Abstract
Aflatoxigenic molds can grow and produce aflatoxins on dry-fermented meat and cheese. The small, basic, cysteine-rich antifungal protein PgAFP displays a time-limited inhibitory ability against unwanted molds by increasing reactive oxygen species (ROS), which can lead to increased aflatoxin production. However, calcium abolishes the inhibitory effect of PgAFP on certain Aspergillus spp. To maximize the antifungal effect, this protein may be combined with protective cultures. Yeasts and lactic acid bacteria may counteract the impact of calcium on PgAFP fungal inhibition. The objective of this work was to study the effect of PgAFP and different combined treatments with Debaryomyces hansenii and/or Pediococcus acidilactici against growth of and aflatoxin production by an aflatoxigenic strain of Aspergillus parasiticus in both culture media and dry-fermented foods with low or high calcium levels. Aflatoxins production was increased by PgAFP but dramatically reduced by P. acidilactici in low calcium culture medium, whereas in the Ca-enriched culture medium, all treatments tested led to low aflatoxins levels. To study whether PgAFP and the protective microorganisms interfere with ROS and aflatoxin production, the relative expression of genes foxA, which is involved in peroxisomal β-oxidation, and aflP, which is required for aflatoxin biosynthesis, were evaluated. The aflatoxin overproduction induced by PgAFP seems not to be linked to peroxisomal β-oxidation. The combination of PgAFP and D. hansenii provided a successful inhibitory effect on A. parasiticus growth as well as on aflatoxin production on sliced dry-fermented sausage and cheese ripened up to 15 days, whereas P. acidilactici did not further enhance the protective effect of the two former agents. Therefore, the combined treatment of PgAFP and D. hansenii seems to provide a promising protective mean against aflatoxin-producing A. parasiticus on dry-fermented foods.
1. Introduction
Various microorganisms, including bacteria and fungi, decisively contribute to the specific characteristics of dry-fermented foods. However, the environmental conditions during the ripening of dry-fermented foods favor colonization of their surface by toxigenic molds that are able to produce mycotoxins on such foods. Aflatoxins have long been a major concern in cheese [1,2,3,4,5] and recent studies have highlighted their presence in dry-cured meats [6,7,8,9]. Aflatoxins, produced mainly by Aspergillus flavus and Aspergillus parasiticus, are classified as group 1 carcinogenic to humans by the International Agency for Research on Cancer. Therefore, it is necessary to design strategies to prevent aflatoxigenic molds growth on dry-fermented foods. The use of protective cultures could provide widely accepted preventive means to control mycotoxin production in ripened foods.
Some molds produce proteins that inhibit other molds and some yeasts, whereas the activity against prokaryotes is quite limited [10,11,12,13,14]. These antifungal proteins may serve to combat unwanted molds in foods. PgAFP from Penicillium chrysogenum [15] is one of these antifungal proteins with fungistatic effect, requiring less than 4.9 μg/mL for the highly sensitive molds, including A. flavus on dry-fermented sausage [16]. However, higher concentrations of up to 312.7 μg/mL are required for low sensitive molds, including A. parasiticus. In addition, the antifungal capability of this kind of proteins is dramatically reduced in presence of divalent cations, such as Ca2+ [17,18,19], rendering this protein ineffective against A. flavus on cheese [20]. PgAFP not only can inhibit growth, but also hamper aflatoxins biosynthesis. A. flavus treated with PgAFP for 24 h showed lower relative abundance of key enzymes in aflatoxin biosynthesis pathway, including O-methyltranferase, versicolorin B (VERB) synthase, and VER1 dehydrogenase [16]. However, the effect on aflatoxin production in low sensitive molds has not been tested yet. On the other hand, it is known that reactive oxygen species (ROS) contribute to aflatoxin biosynthesis [21,22], which makes it necessary to study the influence of subinhibitory concentrations of PgAFP on aflatoxin production.
The multifactorial mechanism of action of PgAFP increases ROS levels on sensitive fungal strains [16]. Some aspergilli are able to deal with the excess of toxic ROS by synthetizing aflatoxins, where peroxisomal β-oxidation of fatty acids plays a key role [23]. Expression of gene foxA has been used as a marker for β-oxidation in A. flavus [24] and A. parasiticus [25]. Similarly, the expression of gene aflP, coding for O-methyltransferase A, has been used to monitor aflatoxin biosynthesis [26,27]. Therefore, studying the expression of these two genes could provide information on whether PgAFP treatment increases aflatoxin production by oxidative stress.
On the other hand, the limited time extension of PgAFP antifungal activity makes it necessary to develop strategies that extend mold growth inhibition. A plausible alternative can be based on the combined use of PgAFP and microbial strains endowed with antifungal activity. Among the microorganisms usually present on dry-fermented products, yeasts thrive on dry-cured hams for the whole ripening time, being the dominant species Debaryomyces hansenii [28,29]. Some of these strains have shown antifungal capability against toxigenic Penicillia on dry-cured meat products [30,31,32]. Also, lactic acid bacteria have been proved as potent mold inhibitors [33]. Pediococcus acidilactici produces antimicrobial compounds [34,35], some of them being active against A. parasiticus [36].
The aim of this work was to investigate the antifungal capability of PgAFP alone and with D. hansenii and/or Pediococcus acidilactici against A. parasiticus in culture medium as well as on dry-fermented sausage and on cheese. To understand how these agents affect β-oxidation and aflatoxin production, their effect on foxA and aflP genes expression was also studied.
2. Material and Methods
2.1. Microbial Strains
Two mold strains obtained from the Spanish Type Culture Collection (CECT, Valencia, Spain) were used in this study: the PgAFP sensitive aflatoxin-producing A. parasiticus CECT 2682 [10] and the PgAFP-producer P. chrysogenum CECT 20922. In addition, the yeast D. hansenii FHSCC 253H from the microbial collection of Food Hygiene and Safety of the University of Extremadura (Cáceres, Spain) and P. acidilactici fargo 35 supplied by Laboratorios Amerex (Colmenar Viejo, Spain) were tested as potential antifungal organisms.
2.2. PgAFP Production and Purification
P. chrysogenum CECT 20922 was grown in potato dextrose broth (PDB; Scharlab, Barcelona, Spain) pH 4.5 incubated statically for 21 days at 25 °C. The mycelium was removed and the medium was filtered through a 0.22 μm-pore-size nylon membrane (MSI, Westboro, USA) to obtain a cell free medium. PgAFP was isolated from 450 mL of the cell-free medium through fast protein liquid chromatography (FPLC) with a cationic exchange column HiTrap SP HP (Amersham Biosciences, Uppsala, Sweden), further purified with a HiLoad 26/60 Superdex 75 gel filtration column (Amersham Biosciences) as previously described [37]. The isolated and concentrated protein was sterilized through filtration (0.22 µm, Thermo Fisher Scientific, Waltham, MA, USA) and its concentration was assessed by the Lowry method [38].
