts against Streptococcus agalactiae infection in Nile tilapia (Oreochromis niloticus). J. Biosci. Bioeng. 2009, 107, 579–582. [Google Scholar] [CrossRef]
- Liu, I.X.; Durham, D.G.; Richards, R.M.E. Baicalin synergy with β-lactam antibiotics against methicillin–resistant Staphylococcus aureus and other β-lactam-resistant strains of S. aureus. J. Pharm. Pharmacol. 2000, 52, 361–366. [Google Scholar] [CrossRef]
- Haroun, M.; Al-Kayali, R.S. Synergistic effect of Thymbra spicata L. extracts with antibiotics against multidrug-resistant Staphylococcus aureus and Klebsiella pneumoniae strains. Iran. J. Basic Med. Sci. 2016, 19, 1193–1200. [Google Scholar]
- Liu, H.; Zhang, W.; Wu, Y.; Sun, L.; Wang, Y.; Liu, Y.; Zhang, X.; Hong, P.; Ji, H. Synergistic antimicrobial effect and mechanism of lipopeptides and tea polyphenols against Vibrio parahaemolyticus. Food Sci. 2017, 38, 14–19. [Google Scholar]
- Gong, X.; Le, G.; Li, Y. Antibacterial spectrum of antibacterial peptides from Musca dommestica larvae and synergic interaction between the peptides and antibiotics. Acta Microbiol. Sin. 2005, 45, 516–520. [Google Scholar]
- Zou, W.; Yang, C.; Jiang, I.; Wu, S.; Yi, Q.; Wu, J. Guideline of fishery drug application (NY5071-2002). Ocean. Fish. 2007, 12, 39–41. [Google Scholar]
- Pillai, S.K.; Moellering, R.C.; Eliopoulos, G.M. Antimicrobial combinations. In Antibiotics in Laboratory Medicine, 5th ed.; Lorian, V., Ed.; Williams & Wilkins: Baltimore, MD, USA, 2005; pp. 365–440. [Google Scholar]
- Dierckens, K.R.; Vandenberghe, J.; Beladjal, L.; Huys, G.; Mertens, J.; Swings, J. Aeromonas hydrophila causes ‘black disease’ in fairy shrimps (Anostraca; Crustacea). J. Fish. Dis. 1998, 21, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Jiravanichpaisal, P.; Roos, S.; Edsman, L.; Liu, H.; Soderhall, K. A highly virulent pathogen, Aeromonas hydrophila, from the freshwater crayfish Pacifastacus leniusculus. J. Invertebr. Pathol. 2009, 101, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.F.; Rashid, M.M.; Sayed, M.A. Experimental infection of indigenous climbing perch Anabas testudineus with Aeromonas hydrophila bacteria. Prog. Agric. 2011, 22, 105–114. [Google Scholar] [CrossRef]
- Zheng, W.; Cao, H.; Yang, X. Grass carp (Ctenopharynngodon idellus) infected with multiple strains of Aeromonas hydrophila. Afr. J. Microbiol. Res. 2012, 21, 4512–4520. [Google Scholar]
- Rashid, M.M.; Hossain, M.S.; Ali, M.F. Isolation and identification of Aeromonas hydrophila from silver carp and its culture environment from Mymensingh region. J. Bangladesh Agric. Univ. 2013, 2, 373–376. [Google Scholar] [CrossRef]
- Zhu, C.; Zhou, X.; Zhang, Q. Pathogenic bacterium identification and histopathology of septicemia of juvenile southern catfish, Silurus meridionalis Chen. J. Fish. Sci. China 2011, 2, 360–370. [Google Scholar] [CrossRef]
- Ye, Y.W.; Fan, T.F.; Li, H.; Lu, J.F.; Jiang, H.; Hu, W.; Jiang, Q.H. Characterization of Aeromonas hydrophila from hemorrhagic diseased freshwater fishes in Anhui Province, China. Int. Food Res. J. 2013, 3, 1449–1452. [Google Scholar]
- Gai, C.; Ye, W.; Lu, L.; Li, Y.; Yang, X.; Cao, H. Aeromonas hydrophila: A causative agent for tail rot disease in freshwater cultured Murray cod Maccullochella peelii. Isr. J. Aquacult. Bamid. 2016, 68, 1–8. [Google Scholar]
- Altarriba, M.; Merino, S.; Gavin, R.; Canals, R.; Rabaan, A.; Shaw, J.G.; Tomas, J.M. A polar flagella operon (flg) of Aeromonas hydrophila contains genes required for lateral flagella expression. Microb. Pathol. 2003, 34, 249–259. [Google Scholar] [CrossRef]
- Cascón, A.; Yugueros, J.; Temprano, A.; Sánchez, M.; Hernanz, C.; Luengo, J.M.; Naharro, G. A major secreted elastase is essential for pathogenicity of Aeromonas hydrophila. Infect. Immun. 2000, 68, 3233–3241. [Google Scholar] [CrossRef]
- Gonzalez-Serrano, C.J.; Santos, J.A.; García-López, M.L.; Otero, A. Virulence markers in Aeromonas hydrophila and Aeromonas veronii biovar sobria isolates from freshwater fish and from a diarrhea case. J. Appl. Microbiol. 2002, 93, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Erova, T.E.; Pillai, L.; Fadl, A.A.; Sha, J.; Wang, S.; Galindo, C.L.; Chopra, A.K. DNA adenine methyltransferase influences the virulence of Aeromonas hydrophila. Infect. Immun. 2006, 74, 410–424. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dong, J.; Liu, Y.; Xu, N.; Yang, Q.; Yang, Y.; Ai, X. Expression, purification and characterization of aerolysin from Aeromonas hydrophila. Chin. Fish. Qual. Stand. 2019, 9, 27–33. [Google Scholar]
- Jiang, Q.; Ye, Y.; Hu, W.; Jiang, H.; Lu, J. Research progresses of virulence factors and control technologies in Aeromonas hydrophila. Mod. Agric. Sci. Technol. 2012, 6, 324–327. [Google Scholar]
- Haldar, S.; Chatterjee, S.; Asakura, M.; Vijayakumaran, M.; Yamasaki, S. Isolation of Vibrio parahaemolyticus and Vibrio cholerae (Non-O1 and O139) from moribund shrimp (Penaeus monodon) and experimental challenge study against post larvae and juveniles. Ann. Microbiol. 2007, 57, 55–60. [Google Scholar] [CrossRef]
- Liu, C.H.; Chen, J.C. Effect of ammonia on the immune response of white shrimp Litopenaeus vannamei and its susceptibility to Vibrio alginolyticus. Fish. Shellfish. Immunol. 2004, 16, 321–334. [Google Scholar] [CrossRef]
- Hsu, S.W.; Chen, J.C. The immune response of white shrimp Penaeus vannamei and its susceptibility to Vibrio alginolyticus under sulfide stress. Aquaculture 2007, 271, 61–69. [Google Scholar] [CrossRef]
- Vivekanandhan, G.; Savithamani, K.; Hatha, A.A.M.; Lakshmanaperumalsamy, P. Antibiotic resistance of Aeromonas hydrophila isolated from marketed fish and prawn of South India. Int. J. Food Microbiol. 2002, 76, 165–168. [Google Scholar] [CrossRef]
- Hatha, M.; Vivekanandhan, A.A.; Joice, G.J.; Christol. Antibiotic resistance pattern of motile aeromonads from farm raised fresh water fish. Int. J. Food Microbiol. 2005, 98, 131–134. [Google Scholar] [CrossRef]
- Belém-Costa, A.; Cyrino, J.E.P. Antibiotic resistance of Aeromonas hydrophila isolated from Piaractus mesopotamicus (Holmberg, 1887) and Oreochromis niloticus (Linnaeus, 1758). Sci. Agric. 2006, 63, 281–284. [Google Scholar] [CrossRef]
- Defoirdt, T.; Sorgeloos, P.; Bossier, P. Alternatives to antibiotics for the control of bacterial disease in aquaculture. Curr. Opin. Microbiol. 2011, 14, 251–258. [Google Scholar] [CrossRef]
- Lee, C.K.; Kim, H.; Moon, K.H.; Shin, K.H. Screening and isolation of antibiotic resistance inhibitors from herb materials-resistance inhibition of volatile components of Korean aromatic herbs. Arch. Pharm. Res. 1998, 21, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Endo, E.H.; Cortez, D.A.G.; Ueda-Nakamura, T.; Nakamura, C.V.; Filho, B.P.D. Potent antifungal activity of extracts and pure compound isolated from pomegranate peels and synergism with fluconazole against Candida albicans. Res. Microbiol. 2010, 161, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Prashanth, D.; Asha, M.K.; Amit, A. Antibacterial activity of Punica granatum. Fitoterapia 2001, 72, 171–173. [Google Scholar] [CrossRef]
- Li, G.; Guo, W.; Gao, X.; Wang, Y.; Sun, S.; Xia, X. Synergistic antimicrobial effect and mechanism of punicalagin and chlorogenic acid against Staphyloccocus aureus. Sci. Tech. Food Ind. 2018, 39, 17–21. [Google Scholar]