2.3. PgAFP and Microbial Inocula Preparation
A. parasiticus was grown on potato dextrose agar (PDA; Scharlab, Barcelona, Spain) for 15 days at 25 °C. Then, conidia were harvested by washing the surface of incubated plates with sterile phosphate-saline buffer (PBS). D. hansenii was grown in yeast extract sucrose (20 g/L yeast extract, 125 g/L sucrose; YES) broth for 72 h under continuous shaking at 200 rpm and 25 °C. YES medium containing D. hansenii was centrifuged to concentrate cells and the pellet was washed twice with sterile PBS. P. acidilactici was grown in de Man-Rogosa-Sharpe (MRS; Scharlab, Barcelona, Spain) broth for 24 h at 30 °C, and to concentrate the cells the same procedure previously described for yeasts was followed. Spore or yeast suspensions were counted in a Thoma counting chamber.
2.4. Mold Growth Inhibition and Mycotoxin Extraction in Culture Media
The effect of PgAFP, D. hansenii, and P. acidilactici on A. parasiticus was evaluated both separately and in different combinations. Samples were inoculated with A. parasiticus to reach 105 conidia/mL or cm2. To provide a high level of potential antifungal agents, 106 cells/mL or cm2 D. hansenii, 106 cells/mL or cm2 P. acidilactici, or 10 µg/mL or cm2 PgAFP were added as required. An untreated batch inoculated only with A. parasiticus was used in each experiment as a control. The remaining batches were prepared using A. parasiticus and different combination of the aforementioned biopreservative agents, as described below.
The effect of PgAFP, D. hansenii, and P. acidilactici on A. parasiticus was evaluated in 5 mL YES broth and 0.1 M CaCl2-enriched YES (Ca-YES) broth. For this, the seven following treatments were tested in YES broth and Ca-YES broth: PgAFP alone (Pg), D. hansenii alone (Dh), P. acidilactici alone (Pa), PgAFP combined with D. hansenii (Pg + Dh), PgAFP combined with P. acidilactici (Pg + Pa), D. hansenii combined with P. acidilactici (Dh + Pa), and PgAFP combined with D. hansenii and P. acidilactici (Pg + Dh + Pa). Samples were incubated statically for 15 days at 25 °C to stimulate aflatoxin production [26]. All the assays were carried out in triplicate.
For mycotoxin extraction after incubation, test tubes were added with 5 mL of chloroform and shaken at 100 rpm for 1 h at room temperature in darkness. Then, the chloroform was separated and evaporated to dryness under a gentle stream of N2. To evaluate mold growth, the mycelium was recovered from the aqueous residue by filtering through a Miracloth (Calbiochem, Darmstadt, Germany), dried to constant weight at 100 °C, and weighed.
2.5. Mold Growth Inhibition on Dry-Ripened Foods
A commercial raw dry-fermented sausage (pH 5.8, 0.96 aw) shortly after stuffing in natural beef casing was cut into 7–8 mm thick slices. Similarly, commercial Gouda cheese was cut into 5 mm thick slices. Both sausage and cheese slices were dipped into ethanol to eliminate outer contamination. The ethanol was left to evaporate in a laminar flow cabinet Bio II (Telstar, Tarrasa, Spain) prior to placing the slices in ethanol-sterilized receptacles containing a saturated KCl solution to keep relative humidity constant at 84% after vapor-liquid equilibrium at 25 °C. A. parasiticus was inoculated on both sides of each sausage and cheese slice at 105 conidia/cm2. No antifungal treatment was applied to control samples. The two treated batches were prepared by adding either 10 µg/cm2 PgAFP and 106 cells/cm2 D. hansenii (Pg + Dh) or 10 µg/cm2 PgAFP and 106 cells/cm2 each of D. hansenii and P. acidilactici (Pg + Dh + Pa) onto the sausage and cheese slices. All samples received the same total volume of liquid by adding the required amount of sterile phosphate-saline buffer (PBS). After spreading uniformly the inoculum over the cheese surface with a sterile bent glass rod, the liquid excess was left to dry in the laminar flow cabinet. Every batch was prepared in triplicate.
To stimulate aflatoxin production, all batches were incubated at 25 °C [26]. Samples were taken after 5 and 15 days of incubation. Water activity (aw) values were determined with a LabMaster-aw (Novasina, Lachen, Switzerland). A. parasiticus and D. hansenii counts were assessed on PDA incubated for 72 h at 25 °C, according to the characteristic colony morphology. Lactic acid bacteria counts were evaluated on MRS agar incubated for 72 h at 30 °C.
2.6. Mycotoxin Extraction
Mycotoxins produced on sausage or cheese slices were extracted as previously described [9]. Briefly, each sample slice was macerated under shaking in a dark flask with 60 mL acetonitrile-water (9:1, v/v) containing 0.1% formic acid, together with 50 mL hexane. The acetonitrile-water phase was recovered and filtered through sodium sulphate anhydrous. Then, the filtrate was mixed with additional 50 mL of hexane and shaken. The acetonitrile-water phase was filtered again and evaporated in a rotary evaporator at 40–45 °C. The residue was resuspended in 1 mL chloroform, filtered through a 0.45 µm nylon membrane (MSI, Westboro, MA, USA) and evaporated to dryness under a gentle stream of nitrogen.
2.7. Mycotoxin Quantification
The dried chloroform extracts from culture media, sausage, and cheese slices were resuspended in 100 μL of high-performance liquid chromatography (HPLC)-grade acetonitrile (Sharlab, Barcelona, Spain) and analyzed by ultra high-performance liquid chromatography–mass spectrometry (UHPLC-MS) in a Dionex UltiMate 3000 HPLC system (Thermo Fisher Scientific) coupled to an ion trap mass spectrometer (MS) model Amazon SL (Bruker Daltonics, Bremen, Germany). A C18 reverse-phase column of 10 cm length, 2.1 mm inner diameter and 1.8 μm particle size (Agilent Technologies, Santa Clara, CA, USA) was used as stationary phase. The mobile phases were (A) 0.1% formic acid-10 mM ammonium formate and (B) acetonitrile. The separation was performed at 200 µL/min flow rate and the following gradient: 0 min, 2% B; 0–0.1 min, 2–40% B; 0.1–4 min, 40–60% B; 4–7 min, 60–80% B; 7–8.5 min, 80% B; 8.5–8.51 min, 80–98% B; 8.51–12 min, 98% B; 12–12.01 min, 98–2% B; and 12.01–15 min, 2% B. Retention time for aflatoxins B1 and G1 (AFB1 and AFG1) were 6.4 ± 0.5 and 6.3±0.5 min respectively. Precursor ions 313 and 329, and quantitation ions 285 and 311 for AFB1 and AFG1, respectively, were used.
These parameters were compared to those obtained from commercial mycotoxins (Sigma-Aldrich, Madrid, Spain). The calibration curves for AFB1 and AFG1 (1–500 ng) by uHPLC-MS revealed a linear relationship (r2 ≥ 0.99) between the detector response and the amount of AFB1 and AFG1 standards. The minimum detectable value or limit of detection (LOD) was estimated from the calibration curve, according to the equation: LOD = 3 (sB2 + si2 + (i/m)2 sm)1/2/m [39], with “m” being the slope of the calibration curve, “I” being the intercept term, and “sB”, “si”, and “sm” being the standard errors of the blank, the intercept term, and the slope of the calibration curve, respectively. Assuming a normal distribution of the estimated quantities, α (error of the first type) = β (error of the second type) = 0.05, the quantification limit (LOQ) was 3.04 LOD [40]. The LOD obtained in this study were 4 ppb and 1.5 ppb, and the LOQ were 12 ppb and 4.5 ppb, respectively, for AFB1 and AFG1.
2.8. Gene Expression Studies
To assess the influence of PgAFP alone or in combination with D. hansenii and P. acidilactici on A. parasiticus gene expression, four batches were prepared in YES broth and Ca-YES broth. A. parasiticus was co-cultivated in tubes with Pg, Dh, Pa, and Pg + Dh + Pa. An untreated batch was inoculated solely with A. parasiticus (control). The tubes were incubated at 25 °C for 5 days. After this, mycelium was harvested and washed twice with sterile PBS, frozen in liquid nitrogen, ground with a mortar and pestle, and stored at −80 °C until RNA extraction.
2.8.1. RNA Isolation and Complementary DNA Synthesis
For RNA extraction, the frozen ground mycelium was resuspended in 750 µL RLT buffer (RNeasy® Plant Mini kit, Qiagen, Hilden, Germany) containing 15 µL β-mercaptoethanol. Samples were processed with the RNeasy® Plant Mini kit according to manufacturer’s instructions. RNA quality and quantity were spectrophotometrically determined using in a NanoDrop 2000c spectrophotometer (Thermo Scientific, Waltham, MA, USA). To remove genomic DNA contamination, samples were treated with DNase I, RNase-free (Fermentas, St. Leon-Rot, Germany) following manufacturer’s instructions. Finally, complementary DNA (cDNA) was synthesized using 500 ng of total RNA according to the PrimeScript™ RT Reagent kit protocol (Takara, Otsu, Japan).
2.8.2. Relative Quantification of Gene Expression by Real Time-Polymerase Chain Reaction
The expression of genes aflP (formerly omt-1) for aflatoxin biosynthesis [27], foxA for β-oxidation [25], and β-tubulin as housekeeping marker [41] were assessed to evaluate the effect of the different treatments on A. parasiticus. Primers F-Omt-1 and R-Omt-1, previously designed by [42], were used to evaluate aflP expression. To evaluate foxA expression, primers F-foxA and R-foxA were designed from the gene AFLA_041590 (GenBank accession no. XM_002377580). Similarly, to study β-tubulin expression, primers F-β-tub and R-β-tub were designed from A. parasiticus β-tubulin sequence (GenBank accession no. FR775333.1). Primers were designed on exon in the target using the Primer-Blast tool from the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/tools/primer-blast/). Nucleotide sequences of primers used in the real time-quantitative polymerase chain reaction (RT-qPCR) assays are shown in Table 1.

Table 1.
Nucleotide sequence of primers used for the quantitative polymerase chain reaction (qPCR) assay.
The ViiATM 7 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) was used to carry out the RT-qPCR assays. To optimize the primer concentration, different concentrations from 800 to 200 nM were tested. SYBR Green methodology was applied. The optimized SYBR Green protocols were carried out in a final volume of 25 µL containing 5 µL of template cDNA, 12.5 µL of 2x SYBR® Premix Ex Taq™ (Takara, Otsu, Japan), 0.5 µL of 50x ROX™ Reference Dye (Takara) and 400 nM of each primer. The thermal cycling conditions were the following: a single step of 10 min at 95 °C, 40 cycles of 95 °C for 15 s, and 60 °C for 1 min. After the final PCR cycle, melting curves analysis of the PCR products were carried out by heating from 60 to 95 °C and continuous measurement of the fluorescence to verify the PCR product. The aflP gene amplification was evaluated following the concentrations and conditions previously described [42]. Ct determinations were automatically performed by the instrument using defaults parameters. For every primer pair, a standard curve was generated to check the amplification efficiency from ten-fold serial dilution 100 to 0.01 ng/µL of DNA from A. parasiticus. Every qPCR assay described above was carried out by triplicate.
To quantify the relative expression of the genes, foxA and β-tubulin genes, the 2−ΔΔCT method was used [43]. The endogenous control was set as the expression of β-tubulin gene and the untreated samples (control batch) was used as calibrator sample. To check the amplification efficiency for every primer pair, a standard curve was generated from 10-fold serial dilutions (100 to 0.01 ng/µL) of DNA from A. parasiticus. The differences in amplification efficiencies between the housekeeping and the target gene were 3 and 5% for aflP and foxA genes, respectively, as required to apply the relative quantification method [44].
2.9. Statistical Analysis
Statistical analyses were performed with the IBM SPSS v.22. Data from mycelia weight, microbial counts, mycotoxin concentration and gene expression were tested for normality (Kolmogorov-Smirnov with Lilliefors correction) and homoscedasticity (Levene’s test). Given that these data were non-normally distributed, untreated and treated samples were compared in pairs using the nonparametric Mann-Whitney U test (p ≤ 0.05). To study the relationship between expression of aflP and foxA genes, a Spearman correlation was applied (p ≤ 0.05).
3. Results
3.1. A. parasiticus Growth Inhibition and Mycotoxin Production in Culture Media
To study the antifungal capability of the three agents, both separately and in different combinations, seven treatments (Pg, Dh, Pa, Pg + Dh, Pg + Pa, Dh+Pa, and Pg + Dh + Pa) were tested against A. parasiticus for 15 days in YES and Ca-YES broth. A. parasiticus growth was substantial in YES broth, regardless of the treatment applied (Figure 1). As a result of the high standard deviation, there was no statistically significant difference (p > 0.05) in A. parasiticus growth between any treatment and the untreated control. On the other hand, A. parasiticus grew poorly in Ca-YES broth and was further inhibited by most treatments, but not by PgAFP alone (Figure 1).
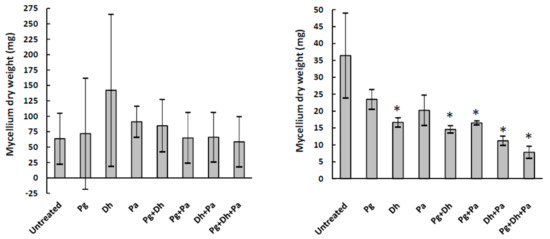
Figure 1.
Mycelium dry weight of A. parasiticus cultured in yeast extract sucrose broth (left) and calcium-enriched yeast sucrose broth (right) for 15 days with different combinations of biopreservative agents. Pg: PgAFP, Dh: Debaryomyces hansenii, Pa: Pediococcus acidilactici. * Means are significantly different from untreated control (p ≤ 0.05).