- Lu, X.; Zhou, L.; Xie, K.; Jing, Y.; Yun, B.; Wang, Y.; Xie, M. Antibacterial activity and mechanism of gallic acid on Staphylococcus aureus. Edib. Fung. China 2012, 31, 54–56. [Google Scholar]
- Choi, M.J.; Lee, E.; Lee, S.; Reza, M.A.; Le, J.; Gebru, E. The in vitro antibacterial activity of florfenicol in combination with amoxicillin or cefuroxime against pathogenic bacteria of animal origin. Pak. Vet. J. 2011, 31, 141–144. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests | Reaction | |
|---|---|---|
| Isolate WS05 | A. hydrophilaa | |
| Arginine dihydrolase | R + | R + |
| Cytochrome oxidase | R + | R + |
| β-Galactosidase | R + | R + |
| Gelatinase | R + | R + |
| Lysine decarboxylase | R + | R + |
| Ornithine decarboxylase | R − | R − |
| Tryptophan deaminase | R − | R − |
| Urease | R − | R − |
| Citrate utilization | R + | R + |
| Acetoin production | R + | R + |
| Indole production | R + | R + |
| H2S production | R − | R − |
| Arabinose fermentation | R + | R + |
| Amygdalin fermentation | R − | R − |
| Glucose fermentation | R + | R + |
| Inositol fermentation | R − | R − |
| Mannitol fermentation | R + | R + |
| Melibiose fermentation | R − | R − |
| Rhamnose fermentation | R − | R − |
| Sucrose fermentation | R + | R + |
| Sorbitol fermentation | R − | R − |
| Drug Alone | MICs | Drug Combination | MICs in Combination | FICI | |
|---|---|---|---|---|---|
| FFC | Herb Extract | ||||
| FFC | 0.50 ± 0 h | FFC + AA | 0.13 ± 0 d | 15.63 ± 0 e | 0.75 ± 0 fg |
| AA | 31.25 ± 0 e | FFC + AC | 0.03 ± 0 e | 62.5 ± 0 b | 0.56 ± 0 gh |
| AC | 125 ± 0 b | FFC + RAT | 0.5 ± 0 b | 15.63 ± 0 e | 1.06 ± 0 def |
| RAT | 250 ± 0 a | FFC + CS | 0.03 ± 0 e | 62.5 ± 0 b | 1.06 ± 0 def |
| CS | 62.5 ± 0 c | FFC + FT | 1.00 ± 0 a | 5.86 ± 2.76 ef | 2.19 ± 0.09 a |
| FT | 31.25 ± 0 e | FFC + GC | 0.21 ± 0.06 c | 2.60 ± 0.92 f | 0.67 ± 0.12 gh |
| GC | 10.41 ± 3.68 g | FFC + PM | 0.03 ± 0 e | 7.81 ± 0 ef | 0.89 ± 0.24 efg |
| PM | 10.41 ± 3.68 g | FFC + PA | 0.5 ± 0 b | 52.08 ± 14.73 c | 1.42 ± 0.12 cd |
| PA | 125 ± 0 b | FFC + PG | 0.03 ± 0 e | 7.81 ± 0 ef | 0.31 ± 0 h |
| PG | 31.25 ± 0 e | FFC + PC | 1.00 ± 0 a | 15.63 ± 0 e | 2.06 ± 0 ab |
| PC | 250 ± 0 a | FFC + RRR | 0.21 ± 0.06 c | 31.25 ± 0 d | 1.25 ± 0.2 de |
| RRR | 41.67 ± 14.73 d | FFC + RA | 0.13 ± 0 d | 26.04 ± 7.36 d | 0.67 ± 0.12 gh |
| RA | 62.5 ± 0 c | FFC + RS | 0.03 ± 0 e | 31.25 ± 0 d | 1.73 ± 0.47 bc |
| RS | 20.84 ± 7.36 f | FFC + RSF | 0.03 ± 0 e | 62.5 ± 0 b | 0.56 ± 0 gh |
| RSF | 125 ± 0 b | FFC + RSA | 0.21 ± 0.06 c | 11.72 ± 5.52 ef | 1.08 ± 0.24 def |
| RSA | 20.84 ± 7.36 f | FFC + RB | 0.03 ± 0 e | 500 ± 0 a | 2.06 ± 0 ab |
| RB | 250 ± 0 a | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Gai, C.; Ye, G.; An, J.; Liu, K.; Xu, L.; Cao, H. Aeromonas hydrophila, an Emerging Causative Agent of Freshwater-Farmed Whiteleg shrimp Litopenaeus vannamei. Microorganisms 2019, 7, 450. https://doi.org/10.3390/microorganisms7100450
Zhou H, Gai C, Ye G, An J, Liu K, Xu L, Cao H. Aeromonas hydrophila, an Emerging Causative Agent of Freshwater-Farmed Whiteleg shrimp Litopenaeus vannamei. Microorganisms. 2019; 7(10):450. https://doi.org/10.3390/microorganisms7100450
Chicago/Turabian StyleZhou, Huihua, Chunlei Gai, Guifang Ye, Jian An, Kai Liu, La Xu, and Haipeng Cao. 2019. "Aeromonas hydrophila, an Emerging Causative Agent of Freshwater-Farmed Whiteleg shrimp Litopenaeus vannamei" Microorganisms 7, no. 10: 450. https://doi.org/10.3390/microorganisms7100450
APA StyleZhou, H., Gai, C., Ye, G., An, J., Liu, K., Xu, L., & Cao, H. (2019). Aeromonas hydrophila, an Emerging Causative Agent of Freshwater-Farmed Whiteleg shrimp Litopenaeus vannamei. Microorganisms, 7(10), 450. https://doi.org/10.3390/microorganisms7100450