Both AFB1 and AFG1 were produced in the untreated batch by A. parasiticus in both culture media (Figure 2). Aflatoxin production showed differences (p ≤ 0.05) depending on the treatment applied. PgAFP treatment increased AFB1 and AFG1 production in YES broth, but led to lower aflatoxin concentrations in Ca-YES. All treatments containing P. acidilactici strongly inhibited aflatoxins production. Batches inoculated with D. hansenii also showed lower AFB1 production in both culture media. In YES broth, Pg + Dh induced a higher AFG1 quantity whereas no statistically significant increase was obtained for AFB1 compared to the untreated control. In Ca-YES broth, A. parasiticus produced both AFB1 and AFG1 above the LOD only in the untreated control, and AFG1 in the batch treated solely with D. hansenii. Therefore, PgAFP neither alone nor combined with any of the remaining protective agents tested increased aflatoxin production in Ca-YES broth.
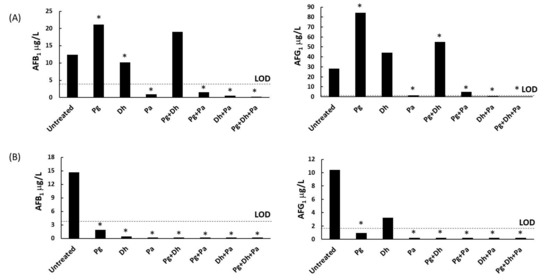
Figure 2.
Effect of different combinations of biopreservative agents in aflatoxin B1 (left) and G1 (right) production by Aspergillus parasiticus in yeast extract sucrose broth (A) and calcium-enriched yeast extract sucrose broth (B) after 15 days. Pg: PgAFP; Dh: Debaryomyces hansenii; Pa: Pediococcus acidilactici. LOD: Limit of detection. * Means are significantly different from untreated batch (p ≤ 0.05).
3.2. A parasiticus Growth Inhibition and Mycotoxin Production on Sliced Sausage
The effect of PgAFP-combined treatments on mold development was evaluated on slices of raw sausage kept under controlled environmental conditions simulating normal ripening. Aw values reached 0.93 and 0.86 at 5 and 15 days of incubation, respectively (Table 2). The microbial load in the pre-sterilized sliced sausage just before inoculation was 2.3 log cfu/cm2 for molds, 1 log cfu/cm2 for lactic acid bacteria, and lower than 1.7 log cfu/cm2 for yeasts. After treatment application, yeast counts were always over 5 log cfu/cm2 in D. hansenii-inoculated batches, but lower than 2 log cfu/cm2 in the untreated batch. Similarly, MRS counts were over 6.8 log cfu/cm2 in P. acidilactici-inoculated samples, but lower than 4.5 log cfu/cm2 in the remaining ones. Mold counts from untreated batches were over 6.5 and 7.4 log cfu/cm2 at five and 15 days, respectively (Table 2). Pg + Dh or Pg + Dh + Pa treatments dramatically decreased A. parasiticus counts by about 3 log units.
Table 2.
Aflatoxins concentrations and microbial counts on slices of dry-fermented sausage at five and 15 days of incubation. Data are given as mean ± SD.
On the other hand, mycotoxin production in the sausages was somehow parallel to fungal growth. A. parasiticus produced high AFB1 and AFG1 levels in untreated sausages, both at five and 15 days of incubation (Table 2). However, aflatoxin production in Pg + Dh and Pg + Dh + Pa batches was lower (p ≤ 0.05) than in the untreated control at both sampling times.
3.3. A. parasiticus Growth Inhibition and Mycotoxin Production on Cheese Slices
The microbial load on cheese slices just before inoculation was about 3 log cfu/cm2 for lactic acid bacteria (LAB) and below 2 log cfu/cm2 for both molds and yeasts (Table 3). LAB increased just after inoculation to 6–6.7 log cfu/cm2 in Pg + Dh + Pa batch, which was the only group of cheese samples inoculated with P. acidilactici. At five and 15 incubation days, LAB counts remained around 3.5 log cfu/cm2 for batches not inoculated with P. acidilactici (untreated and Pg + Dh), but over 6 log cfu/cm2 for Pg + Dh + Pa batches. Similarly, the yeast load was always below 2 log cfu/cm2 in the untreated control, but over 5 log cfu/cm2 in Pg + Dh and Pg + Dh + Pa batches. The aw value of cheese slices after inoculation was 0.96, and went down to 0.86 at 15 days of incubation (Table 3). Fungal loads at five days were around 2 log units higher (p≤0.05) in untreated than in Pg + Dh and Pg + Dh + Pa batches (Table 3). At 15 days, fungal counts in untreated samples were lower than 5 log cfu/cm2. Counts around 2–3 log cfu units lower (p ≤ 0.05) were also obtained with Pg + Dh and Pg + Dh + Pa treatments after incubation for 15 days.
Table 3.
Water activity (aw), microbial counts and aflatoxins concentration in cheese slices inoculated with A. parasiticus at five and 15 days of incubation. Data are given as mean ± SD.
The only mycotoxin obtained from cheese at the conditions tested was AFG1 produced by A. parasiticus in the untreated batch at 5 and 15 days (Table 3). The high AFG1 levels detected in these samples at day 15 were well over the 10–20 μg/kg limit established for some foods in different countries.
3.4. Gene Expression
The effect of PgAFP, D. hansenii, and P. acidilactici on the expression of genes aflP for aflatoxin biosynthesis, foxA for β-oxidation, and β-tubulin as housekeeping were assessed in A. parasiticus cultures grown in YES and Ca-YES broth for five days, when PgAFP activity on A. parasiticus is expected to be high. The expression of aflP gene was repressed in A. parasiticus by the sole action of either PgAFP or P. acidilactici as well as the Pg + Dh + Pa combined treatment in YES broth (Figure 3). When A. parasiticus was grown in Ca-YES broth, aflP was also repressed (p ≤ 0.05) in Pg batch, but overexpressed (p ≤ 0.05) in Pa batch (Figure 3).
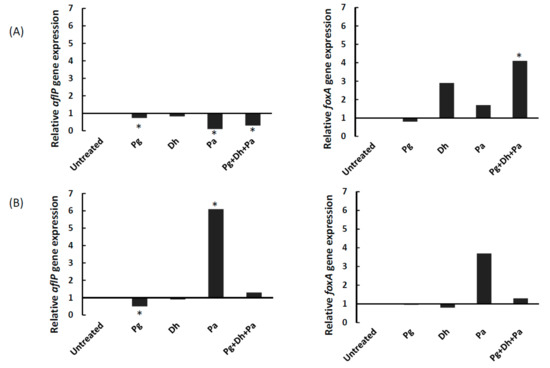
Figure 3.
Effect of different combinations of biopreservative agents on relative gene expression of aflP (left) and foxA (right) genes in yeast extract sucrose broth (A) or calcium-enriched yeast extract sucrose broth (B) after five days. Pg: PgAFP; Dh: Debaryomyces hansenii; Pa: Pediococcus acidilactici. *Means are significantly different from untreated batch (p ≤ 0.05).
The expression of foxA only differed (p ≤ 0.05) from its untreated control in the Pg + Dh + Pa samples from YES broth, leading to a 4-fold overexpression (Figure 3). No treatment led to statistically significant changes for foxA expression in Ca-YES broth, despite the high expression rate obtained from Pa samples (Figure 3). In addition, foxA gene expression values did not correlate with aflP expression values (p > 0.05).
4. Discussion
As expected, PgAFP did not inhibit A. parasiticus growth in YES or Ca-YES media (Figure 1). The level of 10 µg/mL PgAFP used was too low to reach a significant effect, as it was shown previously [10]. However, to test the effect of PgAFP on aflatoxins production, a concentration below that causing moderate inhibition had to be tested. Additionally, neither P. acidilactici nor D. hansenii effectively inhibited A. parasiticus growth in the YES medium. On the other hand, most treatments tested inhibited (p ≤ 0.05) A. parasiticus growth in Ca-YES broth (Figure 1). The highest inhibition reached by Pg + Dh + Pa seemed to be a consequence of the combined effect of the different mechanisms of action. PgAFP activity is based on permeability induction, loss of membrane integrity, and apoptosis induction as a consequence of ROS [16], whereas the inhibitory effect of D. hansenii is attributed to volatile compounds and competition for nutrients and space [31]. In addition, P. acidilactici produces bacteriocins and organic acids that inhibit mold growth [35,45].
Pg + Dh and Pg + Dh + Pa treatments showed a remarkable inhibition on A. parasiticus on the sliced sausages (Table 2). Mold counts in non-treated sausages were higher than those in Pg + Dh and Pg + Dh + Pa batches by Day 5. Given that 3.5 log cfu/cm2 has been suggested as the limit to keep minimal risk of AFB1 accumulation in dry-cured hams [9], the two PgAFP-combined treatments applied could be useful to control aflatoxigenic molds in dry-cured meat products. At 15 days incubation time, which covers most of the ripening stage of dry-fermented sausages, mold counts in treated samples were still around 0.1% of the mold load on the untreated samples.
The lack of inhibitory effect of the tested antifungal agents in the culture medium as compared to the efficient inhibition in dry-fermented sausages can be attributed to the differences in substrate availability. The mode of action proposed for yeasts to control molds in culture media is competition for nitrogen compounds, sugars, and vitamins [46]. In addition, some volatile compounds derived from branched amino acids by D. hansenii are responsible for mold inhibition [31,47]. YES broth contains 125 g/L sucrose and 20 g/L yeast extract, providing readily available sources of carbon and nitrogen compounds, as well as vitamins, especially from the B complex. Conversely, sausages contain lower amounts of sugars and vitamins, but higher levels of free amino acids throughout the ripening process [48,49]. Therefore, these differences could be the key factor for the efficient inhibition of A. parasiticus by D. hansenii, and P. acidilactici combined with PgAFP on sliced sausage.
On the other hand, the same strain of D. hansenii used here was barely active at low (0.84) compared to high (0.94) aw values when grown on dry-cured ham slices for 15 days [30]. Therefore, D. hansenii alone might not efficiently inhibit A. parasiticus on sausages at aw values around 0.85. Conversely, the combination of PgAFP and D. hansenii (Pg + Dh) efficiently inhibited A. parasiticus growth on sliced sausage (Table 2). Thus, the combined PgAFP could be decisive to help the other biological agents reach an efficient inhibition of mycotoxigenic molds in dry-cured meats, particularly at low aw values.
The impact of PgAFP on aflatoxins production greatly differs with the culture medium used (Figure 2). The increased aflatoxins levels in YES broth is attributed to ROS induction by antifungal proteins, which is a prerequisite for aflatoxin production in A. parasiticus [21]. Conversely, the decreased aflatoxins levels in Ca-YES broth must be related to the calcium-triggered resistance against PgAFP mediated by calcineurin, G protein, and γ-glutamyltraspeptidase, that combat oxidative stress in A. flavus [20]. In fact, aflatoxin production was also inhibited in Ca-YES by most of the tested treatments, except for AFG1 in the batch inoculated only with D. hansenii (Figure 2).
Cocultivation of A. parasiticus with P. acidilactici in YES led to lower AFB1 production, irrespective of the addition of PgAFP (Figure 2). Therefore, P. acidilactici seems to prevent the increment of mycotoxin production provoked by PgAFP. The mechanism behind the lower aflatoxin production with lactic acid bacteria is not known, but it has been related to low molecular weight inhibitory compounds as well as removing due to aflatoxin binding [50].
The oxidative status in the fungal cell severely affects mycotoxin production [51]. Several antifungal proteins from molds provoke ROS generation in sensitive strains [18,19], including PgAFP in A. flavus [16]. ROS production is regarded as a prerequisite for the onset of aflatoxin production [22,23]. Conversely, several antioxidant substances and enzymes inhibit aflatoxin production [22,52]. For all this, the effect of PgAFP, used alone or combined with the two microbial agents, on the expression of foxA and aflP genes was studied in A. parasiticus.
ROS induction has been proposed to play a key role in the mechanism of action of PgAFP on sensitive molds [16]. The effect of ROS on aflatoxin synthesis has been related to redox balance through β-oxidation, as studied through foxA gene expression [23]. Besides, the aflP gene has been used to study the effect of environmental factors on aflatoxin production [26]. Thus, aflP and foxA expression was assessed to study the effect of PgAFP and the other two biocontrol agents on aflatoxin production.
aflP expression in A. parasiticus grown in YES broth was not overexpressed, but repressed by PgAFP, as well as by P. acidilactici and the combined Pg + Dh + Pa treatment (Figure 3). In Ca-YES broth, aflP expression was also repressed by PgAFP, but overexpressed in samples treated only with P. acidilactici. Despite the differences in aflP expression, no statistically significant difference was obtained in the aflatoxin levels from these batches. The lack of correlation between gene expression and aflatoxin production may be related to the fact that aflP is a structural rather than a regulatory gene [53]. According to these results, the potential effect of PgAFP promoting aflatoxin production has to be ruled out in calcium-enriched media.
On the other hand, the foxA gene expression showed no statistically significant difference due to PgAFP treatment in both YES and Ca-YES broth. These results are consistent with the lower relative abundance of O-methyltransferase, coded by the aflP gene, and the unaffected level of peroxisomal multifunctional β-oxidation protein, coded by the foxA gene, found in PgAFP-treated A. flavus [16], as well as with the low levels of intracellular ROS detected in PgAFP-treated A. flavus grown in Ca-enriched PDB [20]. In addition, D. hansenii did not alter foxA expression, supporting that the effect of this yeast reducing aflatoxin levels is not due to a potential antioxidant effect. Therefore, the overproduction of aflatoxin observed in PgAFP-treated A. parasiticus grown in Ca-YES broth (Figure 2) may not be initiated by an increased β-oxidation due to ROS. In addition, the increased foxA expression obtained in Pg + Dh + Pa treated samples from YES broth was not accompanied by higher aflatoxin production at the 15th day. These results suggest that the effect of ROS on aflatoxin biosynthesis cannot be explained solely through β-oxidation. The generation of secondary ROS in endosomes upon exposure to exogenous ROS during aflatoxins biosynthesis [54] suggests that other pathways may be involved in the metabolism of these toxic compounds, such as calcineurin signaling related to the role of calcium as a secondary messenger in the response to antifungal proteins in fungi [55]. However, these studies are still in a preliminary stage and require further investigation.
A. parasiticus produced AFB1 and AFG1 when grown on the untreated sausages, reaching levels well above the levels regarded as safe for various foods [56] both at five and 15 days of incubation (Table 1). The amount of AFB1 and AFG1 produced on sausages was higher than that obtained in YES broth, which can be explained by the limited availability of nutrients [57] and the increased levels of free amino acids [58] in the sausage. Aflatoxin production was greatly reduced by Pg + Dh and Pg + Dh + Pa treatments on sausage samples (Table 2). AFB1 and AFG1 were not detected in treated samples at Day 5 of incubation and only slightly above the limits set for foods [56] at 15 days. However, given that the conditions set were favorable for A. parasiticus development and mycotoxin production, it is expected that any of the two combinations tested may be sufficient to control the usual fungal contamination during dry sausage ripening.
The successful antifungal effect of both treatments with Pg + Dh on dry-fermented sausage supports their use as a preventive measure against aflatoxigenic A. parasiticus or as a corrective action when this mold has been detected on the meat product. The inclusion of P. acidilactici as an additional protective culture did not substantially improve the inhibition on A. parasiticus growth or mycotoxin production. Thus, the contribution of P. acidilactici seems to be negligible. This fact has to be related to the poor ability of this bacterium to grow on dry-fermented sausages.
Given that both the mycelial weight and mycotoxin production were inhibited by some treatments in Ca-YES broth, the efficacy of the most effective treatments containing PgAFP was tested on cheese slices. The low aw levels reached during cheese ripening, even lower than 0.90 in the rind of some cheeses [59], could restrict survival of P. acidilactici and D. hansenii. The aw values in treated cheese slices decreased to 0.89 and 0.86 at Days 5 and 15, respectively (Table 2), which are below the 0.90 minimum aw level for Pediococcus growth in foods [60] , but not below the 0.83 minimum value for D. hansenii [61] and aflatoxins formation by A. parasiticus [62].
On cheese slices, both Pg + Dh and Pg + Dh + Pa treatments effectively inhibited A. parasiticus growth and aflatoxins production for 15 days. This effect was stronger than that reported by the sole action of D. hansenii on ochratoxigenic P. nordicum in dry-cured ham incubated at 0.84 aw [30]. No additional inhibition was observed by the combined use of P. acidilactici with PgAFP and D. hansenii (Table 2), which could be due to P. acidilactici failing to thrive under the restrictive aw values. However, the samples inoculated with P. acidilactici showed the highest LAB load. Although production of antifungal proteinaceous compounds by LAB in fresh cheese has been reported [63], the lower aw values in ripened cheese might limit the production of antifungal compounds by P. acidilactici.
In conclusion, the combined treatment of PgAFP and D. hansenii inhibits A. parasiticus growth and mycotoxin production in dry-fermented sausages and cheese at similar conditions to those during the ripening process. P. acidilactici down-regulates aflP gene in A. parasiticus grown in YES broth, lowering aflatoxin production, but it does not provide further protection in sausage or cheese treated with PgAFP and D. hansenii. The increased aflatoxin production induced by PgAFP does not involve an increased foxA expression, but knowing the exact role of ROS will require further studies. On the whole, these results confirm the usefulness of PgAFP to control the aflatoxigenic population on dry-fermented foods, even on calcium-rich ones.
Author Contributions
Conceptualization, J.D., A.R. and F.N.; Methodology, J.D., A.R. and F.N.; Software, J.D. and A.R.; Validation, A.R., M.A. and F.N.; Formal Analysis, J.D. and A.R.; Investigation, J.D. and A.G.; Resources, F.N.; Data Curation, A.R., M.A. and F.N.; Writing-Original Draft Preparation, J.D. and A.R.; Writing-Review & Editing, M.A. and F.N.; Visualization, J.D., A.R., M.A. and F.N.; Supervision, M.A. and F.N.; Project Administration, F.N.; Funding Acquisition, F.N.
Funding
This work was supported by the Spanish Ministry of Education and Science, Ministry of Economy and Competitiveness (AGL2013-45729-P, AGL2016-80209-P), and Junta de Extremadura and FEDER (GR15108).
Acknowledgments
Josué Delgado was recipient of a FPI grant from the Spanish Ministry of Education and Science (BES-2011-043422). Alicia Rodríguez is supported by a Juan de la Cierva research contract (IJCI-2014-20666) from the Spanish Ministry of Economy and Competitiveness. Alfredo García acknowledges his contract (TA13003) from Junta de Extremadura and European Social Fund.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alapont, C.; López-Mendoza, M.C.; Gil, J.V.; Martínez-Culebras, P.V. Mycobiota and toxigenic Penicillium species on two Spanish dry-cured ham manufacturing plants. Food Addit. Contam. A 2014, 31, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.L.; Domínguez, L.; Gómez-Lucía, E.; Garayzabal, J.F.F.; Goyache, J.; Suárez, G. Experimental aflatoxin production in Manchego-type cheese. J. Appl. Bacteriol. 1988, 64, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Hymery, N.; Vasseur, V.; Coton, M.; Mounier, J.; Jany, J.L.; Barbier, G.; Coton, M. Filamentous fungi and mycotoxins in cheese: A review. Compr. Rev. Food Sci. F 2014, 13, 437–456. [Google Scholar] [CrossRef]
- Lie, J.L.; Marth, E.H. Formation of aflatoxin in cheddar cheese by Aspergillus flavus and Aspergillus parasiticus. J. Dairy Sci. 1967, 50, 1708–1710. [Google Scholar] [CrossRef]
- Taniwaki, M.H.; Hocking, A.D.; Pitt, J.I.; Fleet, G.H. Growth of fungi and mycotoxin production on cheese under modified atmospheres. Int. J. Food Microbiol. 2001, 68, 125–133. [Google Scholar] [CrossRef]
- Markov, K.; Pleadin, J.; Bevardi, M.; Vahčić, N.; Sokolić-Mihalak, D.; Frece, J. Natural occurrence of aflatoxin B1, ochratoxin A and citrinin in Croatian fermented meat products. Food Control 2013, 34, 312–317. [Google Scholar] [CrossRef]
- Pleadin, J.; Malenica, M.; Vah, N.; Milone, S.; Safti, L. Survey of aflatoxin B1 and ochratoxin A occurrence in traditional meat products coming from Croatian households and markets. Food Control 2015, 52, 71–77. [Google Scholar] [CrossRef]
- Rodríguez, A.; Rodríguez, M.; Martín, A.; Delgado, J.; Córdoba, J.J. Presence of ochratoxin A on the surface of dry-cured Iberian ham after initial fungal growth in the drying stage. Meat Sci. 2012, 92, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Rodríguez, M.; Martín, A.; Núñez, F.; Córdoba, J.J. Evaluation of hazard of aflatoxin B1, ochratoxin A and patulin production in dry-cured ham and early detection of producing moulds by qPCR. Food Control 2012, 27, 118–126. [Google Scholar] [CrossRef]
- Delgado, J.; Acosta, R.; Rodríguez-Martín, A.; Bermúdez, E.; Núñez, F.; Asensio, M.A. Growth inhibition and stability of PgAFP from Penicillium chrysogenum against fungi common on dry-ripened meat products. Int. J. Food Microbiol. 2015, 205, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Marx, F. Small, basic antifungal proteins secreted from filamentous ascomycetes: a comparative study regarding expression, structure, function and potential application. Appl. Microbiol. Biotechnol. 2004, 65, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Tóth, L.; Kele, Z.; Borics, A.; Nagy, L.G.; Váradi, G.; Virágh, M.; Takó, M.; Vágvölgyi, C.; Gallgóczy, L. NFAP2, a novel cysteine-rich anti-yeast protein from Neosartorya fischeri NRRL 181: isolation and characterization. AMB Express 2016, 6, 75. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.; Hajdu, D.; Bratschun-Khan, D.; Gáspári, Z.; Varbanov, M.; Philippot, S.; Fizil, A.; Czajlik, A.; Kele, Z.; Sonderegger, C.; et al. New antimicrobial potential and structural properties of PAFB: A cationic, cysteine-rich protein from Penicillium chrysogenum Q176. Sci. Rep. 2018, 8, 1751. [Google Scholar] [CrossRef] [PubMed]
- Tóth, L.; Váradi, G.; Borics, A.; Batta, G.; Kele, Z.; Vendrinszky, A.; Tóth, R.; Ficze, H.; Tóth, G.; Vagvölgy, C.; Marx, F.; Galgóczy, L. Anti-candidal activity and functional mapping of recombinant and synthetic Neosartorya fischeri antifungal protein 2 (NFAP2). Front. Microbiol. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martín, A.; Acosta, R.; Liddell, S.; Núñez, F.; Benito, M.J.; Asensio, M.A. Characterization of the novel antifungal protein PgAFP and the encoding gene of Penicillium chrysogenum. Peptides 2010, 31, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Delgado, J.; Owens, R.A.; Doyle, S.; Asensio, M.A.; Núñez, F. Impact of the antifungal protein PgAFP from Penicillium chrysogenum on the protein profile in Aspergillus flavus. Appl. Microbiol. Biotechnol. 2015, 99, 8701–8715. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ao, J.; Yang, W.; Jiao, L.; Zheng, T.; Chen, X. Purification and characterization of a novel antifungal protein secreted by Penicillium chrysogenum from an Arctic sediment. Appl. Microbiol. Biotechnol. 2013, 97, 10381–10390. [Google Scholar] [CrossRef] [PubMed]
- Galgóczy, L.; Kovács, L.; Karácsony, Z.; Virágh, M.; Hamari, Z.; Vágvölgyi, C. Investigation of the antimicrobial effect of Neosartorya fischeri antifungal protein (NFAP) after heterologous expression in Aspergillus nidulans. Microbiology 2013, 159, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Kaiserer, L.; Oberparleiter, C.; Weiler-Görz, R.; Burgstaller, W.; Leiter, E.; Marx, F. Characterization of the Penicillium chrysogenum antifungal protein PAF. Arch. Microbiol. 2003, 180, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Delgado, J.; Owens, R.A.; Doyle, S.; Núñez, F.; Asensio, M.A. Quantitative proteomics reveals new insights into calcium-mediated resistance mechanisms in Aspergillus flavus against the antifungal protein PgAFP in cheese. Food Microbiol. 2017, 66, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jayashree, T.; Subramanyan, C. Oxidative stress as a prerequisite for aflatoxin production by Aspergillus parasiticus. Free Radical Biol. Med. 2000, 29, 981–985. [Google Scholar] [CrossRef]
- Reverberi, M.; Fabbri, A.A.; Zjalic, S.; Ricelli, A.; Punelli, F.; Fanelli, C. Antioxidant enzymes stimulation in Aspergillus parasiticus by Lentinula edodes inhibits aflatoxin production. Appl. Microbiol. Biotechnol. 2005, 69, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Punelli, M.; Smith, C.A.; Zjalic, S.; Scarpari, M.; Scala, V.; Fanelli, C. How peroxisomes affect aflatoxin biosynthesis in Aspergillus flavus. PLoS ONE 2012, 7, e48097. [Google Scholar] [CrossRef] [PubMed]
- Maggio-Hall, L.A.; Wilson, R.A.; Keller, N.P. Fundamental contribution of beta-oxidation to polyketide mycotoxin production in planta. Mol. Plant Microbe Int. 2005, 18, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Chanda, A.; Laivenieks, M.; Beaudry, R.M.; Artymovich, K.A.; Koptina, A.V.; Linz, J.E. Volatile profiling reveals intracellular metabolic changes in Aspergillus parasiticus: veA regulates branched chain amino acid and ethanol metabolism. BMC Biochem. 2010, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Ojalvo, D.; Rodríguez, A.; Bernáldez, V.; Córdoba, J.J.; Rodríguez, M. Influence of temperature and substrate conditions on the omt-1 gene expression of Aspergillus parasiticus in relation to its aflatoxin production. Int. J. Food Microbiol. 2013, 166, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, R.C.S.; Gouvêa, D.M.; Hungaro, H.M.; Sodré, A. de F.; Querol-Simon, A. Dynamics of the yeast flora in artisanal country style and industrial dry cured sausage (yeast in fermented sausage). Food Control 2013, 29, 143–148. [Google Scholar] [CrossRef]
- Núñez, F.; Rodríguez, M.M.; Córdoba, J.J.; Bermúdez, M.E.; Asensio, M.A. Yeast population during ripening of dry-cured Iberian ham. Int. J. Food Microbiol. 1996, 29, 271–280. [Google Scholar] [CrossRef]
- Andrade, M.J.; Thorsen, L.; Rodríguez, A.; Córdoba, J.J.; Jespersen, L. Inhibition of ochratoxigenic moulds by Debaryomyces hansenii strains for biopreservation of dry-cured meat products. Int. J. Food Microbiol. 2014, 170, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Núñez, F.; Lara, M.S.; Peromingo, B.; Delgado, J.; Sanchez-Montero, L.; Andrade, M.J. Selection and evaluation of Debaryomyces hansenii isolates as potential bioprotective agents against toxigenic penicillia in dry-fermented sausages. Food Microbiol. 2015, 46, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Peromingo, B.; Núñez, F.; Rodríguez, A.; Alía, A.; Andrade, M.J. Potential of yeasts isolated from dry-cured ham to control ochratoxin A production in meat models. Int. J. Food Microbiol. 2018, 268, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, S.; Lu, J.; Liu, L.; Uluko, H.; Pang, X.; Lv, J. Antifungal activities and effect of Lactobacillus casei AST18 on the mycelia morphology and ultrastructure of Penicillium chrysogenum. Food Control 2014, 43, 57–64. [Google Scholar] [CrossRef]
- Effat, B.A.; Ibrahim, G.A.; Tawfik, N.F.; Sharaf, O.M. Comparison of antifungal activity of metabolites from Lactobacillus rhamnosus, Pediococcus acidilactici and Propionibacterium thoenii. Egypt. J. Dairy Sci. 2001, 29, 251–262. [Google Scholar]
- Montiel, R.; Bravo, D.; Medina, M. Commercial biopreservatives combined with salt and sugar to control Listeria monocytogenes during smoked salmon processing. J. Food Prot. 2013, 76, 1463–1465. [Google Scholar] [CrossRef] [PubMed]
- Mandal, V.; Sen, S.K.; Mandal, N.C. Production and partial characterisation of an inducer-dependent novel antifungal compound(s) by Pediococcus acidilactici LAB 5. J. Sci. Food Agr. 2013, 93, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Acosta, R.; Rodríguez-Martín, A.; Martín, A.; Núñez, F.; Asensio, M.A. Selection of antifungal protein-producing molds from dry-cured meat products. Int. J. Food Microbiol. 2009, 135, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Long, G.L.; Winefordner, J.D. Limit of detection, a closer look at the IUPAC definition. Anal. Chem. 1983, 55, 712–724. [Google Scholar]
- Currie, L.A. Nomenclature in evaluation of analytical methods including detection and quantification capabilities. Anal. Chim. Acta 1999, 391, 105–126. [Google Scholar] [CrossRef]
- Rodríguez, A.; Medina, Á.; Córdoba, J.J.; Magan, N. The influence of salt (NaCl) on ochratoxin A biosynthetic genes, growth and ochratoxin A production by three strains of Penicillium nordicum on a dry-cured ham-based medium. Int. J. Food Microbiol. 2014, 178, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Rodríguez, M.; Luque, M.I.; Martín, A.; Córdoba, J.J. Real-time PCR assays for detection and quantification of aflatoxin-producing molds in foods. Food Microbiol. 2012, 31, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Prot. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Cizeikiene, D.; Juodeikiene, G.; Paskevicius, A.; Bartkiene, E. Antimicrobial activity of lactic acid bacteria against pathogenic and spoilage microorganism isolated from food and their control in wheat bread. Food Control 2013, 31, 539–545. [Google Scholar] [CrossRef]
- Liu, P.; Luo, L.; Long, C. Characterization of competition for nutrients in the biocontrol of Penicillium italicum by Kloeckera apiculata. Biol. Control 2013, 67, 157–162. [Google Scholar] [CrossRef]
- Andrade, M.J.; Córdoba, J.J.; Casado, E.M.; Córdoba, M.G.; Rodríguez, M. Effect of selected strains of Debaryomyces hansenii on the volatile compound production of dry fermented sausage “salchichón”. Meat Sci. 2010, 85, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Demasi, T.W.; Wardlaw, F.B.; Dick, R.L.; Acton, J.C. Nonprotein nitrogen (NPN) and free amino acid contents of dry, fermented and nonfermented sausages. Meat Sci. 1990, 27, 1–12. [Google Scholar] [CrossRef]
- Hierro, E.; de la Hoz, L.; Ordóñez, J.A. Contribution of microbial and meat endogenous enzymes to the lipolysis of dry fermented sausages. J. Agric. Food Chem. 1999, 45, 2989–2995. [Google Scholar] [CrossRef]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria. Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Stoll, D.; Schütz, P.; Geisen, R. Oxidative stress induces the biosynthesis of citrinin by Penicillium verrucosum at the expense of ochratoxin. Int. J. Food Microbiol. 2014, 192, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yu, J.; Mahoney, N.; Chan, K.L.; Molyneux, R.J.; Varga, J.; Bhatnagar, D.; Thomas, E.; Cleveland, T.E.; Nierman, W.C.; Campbell, B.C. Elucidation of the functional genomics of antioxidant-based inhibition of aflatoxin biosynthesis. Int. J. Food Microbiol. 2008, 122, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Scherm, B.; Palomba, M.; Serra, D.; Marcello, A.; Migheli, Q. Detection of transcripts of the aflatoxin genes aflD, aflO, and aflP by reverse transcription-polymerase chain reaction allows differentiation of aflatoxin-producing and non-producing isolates of Aspergillus flavus and Aspergillus parasiticus. Int. J. Food Microbiol. 2005, 98, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.; Laivenieks, M.; Hong, S.-Y.; Wee, J.; Wong, S.-S.; Vanos, B.; Linz, J. Aflatoxin biosynthesis is a novel source of reactive oxygen species. A potential redox signal to initiate resistance to oxidative stress? Toxins 2015, 7, 1411–1430. [Google Scholar] [CrossRef] [PubMed]
- Binder, U.; Bencina, M.; Eigentler, A.; Meyer, V.; Marx, F. The Aspergillus giganteus antifungal protein AFPNN5353 activates the cell wall integrity pathway and perturbs calcium homeostasis. BMC Microbiol. 2011, 11, 209. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission regulation (CE) 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Offic. J. Eur. Union 2006, L364, 5–24. [Google Scholar]
- Miller, J.D. Factors that affect the occurrence of fumonisin. Environ. Health Persp. 2001, 109 (Suppl. 2), 321–324. [Google Scholar] [CrossRef]
- Payne, G.A.; Hagler, W.M., Jr. Effect of specific amino acids on growth and aflatoxin production by Aspergillus parasiticus and Aspergillus flavus in defined media. Appl. Environ. Microbiol. 1983, 46, 805–812. [Google Scholar] [PubMed]
- Rüegg, M.; Blanc, B. Influence of water activity on the manufacture and aging of cheese. In Water Activity. Influences on Food Quality; Rockland, L.B., Stewart, G.F., Eds.; Academic Press, Inc.: New York, NY, USA, 1981; pp. 791–893. [Google Scholar]
- Leistner, L.; Rödel, W.; Krispien, K. Microbiology of meat and meat products in high- and intermediate-moisture ranges. In Water Activity. Influences on Food Quality; Rockland, L.B., Stewart, G.F., Eds.; Academic Press, Inc.: New York, NY, USA, 1981; pp. 791–893. [Google Scholar]
- Troller, J.A.; Christian, J.H.B. Water Activity and Foods; Academic Press, Inc.: New York, NY, USA, 1978. [Google Scholar]
- Peromingo, B.; Rodríguez, A.; Bernáldez, V.; Delgado, J.; Rodríguez, M. Effect of temperature and water activity on growth and aflatoxin production by Aspergillus flavus and Aspergillus parasiticus on cured meat model systems. Meat Sci. 2016, 122, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Sedaghat, H.; Eskandari, M.H.; Moosavi-Nasab, M.; Shekarforoush, S.S. Application of non-starter lactic acid bacteria as biopreservative agents to control fungal spoilage of fresh cheese. Int. Dairy J. 2016, 56, 87–91. [Google Scholar] [CrossRef]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).